
干旱和施氮对苹果大苗的水分状况、生长和碳素分配的影响
2024-01-30 15:19薛浩刘毅吕英忠张拥兵王林
经济林研究 2023年3期
薛浩 刘毅 吕英忠 张拥兵 王林


摘 要:【目的】为苹果大苗的水分和养分管理提供理论参考。【方法】选用‘丹霞苹果大苗为材料,分别设置正常浇水(土壤含水量为25% ~ 30%)与干旱(土壤含水量为10% ~ 15%)2 个水分处理,每种水分条件下分别设置每株0、20、60 g 氮肥施用量3 个处理(分别以低氮、中氮、高氮表示)。在处理3 个月后,测定苹果大苗的水分状况、光合作用指标、各部位生物量和非结构性碳水化合物(NSC)含量。【结果】在正常供水条件下:随着施氮量的增加,枝条水势降低,导水损失率增加,水分条件变差;中氮处理对净光合速率的促进作用最大,茎生物量随着施氮量增多显著增加,高氮处理下枝条和叶片的NSC 含量均低于低氮处理。在干旱条件下,随着施氮量的增加,枝条水势降低,导水损失率升高,净光合速率和气孔导度降低,各部位生物量减少,枝条韧皮部及木质部的NSC 含量降低。【结论】正常供水条件下,过量施氮导致苹果大苗水分状况变差,这会抵消氮对光合速率的促进作用,进而影响树木的生长和生物量积累。在干旱条件下,施氮进一步引起水分条件恶化,限制光合作用和生物量积累,同时过量施氮会导致NSC 含量降低,可能进一步影响植株对干旱的适应能力,因此在干旱条件下应减少氮肥的施用量。
关键词:苹果大苗;施氮;干旱;水分状况;光合速率;生长;非结构性碳水化合物
中图分类号:S661.1 文献标志码:A 文章编号:1003—8981(2023)03—0115—09
水分和氮素是树木生长和存活的必要物质[1-2]。二者都会影响到树木的光合作用、生长和生物量分配[3-5]。因此,了解在不同水氮条件下树木的水碳代谢和生长情况,有利于进行针对性的水肥管理。大量研究结果表明,干旱导致树木水势降低,在较大的负压下木质部导管内产生气穴栓塞,影响水分长距离运输的速率[6-7],并进一步限制光合作用[8-9],影响生长和碳水化合物分配。通常认为氮的可用性增加可以促进光合作用的进行[5,10],提高树木的生长速率[11];施氮也会改变树木的碳素分配模式[12],一般认为施氮会较大程度上促進地上部分生长,但目前仍不完全了解水分匮乏和氮素水平的交互作用对树木生长、碳素分配的影响,尤其是对树木水力结构的影响,因此研究树木在不同水氮条件下的水力结构、光合作用、生物量和非结构性碳水化合物分配,有利于全面了解树木在水氮互作下的水分和碳水化合物代谢过程及对生长的影响。
黄土高原产区是我国苹果的主产区之一,水分和氮素管理是该产区基本的栽培管理手段[13]。本研究中以苹果Malus pumila 大苗为研究对象,通过比较不同干旱和氮肥的施入条件下苹果大苗水分状况、光合作用、生物量分配、碳水化合物分配的变化,研究水分条件和施氮对苹果水碳代谢和生物量分配的影响,旨在了解水氮互作对苹果树水碳代谢的响应模式,为苹果水分和氮素综合管理提供理论参考。
1 材料与方法
1.1 试验地概况
在山西农业大学果树研究所(112°50′03″E,37°34′40″N)进行试验。试验区海拔845 m,属温带大陆性季风气候,年均温5 ~ 10 ℃,年均太阳总辐射量5 100 MJ/m2,无霜期160 ~ 190 d。
1.2 试验材料
