
中国东部地区马尾松与黄山松群落分类及群落结构和物种多样性特征
2024-01-23 05:35范明阳方炎明
南京林业大学学报(自然科学版) 2024年1期
范明阳, 胡 萌, 杨 园, 方炎明
(南京林业大学生命科学学院,南方现代林业协同创新中心,亚热带森林生物多样性保护国家林业和草原局重点实验室,江苏 南京 210037)
群落结构主要反映群落中各种植物在空间和时间上的配置状况[1],是生态系统功能的基础,可用来预测群落的演替进程和发展趋势,揭示群落稳定性[2]。物种在功能作用和个体相对多度方面的差异是形成不同群落的基础,组成物种生物学特性的不同,导致植物群落之间结构和功能上的巨大差异,因此,研究群落物种多样性具有重要意义[3],可深度认识群落的组成结构及变化[4],探测群落的生态状况[5],认知森林的生态功能[6],为森林经营管理提供科学依据。物种多样性是群落功能的主要体现,是对一个群落结构功能复杂性的度量[7],亦是植被恢复效果的特征之一[8]。森林生物多样性和生态功能的恢复备受关注,是退化生态系统的恢复与重建的目标之一[9]。一些松属树种适应能力较强,在全球范围内被广泛用于生态恢复与植树造林[10]。
马尾松(Pinusmassoniana)是亚热带地区的主要针叶树种,分布面积位居我国针叶林面积首位[11]。该树种主产长江中下游地区,由中亚热带向北延伸,止于大别山南麓—六合和仪征一线,集中分布于江南、华南的酸性土低山丘陵(海拔800 m以下)和四川盆地瘠薄的丘陵山地(海拔1 200 m以下)[12],是森林演替中的先锋树种和主要的荒山造林树种[13]。黄山松(P.hwangshanensis)为中国特有树种,是中国东部亚热带山地森林的主要建群种[14],在中高山地区严酷的生境中,黄山松的生态功能是马尾松无法替代的[15]。两种松树同属于松属(Pinus)双微管束松亚属(Subgen.Pinus)油松组(Sect.Pinus)油松亚组(Subsect.Pinus)[16]。两种不仅亲缘关系密切,而且水平分布区重叠。但二者的分布样式不同,马尾松在低海拔山丘呈连续分布,黄山松在中高海拔山体呈现“岛状”间断分布[12, 17]。
海拔是反映冷热的综合因子,海拔梯度变化伴随着气温、湿度、光照等环境因子的规律性变化[18],环境因子沿海拔梯度的变化相比于纬度更明显[19],分布在不同海拔梯度上的植物具有不同的生态生理特征,从而决定森林群落的构成[18, 20]。因此,研究海拔梯度对物种多样性和群落结构的影响,可以认知群落对环境条件的响应机制[21],预测环境因子变化规律[22],探明群落演替规律[23]。然而,目前对于物种多样性在不同空间尺度上变化规律了解有限[24]。就重叠分布的马尾松与黄山松而言,海拔梯度与植物物种分布、群落结构的关系就显得尤为重要。
《中国植被》[25]记载了马尾松林与黄山松林。有关两种松树的天然群落研究仅有单一山体的调查报告,如黄山[26-27]和武夷山[28-29]的黄山松,南山[30]和鼎湖山[31]的马尾松等。除此之外,对于两个物种共同分析主要集中于针叶的形态[18]和抗氧化酶活性[32]、基因渐渗[33-34]以及群体遗传与物种分化[35]等方面。为了系统地探究两树种水平分布重叠、垂直分布分离、亲缘关系密切的两种松树的群落结构、物种组成和植被类型划分,本研究选择两种松树组成群落的典型分布地安徽、江西和浙江进行调查研究,拟回答以下科学问题:①马尾松群落与黄山松群落结构沿海拔梯度是否存在差异?两种群落产生差异的原因是什么?②两种群落的物种多样性有何特征?③马尾松林与黄山松林如何分类?以期为深入了解马尾松林和黄山松林的植被格局,以及科学管理保护马尾松与黄山松森林资源提供理论依据。
1 材料及方法
1.1 研究区概况与样点确定
研究区域位于中国东部亚热带地区,由于马尾松相比较黄山松分布更为广泛,笔者选择两种松树典型群落所在地作为研究区域。本研究参考了《中国植物志》、相关省区森林和植被专著、期刊论文以及中国数字植物标本馆(https://www.cvh.ac.cn)的野外调查记录等资料,并为了保证样方在空间尺度上的均匀性与代表性,设定3个选择地点的要求:①有两种松树群落存在且为天然林;②有较大的海拔梯度;③地点之间有较大的地理跨度。按以上要求,最终确定了3个省内8个地点作为研究区域,分别为:安徽省的黄山和桃花冲(鄂皖交界处);江西省的三清山、庐山和井冈山;浙江省的天目山、四明山和凤阳山。这些区域是黄山松的集中分布区,发育了典型的马尾松林和黄山松林。
1.2 样方布设及调查方法
从海拔300 m起始,以200 m为一个区间,分(300,500) m、[500,700) m、[700,900) m、[900,1 100) m和≥1 100 m 5个海拔梯度,进行样方布设,共布设样方40个。记录样方海拔、经纬度等信息(表1)。
样方调查于2020年11月—2022年3月进行。样方面积为20 m×50 m,内分10个10 m×10 m相邻接的小样格。记录样方内所有物种名,按乔木层、灌木层、草本层分别进行调查。对样方内胸径(DBH)≥3 cm的所有乔木进行每木检尺,测定其胸径、树高。对于灌木层,从10个小样格中选取2个进行调查,记录平均高度、盖度等。群落具体分层方案参考《中国植被志》研编规范:乔木层可划分为大乔木层(高度≥25 m)、中乔木层(8 m≤高度<25 m)和小乔木层(5 m≤高度<8 m) 3个亚层,灌木层可划分为大灌木层(200 cm≤高度<500 cm)和中灌木层(50 cm≤高度 <200 cm)[36]。由于马尾松与黄山松常出现在干旱贫瘠的山地,且生长环境在海拔梯度上变化较大,林下很少有草本植物,为了方便比较,本研究对草本层仅进行植被分类。
1.3 数据分析
1.3.1 物种重要值计算
基于重要值(importance value, IV)来衡量各物种在群落内重要性的相对变化[37]。乔木物种重要值为相对多度、相对优势度与相对频度的平均值;灌木层物种重要值为相对多度、相对盖度与相对频度的平均值。
1.3.2 物种多样性的度量
基于调查的40个样方物种分布数据,计算物种丰富度、Shannon指数、Gini-Simpson指数和Chao1指数[38]4种常见的α多样性指数,衡量调查样地群落内的物种多样性。物种丰富度是测定物种多样性的一个重要维度[39],但其往往会随着采样强度而变化[40]。为此绘制每个样方的稀疏曲线验证布设的样方内调查是否充分。基于文献[37]方法获取Shannon指数(H)、Gini-Simpson指数(IGS)和Chao1指数3个指标。
1.3.3 多元回归树及群丛划分
本研究采用多元回归树(multivariate regression trees,MRT)筛选影响马尾松与黄山松群落结构的关键非生物因子。因变量为每个调查样方内物种的重要值,4个非生物因子数据年均温、年降水量、海拔、纬度被纳入自变量中。年均温、年降水量来自WorldClim数据库(http://worldclim.org),空间分辨率为2.5′。地形因子初步考察结果表明,马尾松、黄山松所处地形异质性较高。其中,样方所处最高海拔为1 800 m,最低海拔为300 m,海拔高差1 500 m;纬度26.34°~30.97°N,年均温10.6~18.4 ℃;年降水量1 452~2 118 mm。
1.3.4 群丛相似性分析