苹果大苗为2 年生‘丹霞苹果嫁接苗木,砧木为八棱海棠Malus × robusta,苗木于1 年前定植在直径35 cm、深度40 cm 的塑料控根容器中,苗高为(101.5±12.3) cm,地径(嫁接口向上1 cm)为(16.03±1.82) mm。供试基质为本地园土、草炭土、牛粪、珍珠岩的混合基质,质量比为2∶3∶1∶1,有机质含量为44.5 g/kg,全氮含量为0.54 g/kg,有效磷含量为45.1 mg/kg,速效钾含量为485 mg/kg,pH 值为7.85。
1.3 试验设计
2022 年4 月初,选取长势基本一致且无病虫害的36 株苹果大苗,放入透明避雨大棚中,保持间距。设置2 种土壤水分条件[9]:正常浇水(土壤含水量为25% ~ 30%)与干旱(土壤含水量为10% ~ 15%)。设置3 种供氮水平:低氮(每株共施尿素0 g)、中氮(每株共施尿素20 g)和高氮(每株共施尿素60 g)[14]。设置低氮正常浇水、中氮正常浇水、高氮正常浇水、低氮干旱、中氮干旱、高氮干旱,共6 个处理。将预计施入的尿素量平均分3 份,分别于2022 年4 月初、5 月初、6 月初施入,在第1 次施肥开始的同时进行控水,7 月开始测定枝条水势和导水损失率、光合作用指标、不同部位的生物量和非结构性碳水化合物含量。每个处理的3 株大苗用于测定各生理指标,另外3 株用于测定生物量及生物量分配指标。
1.4 指标测定
1.4.1 枝条水势和导水损失率
在2022 年7 月中旬选择连续晴天,测定枝条的水势。在04:00—05:30( 日出前), 用Model 600 型便携式植物水势仪(PMS,美国)测定枝条水势。枝条导水损失率的测定时间同枝条水势,水下剪取长约20 cm 的树冠中上部向阳面枝条,使剪口保持浸没在水中,迅速带回试验室,参照王林等[15] 的方法测定导水损失率。
1.4.2 叶片光合作用指标
2022 年7 月中旬,选择连续晴天的09:00—11:00,用LI-6400 便携式光合作用测量系统(LICOR,美国)的红蓝光源叶室测定枝条气体交换指标。设定光照强度为1 500 μmol/(m2·s),预试验结果表明此光照强度在该品种的光饱和点之上。选择向阳枝条中部成熟叶片进行测定,记录叶片光合速率、气孔导度、蒸腾速率,并根据光合速率与蒸腾速率的比值计算水分利用效率。
1.4.3 生物量及NSC
在2022 年7 月,每组处理采收3 株,将采集的所有苹果大苗分成叶、茎、粗根(直径大于2 mm)和细根(直径不大于2 mm)4 部分。所有样品置于105 ℃条件下杀青20 min,后于80 ℃条件下烘干至恒定质量,然后称量并记录不同部位的生物量。烘干后粉碎并过土壤筛(筛孔直径0.18 mm),采用硫酸蒽酮法[16],测定茎木质部和韧皮部以及叶片的淀粉、可溶性糖含量,非结构性碳水化合物(non-structural carbohydrate,NSC)含量为可溶性糖和淀粉的含量之和。
1.5 数据处理
每个指标的测定均重复3 次以上。所有数据均使用SPSS 26.0 软件进行统计分析,使用OnewayANOVA 方法和邓肯(Duncan)多重比较法检验测定指标的差异显著性,显著性水平设定为α=0.05,使用双因素(土壤水分条件和施氮量)方差分析法比较土壤水分条件、施氮量及其交互作用对树木生理特征及各部位生物量的影响。使用Sigmaplot 10.0 软件绘图。
2 结果与分析
2.1 干旱和施氮对苹果大苗枝条水势和导水损失率的影响