为了探究沿海拔梯度马尾松、黄山松所在群落是否存在差异性,采用非度量多维标度(NMDS)和相似分析。本研究基于Bray-Curtis距离的物种矩阵来表征不同群丛物种在空间上的分布情况。基于999次置换后的相似性分析,判断群丛间与群丛内差异是否显著,R2为[-1,1]。当R2>0,表示群丛间差异大于群丛内差异;R2<0,表示群丛内差异大于群丛间差异。为了探究年均温、年均降水量、海拔和纬度对物种多样性的影响,采用线性混合效应模型来定量分析各物种多样性指标与各非生物因子是否存在线性关系。由于物种丰富度为计数数据,广义线性混合模型被用来定量分析物种丰富度与非生物因子是否存在线性关系。本研究定义该模型的残差分布遵从泊松分布,地点与每个海拔区间内所有样方被作为随机因子,故采用随机效应混合模型,并遵从以下公式:
yα=x1+x2+x3+x4+VLocation+VType+ε。
(1)
式中:yα为物种多样性指数;x为非生物因子;VLocation与VType分别为由于地点与每个海拔区间内所有样方不同所造成的方差;ε为残差。
数据分析均在R 4.2.0中进行[41]。基于R包LPSC并且结合《中国植物志》电子版查询和处理植物学名;基于R包vegan来计算物种多样性指数;基于R包mvpart进行多元回归树分析;基于R包lme 4构建混合效应模型;最后基于R包tidyverse进行数据清洗和制图工作。
1.4 植被类型的分类及命名
植被类型的划分采用数量分类的方法,数量分类采用双向指示种分析(two-way indicator species analysis,TWINSPAN),将全部样方划分为不同等级群丛。通过双向指示种分析筛选特征种,计算每个物种的特征值(fidelity value,即Ф值),利用Fisher严格检验判断Ф值的显著性(P< 0.05),如果一个物种在特定植被分类单元中出现和聚集的概率显著高于其他群丛,该物种即可确定为特征种。以特征值0.25≤Ф<0.50和Ф≥0.50为依据划分普通特征值物种和高特征值物种[36]。具有1组特征种支持的分类方案将予以接受,否则需重新调整分类方案。对分类结果进行筛选,对于仅有1个凭证样方的群丛,根据经验或相关记录,确定其是否代表1个特殊植被类型,如果没有代表性,则此群丛暂不作为最终的分类结果。
群系和群丛的命名及编码方法参照《中国植被志》的命名规范[36]:群系的科学名称由建群种或共建种的拉丁名加限定词组成;群丛的名称是由群落的建群种以及除优势层以外的其他各个层的特征种(或优势种)名称外加限定词组成。植被类型的划分通常将相对盖度、重要值或优势度75%确定为划分建群种或共建种的阈值[37],因此马尾松林与黄山松林的群落类型可分别归属于不同的植被型,如常绿针叶林和针叶与阔叶混交林等。由于样方数量只有40个,海拔变化较大且群落结构复杂,因此在数据分析中暂且不做植被型以及群丛组划分,仅对马尾松群落与黄山松群落进行初步分类。而限定词所表征的植被分类单位要包含特定群系所描述的全部植被类型,因此,马尾松林与黄山松林的科学名称以森林植被型组(Forest)为限定词,即PinusmassonianaForest Alliance和PinushwangshanensisForest Alliance。马尾松群落与黄山松群落单优势种明显,因此群丛的命名暂用Evergreen Needleleaf Forest作为限定词。由于林下草本层植物稀少甚至缺失导致草本层无特征种而无法命名的群丛,按野外调查记录,暂将样方中出现的常见种作为草本层命名。
2 结果与分析
2.1 群落物种组成及垂直结构