由图1 可见,在正常供水条件下:随着施氮量的增加,枝条水势呈降低趋势,低氮处理的枝条水势显著高于高氮处理(P < 0.05);枝条导水损失率随施氮量的增加呈增加趋势,低氮处理的枝条导水损失率显著低于高氮处理(P < 0.05),中氮处理的枝条导水损失率略高于低氮处理,但两者间无显著差异。在干旱条件下:随着施氮量的增加,枝条水势呈降低趋势,低氮处理的水势显著高于中氮和高氮处理(P < 0.05);枝条导水损失率随施氮量的增加呈增加趋势,低氮处理的枝条导水损失率显著低于高氮处理(P < 0.05),中氮处理的枝条导水损失率略高于低氮处理,但两者间无显著差异。
干旱和施氮的交互作用对苹果大苗的枝条水势有显著影响(P < 0.05),对导水损失率无显著影响(表1)。
2.2 干旱和施氮对苹果大苗叶片光合作用的影响
由图2 可见,在正常供水条件下:叶片净光合速率随施氮量的增加呈现先增加后降低的趨势,中氮处理的净光合速率显著高于低氮和高氮处理(P < 0.05),高氮处理的净光合速率显著高于低氮处理(P < 0.05),中氮和高氮处理的净光合速率分别比低氮处理增加了21.0%、10.0%;叶片的气孔导度和胞间CO2 浓度在不同氮素水平下无显著差异;叶片水分利用效率随施氮量的增加呈增加趋势,中氮和高氮处理的叶片水分利用效率显著高于低氮处理(P < 0.05)。
由图2 可见,在干旱条件下:叶片的净光合速率随施氮量增加呈现降低趋势,不同处理间均存在显著差异(P < 0.05),中氮和高氮处理的净光合速率分别比低氮处理降低了26.6%、50.3%;叶片的气孔导度也随施氮量增加呈现降低趋势,低氮处理下的叶片气孔导度显著高于中氮和高氮处理(P < 0.05);叶片胞间CO2 浓度随施氮量增加呈现降低趋势,低氮处理下的胞间CO2 浓度显著高于高氮处理(P < 0.05);叶片的水分利用效率随施氮量增加呈现增加趋势,高氮处理下叶片的水分利用效率显著高于低氮处理(P < 0.05)。
干旱和施氮的交互作用对苹果大苗的净光合速率有极显著影响(P < 0.01),对胞间CO2 浓度有显著影响(P < 0.05),对气孔导度和水分利用效率无显著影响(表2)。
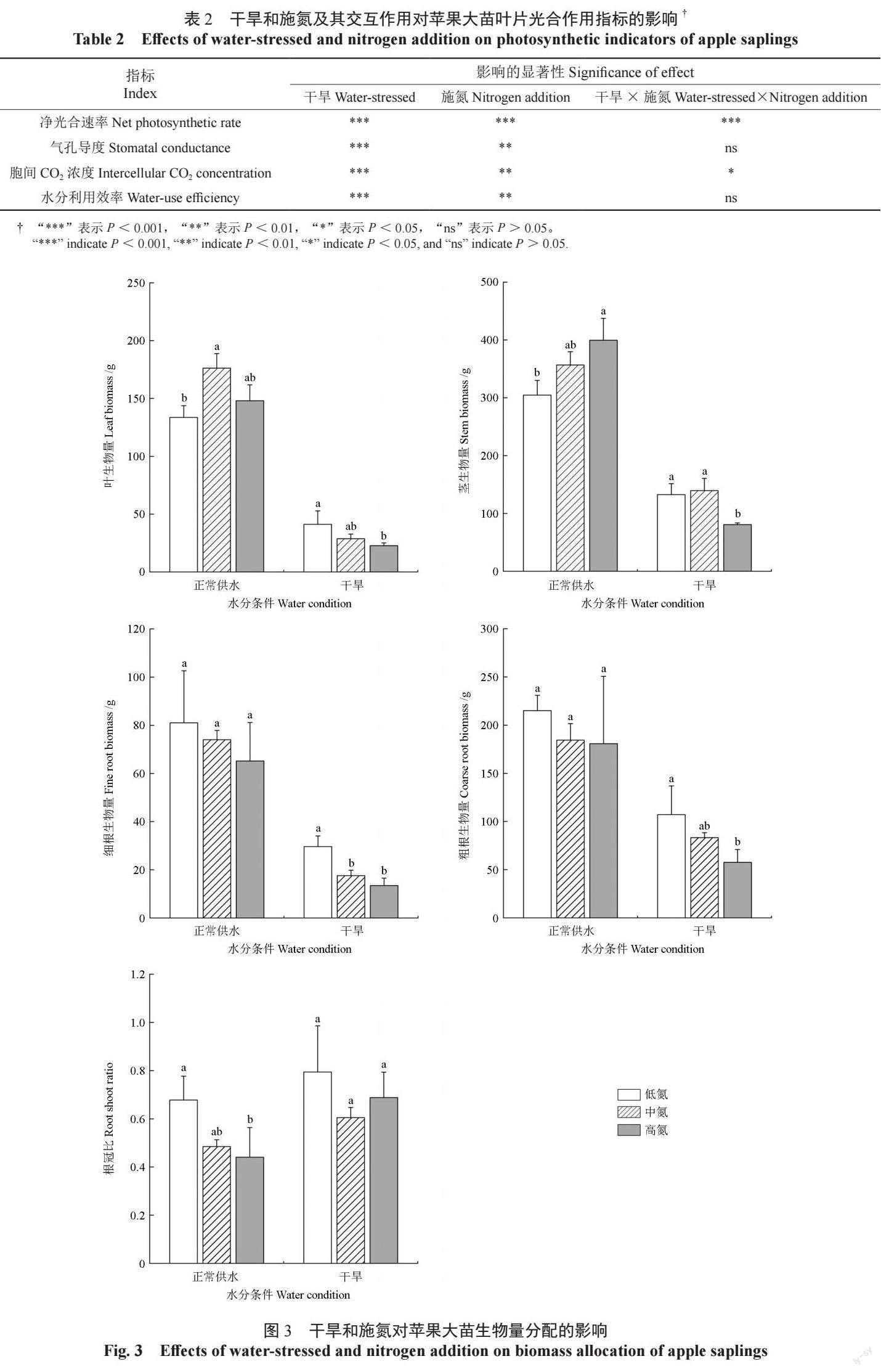
2.3 干旱和施氮对苹果大苗生物量分配的影响
由图3 可见,在正常供水条件下:中氮处理的叶生物量显著高于低氮处理(P < 0.05),高氮处理的叶生物量略高于低氮处理,但两者间无显著差异;高氮处理的茎生物量显著高于低氮处理(P < 0.05),中氮处理的茎生物量略高于低氮处理,但两者间无显著差异;粗根生物量和细根生物量在不同处理间无显著差异,中氮和高氮处理的粗根生物量和细根生物量略低于低氮处理;根冠比随施氮量增加呈现降低趋势,高氮处理的根冠比显著低于低氮处理(P < 0.05),中氮处理的根冠比略高于高氮处理,但两者间无显著差异。
由图3 可见,在干旱条件下:叶生物量、粗根生物量、细根生物量均随着施氮量的增加呈现减小趋势,高氮处理下的叶生物量、粗根生物量均显著低于低氮处理(P < 0.05),中氮处理的叶生物量、粗根生物量均略低于低氮处理,但两者间无显著差异;低氮和中氮处理的茎生物量显著高于高氮处理(P < 0.05);中氮处理和高氮处理的细根生物量显著低于低氮处理(P < 0.05)。干旱条件下不同部位的生物量均显著低于正常供水条件下的生物量(P < 0.05)。
干旱和施氮的交互作用对苹果大苗叶和茎的生物量有显著影响(P < 0.05),对其他部位的生物量无显著影响(表3)。
2.4 干旱和施氮对苹果大苗茎叶NSC 含量的影响
由图4 可见,在正常供水条件下:不同施氮量下木质部和韧皮部可溶性糖含量无显著差异;木质部和韧皮部淀粉含量均随着施氮量的增加呈现减少趋势,且在不同施氮量处理间均有显著差异(P < 0.05); 木质部和韧皮部NSC 含量均随着施氮量的增加呈现减少趋势,低氮处理和中氮处理下的木质部NSC 含量显著高于高氮处理(P < 0.05),低氮处理和中氮处理之间的木质部NSC 含量无显著差异;低氮处理下的韧皮部NSC含量显著高于中氮处理和高氮处理(P < 0.05),中氮处理和高氮处理之间的韧皮部NSC 含量无显著差异;低氮处理的叶片可溶性糖含量显著高于中氮处理(P < 0.05),高氮处理的叶片可溶性糖含量略高于中氮处理,但两者间无显著差异;低氮处理下的叶片淀粉和NSC 含量均显著高于中氮和高氮处理(P < 0.05),中氮和高氮处理下叶片淀粉和NSC 含量均无显著差异。
由图4 可见,在干旱条件下:木质部可溶性糖含量随着施氮量的增加呈现减小趋势,低氮处理下的木质部可溶性糖含量显著高于高氮处理(P < 0.05);低氮处理下的韧皮部可溶性糖含量显著高于中氮处理和高氮处理(P < 0.05),中氮处理和高氮处理之间无显著差异;木质部和韧皮部淀粉含量在不同施氮量下无显著差异;木质部和韧皮部NSC 含量在不同施氮量下无显著差异。
随着施氮量的增加,叶片可溶性糖含量呈现先降低后增加的趋势,且各处理之间均呈现显著差异(P < 0.05);叶片淀粉含量随着施氮量的增加呈现降低趋势,低氮处理的叶片淀粉含量显著高于高氮处理(P < 0.05);叶片NSC 含量随着施氮量的增加呈现降低趋势,低氮处理的叶片NSC 含量显著高于高氮处理(P < 0.05),中氮处理略高于高氮处理,但两者间无显著差异。
干旱和施氮的交互作用对木质部淀粉及NSC含量有极显著影响(P < 0.001),对木质部可溶性糖含量有显著影响(P < 0.05),对其他部位的NSC 含量无影响(表4)。
3 结论与讨论
本研究中探讨了干旱和施氮对苹果容器大苗水分状况、光合作用、生物量分配以及茎叶NSC含量的影响。结果表明:正常供水条件下,过量施氮导致苹果大苗水分状况变差,这会抵消氮对光合速率的促进作用,进而影响树木生长和生物量积累。在干旱条件下,施氮进一步引起水分条件恶化,限制光合作用和生物量积累,同时施氮会导致苹果大苗NSC 含量降低,可能进一步影响其对干旱的适应能力,因此在干旱条件下应减少氮肥的施用量。
干旱导致植物水分状况变差的主要表现是水势降低、水分长距离运输受阻[17],长期的水分匮乏也会使导管直径减小[18-19],导水能力降低,在本研究中干旱处理明显导致苹果大苗凌晨水势下降,导水损失率增加。氮素水平的差异也会影响到植物的水力结构。有研究结果表明,增加氮的可用性会促进导管直径增大[20-21],提高枝条导水率,但导管直径增大可能会降低水力安全性[22-23];李小刚[24] 指出,施氮导致植物水分状况变差是因为较多的氮肥施入土壤导致硝酸盐和铵离子过多,根外部水势降低,水分吸收难度增加,让植物整体水分状况变差。在本研究中,无论是正常供水条件下,还是干旱条件下,苹果大苗的水势均随着施氮量的增加呈现下降趋势,枝条导水损失率呈现上升趋势,这表明施氮量的增加对苹果大苗水分状况产生不利影响。在本研究中,正常供水条件下中氮处理的光合速率显著高于低氮处理和高氮处理,这表明正常供水条件下适量施氮有利于植物光合作用的进行。高氮处理下叶片的光合速率并未进一步提高,呈现降低趋势,这可能与高氮处理下树木的水分状况变差限制了光合作用的进行有关。