调查显示,40个样地内共有植物138种,隶属于56科96属。马尾松群落物种共77种,隶属于38科60属,优势科有壳斗科、樟科和五列木科,分别为4属8种、3属6种和4属5种。黄山松群落物种共有32种,隶属于35科46属,优势科有壳斗科、蔷薇科和杜鹃花科,分别为2属7种、4属6种和2属5种。
在调查中发现,马尾松林与黄山松林垂直结构变化明显,乔木层与灌木层占优势,草本层生长处于弱势。马尾松群落树高在0.5~37.0 m,18个马尾松样方乔木层独立个体共有1 123株,其中马尾松769株。小乔木层独立个体共310株,其中马尾松占40.97%,该层马尾松个体数占马尾松总体的16.51%,主要树种还有枹栎(Quercusserrata)、山鸡椒(Litseacubeba)、杉木(Cunninghamialanceolata)、甜槠(Castanopsiseyrei)等;中乔木层独立个体共781株,其中马尾松占78.49%,该层马尾松占马尾松总体的79.71%,主要树种有杉木、枫香(Liquidambarformosana)、甜槠等。大乔木层独立个体共32株,主要为建群种马尾松。马尾松群落垂直结构示例如图1a。黄山松群落树高在0.5~38.0 m,22个黄山松样方乔木层物种独立个体共1 643株,其中黄山松1 102株。小乔木层中独立个体共625株,其中黄山松占52.8%,该层黄山松个体占黄山松总体的29.95%,主要树种还有枹栎、灯台树(Cornuscontroversa)、交让木(Daphniphyllummacropodum)和云锦杜鹃(Rhododendronfortunei)等;中乔木层独立个体共997株,其中黄山松占75.33%。该层黄山松占乔木层黄山松总体的68.15%。该层主要树种有木荷(Schimasuperba)、杉木、云锦杜鹃、灯台树等;大乔木层独立个体共21株,该层主要树种为建群种黄山松;灌木层独立个体共有556株,其中黄山松占32.37%。而此层次黄山松也是构成群落的重要部分,其中有少数黄山松为群落中处于生长期的幼树,另一部分在陡峭的山地形成低矮但较为稳定的黄山松林(图1b)。

图1 黄山海拔350 m马尾松群落(a)与海拔1 150 m黄山松群落(b)Fig. 1 Photographs of Pinus massoniana at 350 m(a) and P. hwangshanensis community at 1 150 m(b) altitude in Huangshan Mountain
2.2 群落优势种重要值及物种多样性指数
稀疏曲线结果表明大部分样方调查是充分的,能够充分反映该调查区域内物种丰富度水平。马尾松群落优势种重要值为46.36~100.00,物种丰富度为2~17,Chao1指数为2~28,Shannon指数为0.31~2.01,Gini-Simpson指数为0.13~0.80。黄山松群落优势种重要值为37.55~100.00。物种丰富度为2~22,Chao1指数为2~35,Shannon指数为0.12~2.51,Gini-Simpson指数为0.05~0.87。除少数样方外,马尾松群落与黄山松群落单优势种明显(图2)。
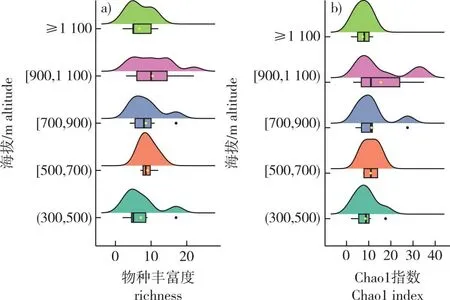
图2 不同海拔的物种α-多样性指数特征Fig. 2 Characteristics of α-diversity indices across different altitude
2.3 群落差异及影响因子分析
NMDS分析表明,马尾松群落与黄山松群落之间的结构存在差异,海拔1 100 m以上的黄山松群落与海拔900 m以下的马尾松群落之间结构几乎无相似性,不同海拔梯度下的群落间结构同样存在明显差异(图3)。