生物量的分配和形态学上的适应性变化是植物应对生长环境特征的主要机制,氮对植物生长的促进作用已被较多研究结果及生产实践证实[25]。
在本研究中,正常供水条件下,从苹果大苗不同部位的生物量等可以看出施氮对生长的积极作用,在正常供水条件下随着施氮量的增加根冠比呈降低趋势,这与前人大部分研究结果一致,即正常供水条件下氮肥更多地促进了地上部分的生长[12,14]。在干旱條件下,植物为了减少光合产物的消耗,通常会降低其生长速度,增加根的生物量分配来促进对水的吸收,同时地上部分叶片数量和面积的减少可以限制蒸腾作用,这有助于实现植株水分平衡。本研究中,干旱条件下苹果大苗较正常供水条件下生物量显著降低,根冠比明显增加。同时,在干旱条件下随着施氮量的增加,不同部位的生物量均呈现明显降低趋势,这是由于在干旱条件下施氮量增加使植物的水分状况变差,进而限制光合作用的进行,导致碳同化减少。本研究结果也表明,干旱条件下中氮和高氮处理的根冠比低于低氮处理,虽然未达到显著水平,但根生物量占比降低会进一步影响干旱条件下植株的水分吸收能力,因此施入过量的氮肥可能会减弱植物整体应对干旱的能力。
本研究结果显示,正常供水条件下,茎的木质部和韧皮部及叶片中的淀粉含量随着施氮量的增加而显著减少。植物在光合作用产物是用于生长还是用于储存上会有一个权衡[26]。在本研究中,中氮处理和高氮处理的茎木质部和韧皮部的淀粉含量和NSC 含量呈现降低趋势,随着施氮量增加茎生物量显著增加,说明随着施氮量的增加,苹果大苗会将更多的光合产物用于茎的生长而不是储存。在干旱条件下,随着施氮量的增加,茎木质部和韧皮部的可溶性糖含量显著降低,叶可溶性糖含量、淀粉含量等有降低趋势。这可能与干旱胁迫条件下气孔关闭限制了碳摄取有关[27-29]。较多研究中报道了非结构性碳水化合物在水力结构维持的渗透调节、气穴栓塞修复中的作用[30],因此施氮量增加导致的NSC 含量降低会进一步影响苹果大苗在干旱条件下水力结构的维持,影响树木的抗旱能力。
本研究中重点关注了地上部分的生长及碳素储存对水氮交互作用的响应,关于施氮和干旱处理对根系水力结构和碳代谢的影响有待进一步深入研究。此外,氮肥施入后土壤理化性质及养分含量会发生变化,这些变化对苹果大苗的生长及碳素储存的影响也有待进一步深入研究。
参考文献:
[1] WU F Z, BAO W K, LI F L, et al. Effects of drought stress andN supply on the growth, biomass partitioning and water-useefficiency ofSophora davidii seedlings[J]. Environmental andexperimental botany,2008,63(1/2/3):248-255.
[2] ZHOU X B, ZHANG Y M, JI X H, et al. Combined effects ofnitrogen deposition and water stress on growth and physiologicalresponses of two annual desert plants in northwestern China[J].Environmental and Experimental Botany,2011,74:1-8.
[3] FANG L D, ZHAO Q, LIU Y Y, et al. The influence of a fiveyearnitrogen fertilization treatment on hydraulic architecture ofPinus sylvestris var.mongolica in a water-limited plantation ofNE China[J]. Forest Ecology and Management,2018,418:15-22.
[4] CHEN C, CHU Y G, HUANG Q J, et al. Morphological andphysiological plasticity response to low nitrogen stress in blackcottonwood (Populus deltoides Marsh.)[J]. Journal of ForestryResearch,2022,33(1):51-62.
[5] 劉双娥, 李义勇, 方熊, 等. 不同氮添加量和添加方式对南亚热带4 个主要树种幼苗生长的影响[J]. 植物生态学报,2015,39(10):950-961.
LIU S E, LI Y Y, FANG X, et al. Effects of the level and regimeof nitrogen addition on seedling growth of four major tree speciesin subtropical China[J]. Chinese Journal of Plant Ecology,2015,39(10):950-961.
[6] ROSNER S, GIERLINGER N, KLEPSCH M, et al. Hydraulicand mechanical dysfunction of Norway spruce sapwood due toextreme summer drought in Scandinavia[J]. Forest Ecology andManagement,2018,409:527-540.
[7] AREND M, LINK R M, PATTHEY R, et al. Rapid hydrauliccollapse as cause of drought-induced mortality in conifers[J/OL].Proceedings of the National Academy of Sciences,2021,118(16):e2025251118. Doi:10.1073/PNAS.2025251118.
[8] HOCHBERG U, WINDT C W, PONOMARENKO A, et al.Stomatal closure, basal leaf embolism, and shedding protectthe hydraulic integrity of grape stems[J]. Plant Physiology,2017,174(2):764-775.
[9] 李娟, 彭镇华, 高健, 等. 干旱胁迫下黄条金刚竹的光合和叶绿素荧光特性[J]. 应用生态学报,2011,22(6):1395-1402.