图3 基于5个海拔梯度物种丰富度的非度量多维标度(NMDS)排序图Fig. 3 NMDS ordination map based on species richness across five altitude
海拔距离差距越大,则群落结构差异越大[应力函数值(stress)为0.2,表明分组较好]。以海拔为基准,将马尾松群落与黄山松群落划分为3个群丛(图4)。基于前人对于黄山松划分的下限[17, 26],可以确定海拔887.5 m为马尾松群落与黄山松群落的分界线;而以海拔1 418 m为基准,黄山松群落结构也存在差异。

图4 基于多元回归树分析环境因子与群落 结构之间关系Fig. 4 Relationship between environmental factors and community structure based on multiple regression tree analysis
相似性分析发现,R2=0.299 4,表明群丛内差异小于群丛间差异(P=0.001),进一步表明不同海拔范围情况下,马尾松群落与黄山松群落内部也存在差异。分别使用广义线性混合效应和线性混合效应模型探究非生物因子对物种丰富度与Shannon 指数和Gini-Simpson指数的影响。通过对马尾松群落与黄山松群落总体的分析,年均降水量对群落总体物种丰富度产生影响,海拔对物种均匀度产生影响。将两种群落分开分析,则年均温对两种群落物种丰富度产生显著影响(表2)。
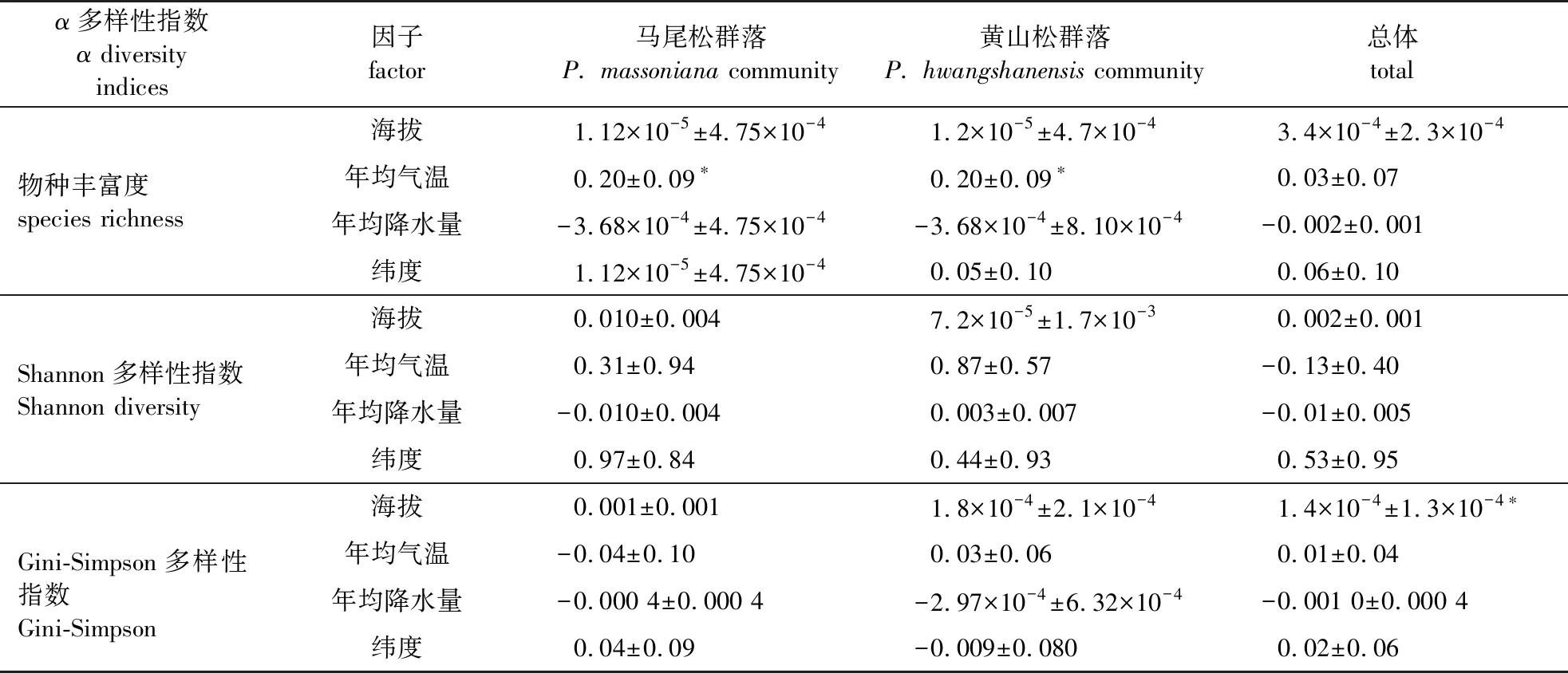
表2 非生物因子对物种α多样性指数的影响
2.4 群落分类与命名结果
通过TWINSPAN分析并对结果进行筛选,马尾松群系被划分为6个群丛,共17个凭证样方,群丛分类简表见表3;黄山松群系被划分为5个群丛,共18个凭证样方,群丛分类简表见表4。分别对两个群系分类进行命名,其中,马尾松群系的两个群丛(PM-2和PM-3)和黄山松群系的1个群丛(PH-2),按中国植被分类系统未出现草本层特征种,本研究按野外调查记录确定上述3个群丛的草本层特征种。
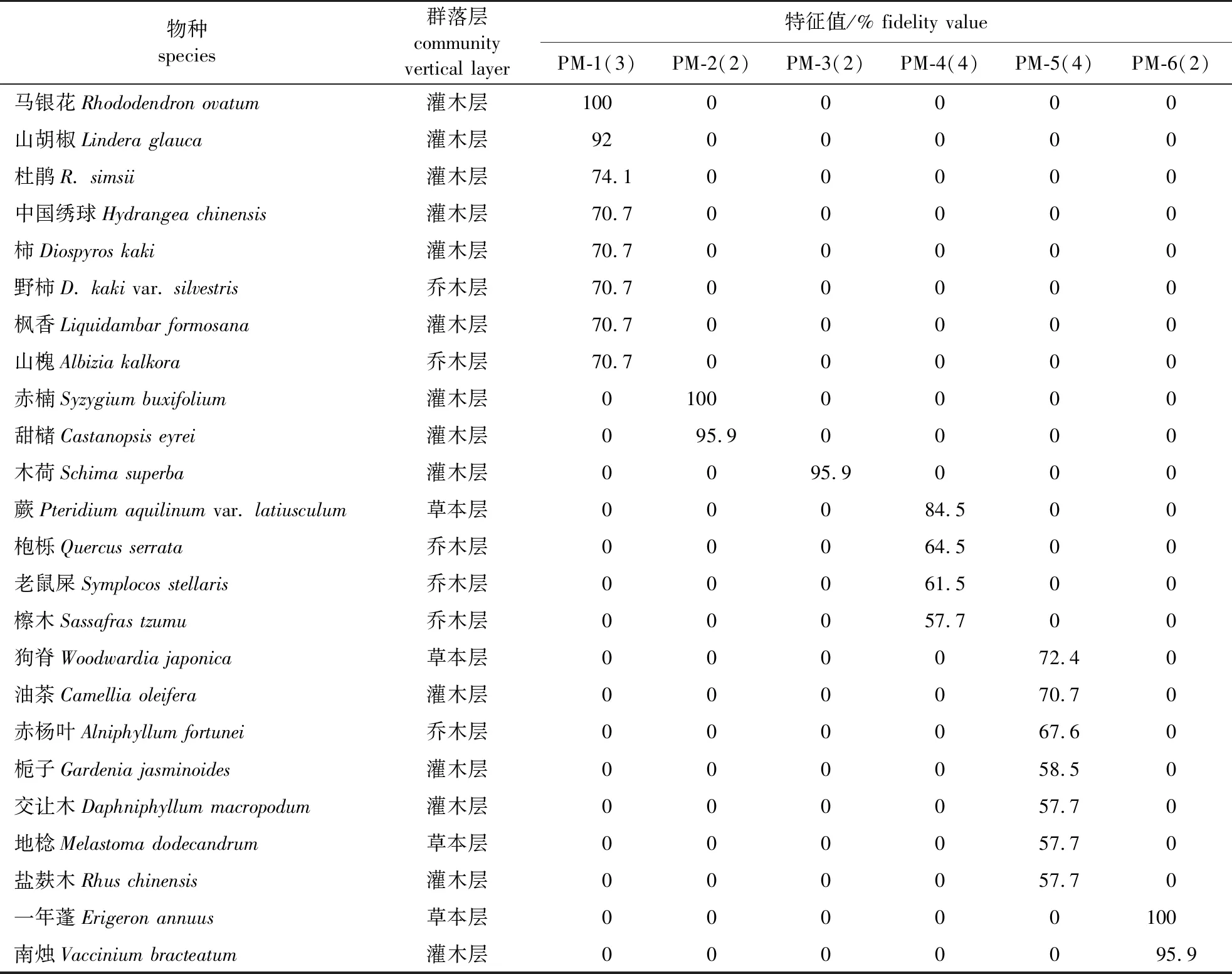
表3 马尾松群系群丛分类简表

表4 黄山松群丛分类简表
2.4.1 马尾松群系
马尾松群系(PinusmassonianaForest Alliance)共分为以下6个群丛。
1)PM-1. 马尾松-马银花-芒萁,常绿针叶林(Pinusmassoniana-Rhododendronovatum-DicranopterispedateaEvergreen Needleleaf Forest)。凭证样方为P19、P26和P27;该群丛主要分布于黄山和天目山,该群丛特征种还有山槐(Albiziakalkora)、野柿(Diospyroskaki);林下则以胡颓子(Elaeagnuspungens)有一定优势。
2)PM-2. 马尾松-赤楠-芒萁,常绿针叶林(Pinusmassoniana-Syzygiumbuxifolium-DicranopterispedateaEvergreen Needleleaf Forest)。凭证样方为P2和P7;该群丛主要分布于江西省的三清山和庐山,林下草本植物有芒萁(Dicranopterispedata)、五节芒(Miscanthusfloridulus)、狗脊(Woodwardiajaponica)等偶见种;赤楠和甜槠同时亦是该群丛的常见种,该群丛的常见种还有甜槠。