LI J, PENG Z H, GAO J, et al. Photosynthetic parametersand chlorophyll fluorescence characteristics ofPleioblastuskongosanensis f.aureostriaus under drought stress[J]. ChineseJournal of Applied Ecology,2011,22(6):1395-1402.
[10] 李佳乐, 梁泳怡, 刘文杰, 等. 有机肥替代化学氮肥对橡胶幼苗生长和土壤环境的影响[J]. 应用生态学报,2022,33(2):431-438.
LI J L, LIANG Y Y, LIU W J, et al. Effects of manure substitutingchemical nitrogen fertilizer on rubber seedling growth and soilenvironment[J]. Chinese Journal of Applied Ecology,2022,33(2):431-438.
[11] 闫慧, 吴茜, 丁佳, 等. 不同降水及氮添加对浙江古田山4 种树木幼苗光合生理生态特征与生物量的影响[J]. 生态学报,2013,33(14):4226-4236.
YAN H, WU Q, DING J, et al. Effects of precipitation andnitrogen addition on photosynthetically ecophysiologicalcharacteristics and biomass of four tree seedlings in GutianMountain, Zhejiang province, China[J]. Acta Ecologica Sinica,2013,33(14):4226-4236.
[12] 崔婉莹, 刘思佳, 魏亚伟, 等. 氮添加和水分胁迫对红松、水曲柳幼苗生物量分配的影响[J]. 应用生态学报,2019,30(5):1454-1462.
CUI W Y, LIU S J, WEI Y W, et al. Effects of nitrogen addition onbiomass allocation ofPinus koraiensis andFraxinus mandshuricaseedlings under water stress[J]. Chinese Journal of AppliedEcology,2019,30(5):1454-1462.
[13] 赵帅翔, 张卫峰, 姜远茂, 等. 黄土高原苹果过量施氮因素分析[J]. 植物营养与肥料学报,2017,23(2):484-491.
ZHAO S X, ZHANG W F, JIANG Y M, et al. Factors leading toexcessive nitrogen fertilization on apple in the Loess Plateau[J].Journal of Plant Nutrition and Fertilizers,2017,23(2):484-491.
[14] 劉洋, 张健, 陈亚梅, 等. 氮磷添加对巨桉幼苗生物量分配和C∶N∶P 化学计量特征的影响[J]. 植物生态学报,2013,37(10):933-941.
LIU Y, ZHANG J, CHEN Y M, et al. Effect of nitrogen andphosphorus fertilization on biomass allocation and C∶N∶Pstoichiometric characteristics ofEucalyptus grandis seedlings[J].Chinese Journal of Plant Ecology,2013,37(10):933-941.
[15] 王林, 代永欣, 樊兴路, 等. 风对黄花蒿水力学性状和生长的影响[J]. 生态学报,2015,35(13):4454-4461.
WANG L, DAI Y X, FAN X L, et al. Effects of wind onhydraulic properties and growth ofArtemisia annua Linn[J]. ActaEcologica Sinica,2015,35(13):4454-4461.
[16] MITCHELL P J, OGRADY A P, TISSUE D T, et al. Droughtresponse strategies define the relative contributions of hydraulicdysfunction and carbohydrate depletion during tree mortality[J].New Phytologist,2013,197(3):862-872.
[17] 程向芬, 马晋, 赵涵, 等. 木本植物水力学结构之导管长度研究进展[J]. 植物生态学报,2018,42(6):609-618.
CHENG X F, MA J, ZHAO H, et al. Vessel length as a keyhydraulic structure in woody plants: a review[J]. Chinese Journalof Plant Ecology,2018,42(6):609-618.
[18] HEIJARI J, NERG A M, HOLOPAINEN J K, et al. Wood borerperformance and wood characteristics of drought-stressedScotspine seedlings[J]. Entomologia Experimentalis et Applicata,2010,137(2):105-110.
[19] 郑勤莹, 张国帅, 赵彬清, 等. 不同坡位水曲柳木质部解剖特征及其与气候关系[J]. 应用生态学报,2021,32(10):3428-3436.