3)PM-3. 马尾松-木荷-芒萁,常绿针叶林(Pinusmassoniana-Schimasuperba-DicranopterispedateaNeedleleaf Forest)。凭证样方为P1和P3;该群丛主要分布于三清山,位于海拔500~800 m,林下草本层有芒萁、淡竹叶(Lophatherumgracile)等偶见种;常见种有鹿角杜鹃(Rhododendronlatoucheae)、檵木(Loropetalumchinense)和老鼠屎(Symplocosstellaris)等。
4)PM-4. 马尾松-老鼠屎-蕨,常绿针叶林(Pinusmassoniana-Symplocosstellaris-Pteridiumaquilinumvar.latiusculumEvergreen Needleleaf Forest)。凭证样方为P17、P18、P20和P32;该群丛主要分布于黄山低山山麓,该群丛特征种还有的枹栎、老鼠屎和檫木(Sassafrastzumu);常见种有枫香树、老鼠屎和芒萁。
5)PM-5. 马尾松-油茶-狗脊,常绿针叶林(Pinusmassoniana-Camelliaoleifera-WoodwardiajaponicaEvergreen Needleleaf Forest):凭证样方为P11、P12、P13和P14;该群丛主要分布于江西省的井冈山,位于海拔500~900 m,该群丛特征种还有赤杨叶(Alniphyllumfortunei);常见种有杉木、枫香树和芒萁等。
6)PM-6. 马尾松-南烛-一年蓬,常绿针叶林(Pinusmassoniana-Vacciniumbracteatum-ErigeronannuusEvergreen Needleleaf Forest)。凭证样方为P5和P6;该群丛主要分布于庐山,常见种还有檵木等。
2.4.2 黄山松群系
黄山松群系(PinushwangshanensisForest Alliance)共分为以下5个群丛。
1)PH-1. 黄山松-交让木-芒萁,常绿针叶林(Pinushwangshanensis-Daphniphyllummacropodum-DicranopterispedateaEvergreen Needleleaf Forest)。凭证样方为P8、P16、P29和P30;该群丛在庐山、井冈山和天目山都有分布,特征种还有多脉青冈(Quercusmultinervis)、小叶青冈(Quercusmyrsinifolia)、山矾(Symplocossumuntia);常见种有杉木、芒萁等。
2)PH-2. 黄山松-马银花-芒萁,常绿针叶林(Pinushwangshanensis-Rhododendronovatum-DicranopterispedateaEvergreen Needleleaf Forest)。凭证样方为P15、P35、P36和P37;该群丛主要分布在凤阳山中山地区,山矾在该群丛占一定优势;该群丛特征种还有甜槠和云锦杜鹃,林下草本植物有芒萁、狗脊、朱砂根(Ardisiacrenata)等;杜鹃(Rhododendronsimsii)是该群丛的常见种。
3)PH-3. 黄山松-枹栎-蛇莓,常绿针叶林(Pinushwangshanensis-Quercusserrata-DuchesneaindicaEvergreen Needleleaf Forest):凭证样方为P21、P22、P24和P31;该群丛主要分布于黄山地区,其分布海拔跨度很大,分布海拔为900~1 700 m;该群丛草本层两个特征种蛇莓和狭叶香港远志(Polygalahongkongensisvar.stenophylla)在其他群丛几乎不出现。
4)PH-4. 黄山松-荚蒾-紫萁,常绿针叶林(Pinushwangshanensis-Viburnumdilatatum-OsmundajaponicaEvergreen Needleleaf Forest):凭证样方为P9和P10;主要分布于庐山,该群丛常见种有山橿和杜鹃等。
5)PH-5. 黄山松-山胡椒-蕨,常绿针叶林(Pinushwangshanensis-Linderaglauca-Pteridiumaquilinumvar.latiusculumEvergreen Needleleaf Forest):凭证样方为P34、P38、P39和P40;主要分布于桃花冲,灯台树亦是该群丛的特征种。
3 讨 论
黄山松和马尾松群落结构沿海拔梯度的变化产生显著的差异,且海拔1 100 m以上的黄山松群落与海拔900 m以下的马尾松群落之间物种分布没有重叠。原因是海拔梯度的异质性导致物种分布替代[42],从而影响植物的分布及群落的构成。经调查发现,马尾松与黄山松主要处于各自群落的中乔木层以及小乔木层中,其林下有阔叶树种的幼树幼苗存在,虽然马尾松与黄山松目前仍占有较大的优势,但随着群落的演替,其当前的优势地位将逐渐被阔叶树种取代。而处于更高海拔、环境恶劣的地方所形成低矮的黄山松林,其他物种在此难以生长,林下物种极为稀少,因而常形成较为稳定的群落。
Wang等[43]基于大尺度的研究表明,气候因子对物种丰富度产生很大影响,尤其是环境能量因子影响显著。本研究结果显示,马尾松群落物种丰富度随着年平均气温的增加而显著增加,黄山松群落亦是如此。这表明马尾松与黄山松林下多生长喜干热的阳性物种。对两个群落总体分析时,年平均降水量成为影响两个群落总体物种丰富度的显著因子,与方文静等[44]对华北地区落叶松林的研究结果相似。但与之不同的是,本研究结果显示两个群落总体物种丰富度随年均降水量的增加而减少,这意味着在降水量大的环境中,两个群落中的物种生长反而受到抑制[45],这也许与研究区域的不同有关,其地貌、气候等存在很大差异,而在大尺度空间内,地貌、气候等间接性生态因子是决定森林物种丰富度的主导因子[46]。前人诸多的研究表明,水分条件是植物群落物种丰富度的重要影响因子[39, 47-48],但不能忽视空间、演化历史和物种间相互作用等潜在因素的影响[49]。前人对全球地理格局的研究结果表明,物种丰富度随着经纬度的增加而显著降低[39, 48],本研究结果表明纬度对马尾松群落、黄山松群落以及两个群落的总物种丰富度影响并不显著,这可能是由于调查样方数量不够多以及样方的纬度差异不够显著的原因。同时,本研究表明马尾松与黄山松群落结构垂直分布的明显差异,物种多样性会随着海拔梯度的变化产生响应,而结论也确是如此,海拔梯度的变化对两个群落总体Shannon-Wiener指数和Gini-Simpson指数的影响是显著的。对两个群落总体分析表明,Shannon-Wiener指数和Gini-Simpson指数随海拔的升高而显著升高。Shannon-Wiener指数能较好地反映物种丰富度和群落均匀度,Gini-Simpson指数主要反应优势种在群落中的地位,随着海拔的升高,黄山松更集中分布,这进一步解释了多元回归树的分析结果。而朱德煌[50]对戴云山的黄山松群落研究的结果表明随着海拔的升高Simpson指数变大,笔者分别分析两种群落,结果发现海拔的变化对Shannon-Wiener指数和Gini-Simpson指数产生的影响并不显著,这可能是因为研究尺度大小以及地点不同导致的差异。事实上,黄山松的分布更多受到山体高度的限制,使其不能达到应有的分布高度,且生境的异质性对黄山松群落结构产生很大的影响,黄山松分布区域内的山体海拔差异很大,因此导致在黄山松群落内Gini-Simpson指数随海拔的变化不显著。