ZHENG Q Y, ZHANG G S, ZHAO B Q, et al. Xylem anatomicalcharacteristics ofFraxinus mandshurica and relationship withclimate in different slope positions[J]. Chinese Journal of AppliedEcology,2021,32(10):3428-3436.
[20] SALA A, MENCUCCINI M. Ecosystem science: plump treeswin under drought[J]. Nature Climate Change,2014,4:666-667.
[21] 张翠萍, 孟平, 张劲松, 等. 固氮植物绿豆对核桃幼苗生长、叶片气孔气体交换及水力特征的作用[J]. 植物生态学报,2014,38(5):499-506.
ZHANG C P, MENG P, ZHANG J S, et al. Effects of a nitrogenfixing plantVigna radiata on growth, leaf stomatal gas exchangeand hydraulic characteristics of the intercroppingJuglans regiaseedlings[J]. Chinese Journal of Plant Ecology,2014,38(5):499-506.
[22] 张海昕, 李姗, 张硕新, 等. 4 个杨树无性系木质部导管结构与栓塞脆弱性的关系[J]. 林业科学,2013,49(5):54-61.
ZHANG H X, LI S, ZHANG S X, et al. Relationships betweenxylem vessel structure and embolism vulnerability in fourPopulus clones[J]. Scientia Silvae Sinicae,2013,49(5):54-61.
[23] MARKESTEIJN L, POORTER L, PAZ H, et al. Ecologicaldifferentiation in xylem cavitation resistance is associated withstem and leaf structural traits[J]. Plant, Cell & Environment,2011,34(1):137-148.
[24] 李小刚. 含盐量对土壤的水汽吸附及土壤水能量状态的影响[J]. 土壤通报,2001,32(6):245-249.
LI X G. The effect of salt content on vapor adsorption and waterpotential in salt-affected soils[J]. Chinese Journal of Soil Science,2001,32(6):245-249.
[25] 徐永刚, 宇万太, 周桦, 等. 氮肥对杉木幼树各部分养分浓度与贮量的影响[J]. 中南林业科技大学学报,2011,31(8):28-34.
XU Y G, YU W T, ZHOU H, et al. Effects of nitrogen fertilizer on indifferent parts nutrient concentration and storage of young Chinesefir[J]. Journal of Central South University of Forestry & Technology,2011,31(8):28-34.
[26] ROUX X L, LACOINTE A, ESCOBAR-GUTI?RREZ A, et al.Carbon-based models of individual tree growth: a criticalappraisal[J]. Annals of Forest Science,2001,58(5):469-506.
[27] NAKAJI T, FUKAMI M, DOKIYA Y, et al. Effects of highnitrogen load on growth, photosynthesis and nutrient status ofCryptomeria japonica andPinus densiflora seedlings[J]. Trees,2001,15(8):453-461.
[28] DIETZE M C, SALA A, CARBONE M S, et al. Nonstructuralcarbon in woody plants[J]. Annual Review of Plant Biology,2014,65:667-687.
[29] 敖红, 王炎. 干旱胁迫下云杉内源激素的响应及其气孔调节[J]. 经济林研究,2011,29(3):28-34.
AO H, WANG Y. Response of endogenous hormones andstomatal regulation of spruce to drought stress[J]. Non-woodForest Research,2011,29(3):28-34.
[30] 余斐, 張水云, 华雷, 等. 旱季补水对桉树幼龄林叶片非结构性碳水化合物的影响[J]. 中南林业科技大学学报,2021,41(5):42-48.
YU F, ZHANG S Y, HUA L, et al. Effects of dry season irrigationon leaf unstructured carbohydrates inEucalyptus young forest[J].Journal of Central South University of Forestry & Technology,2021,41(5):42-48.
[ 本文编校:闻 丽]
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
小读者(2021年2期)2021-03-29
华人时刊(2019年13期)2019-11-17
文苑(2018年22期)2018-11-19
农业环境科学学报(2017年2期)2017-03-20
现代农业科技(2017年1期)2017-03-06
现代农业科技(2016年21期)2017-03-06
红领巾·萌芽(2016年1期)2016-09-10
现代经济信息(2016年4期)2016-06-20
企业文化·中旬刊(2015年12期)2016-03-09