在自然界中,任何植物都占有一定的地理空间并具有一定的生态位[51]。黄山松林垂直分布的下限与马尾松林垂直分布的上限沿着海拔梯度上有密切的关联,两种群落在分布区域内形成占据不同海拔梯度的天然格局。这是由于黄山松垂直分布的下限因地理位置不同而发生变化,一般随纬度的增加,分布下限高度降低;随经度的下降,分布下限高度升高[17]。如凤阳山黄山松林生长于海拔1 000~1 800 m范围内[52]、衡山黄山松生长于海拔1 200~1 300 m范围内及大别山海拔600~1 700 m等[17]。MRT结果不仅表明了沿海拔梯度群落结构存在的差异,而且进一步说明海拔是群落间结构产生差异的主要因子。MRT结果显示海拔887.5 m为群落的分界线,根据前人研究[17, 26],可以确定此界限为马尾松群落与黄山松群落的分界线,但此界限并非两个物种的分界线,因为已有研究发现马尾松林与黄山松林在垂直过渡带上存在天然杂种域[53]。李淑娴[54]通过对黄山地区马尾松林与黄山松林研究发现,其过渡带间松树种子饱满度低可能与两物种存在种间繁殖障碍有关,而此过渡带区域为海拔800~1 100 m。这也是本研究结果中海拔低于前人对黄山松划分的分布下限的原因。综合NMDS和MRT分析,本研究得出的界限与前人研究的马尾松林与黄山松林之间的过渡带基本吻合。该结果对马尾松林与黄山松林及其杂种域的进一步探究提供了线索与参考价值。而海拔梯度的变化实质是温度、降水等环境因子的综合反应。童再康等[17]关于黄山松分布下限的主要气候因子研究显示,北起安徽大别山向南至台湾玉山,年降水量在1 560.0~2 906.4 mm之间,而年均温变化为12.2~17.0 ℃,在这个分布区内,降水量是黄山松分布的主要限制因子。
中国植被分类系统的植被型组及植被型的划分基于群落的生态外貌,而群系和群丛组等的划分基于群落结构及物种组成[25],由于马尾松群落和黄山松群落海拔跨度较大,且出于样方数量的限制,给植被型、群丛组及群丛的划分增加了难度,而选择生境差异较大的群落进行调查,能更加全面地反映我国多样化的植被类型。本研究仅对两个群落进行初步分类,旨在甄别出群落的特征种,并了解群落的物种组成。对于马尾松群落与黄山松群落的植被型及群丛组的划分还需更多的样方数据支持,从而获得完整的群落外貌、分层特征等信息。MRT分析将40个样方划分3个群丛,但此结果并不能较为完整地代表此区域内马尾松与黄山松实际的群落分布类型。将黄山松群落以海拔1 418 m为界限划分为两个群丛,这与前人的研究结果相似,如刘金福等[56]对戴云山黄山松群落的研究表明,随着海拔的升高黄山松群落逐渐趋近于纯林,且在1 400~1 600 m海拔区生长表现突出。而在实际调查中发现,黄山松纯林包括高大的黄山松纯林和低矮的黄山松纯林,前者生长于土壤条件相对较好的环境,其林下植物稀少,黄山松高大且整齐并占绝对优势;而后者生长于陡峭的山地及岩壁上呈低矮灌木状且较为破碎,样方调查较为困难,但从林相观察黄山松确实占绝对优势。因此,关于马尾松林与黄山松林的分类和命名等问题需要更多的凭证样方以补充完善及加以修订。
猜你喜欢
中国水土保持科学(2023年6期)2024-01-05
水土保持研究(2022年1期)2022-12-27
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
黄山学院学报(2017年5期)2017-11-03
山西教育·幼教(2017年11期)2017-04-13
武夷学院学报(2015年3期)2015-07-18
山西大学学报(自然科学版)(2014年2期)2014-05-10
