
陆地环境中纳米塑料毒性效应的研究进展
2024-01-20 07:31:54刘安吴昊何贝贝
生态环境学报 2023年11期
刘安,吴昊,何贝贝
深圳大学化学与环境工程学院,广东 深圳 518060
自从1907 年,第一种塑料材料被合成,塑料产品已广泛应用于材料包装(Castle,1994)、医疗器材(Li,2022)、农业用品(Berenstein et al.,2022)、建筑材料(Nyika et al.,2022)、电子电器(Jia et al.,2022)等方面。塑料的稳定性使得它们在海洋、淡水及陆地栖息地等自然环境中积累而很难降解(Rillig,2012;Topcu et al.,2013;Wagner et al.,2014;Geyer et al.,2017)。因此,塑料废物已在环境中无处不在(易浪等,2022)。
2004 年,Thompson et al.(2004)首次提出了“微塑料(MPs)”一词,这些由大型塑料分解形成的塑料碎片对生态环境构成了严重的威胁。MPs 一般指粒径小于5 mm 的塑料碎片,随着MPs 在环境中的继续分解,逐渐会形成粒径更小的碎片,这些粒径更小的塑料碎片被称为纳米塑料(Nanoplastics,以下简称NPs)。NPs 作为新型的污染物(图1),在世界范围内已受到广泛的关注。针对NPs 的粒径范围,一些学者将其尺寸上限设定为1 000 nm(Cole et al.,2015;Rochman et al.,2016;Ter Halle et al.,2017),而其他学者则设定为100 nm(Koelmans et al.,2015;Gigault et al.,2018)。相较于MPs,NPs 的粒径更小,对其他生物可能具有更大的毒害作用,因而成为了全球关注的污染物。
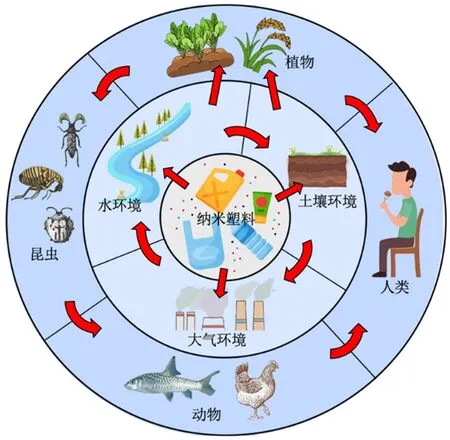
图1 NPS在环境中的分布及归趋Figure 1 Distribution and fate of NPs
NPs 的特点是尺寸极小,形状不规则,并且在成分和表面化学方面具有异质性(Koelmans et al.,2015)。这种特性使得难以使用传统的分离MPs 的方法从环境中收集NPs(Zhang et al.,2020)。NPs较小的尺寸还使其容易形成聚团、也易被生物细胞摄取,并且其中的化学添加剂较易快速释放出来(Chen et al.,2020)。这使得NPs 可以将各种病毒、细菌、重金属、有机污染物从环境中输送到生物体内,从而促进有毒污染物在生物体内累积以及转移(Zhang et al.,2022a)。图1 给出了NPs 在环境中的分布及归趋。目前,由于水体样品容易采集,且针对水体样品中的NPs 进行检测较易进行,所以NPs 对水生环境及水生动植物等影响的研究较多,而NPs 对陆地生物产生的毒性效应及对陆地环境影响机制的研究尚浅。据此,本文旨在综述近年来NPs本身的毒害效应(包括其中的添加剂)、降解产物和与其他环境污染物(有机污染物、无机污染物)耦合后的复合毒性以及NPs 对陆生植物、陆生动物以及土壤环境产生的综合影响方面的研究,并提出了未来的研究重点和方向。
1 纳米塑料污染物
1.1 环境中的纳米塑料污染物
目前在水生环境、陆地生态系统和大气中均能检测到NPs 的存在(Alomar et al.,2016;Allen et al.,2019;Zhang et al.,2019)。环境中的NPs 主要来源于两个方面:初级NPs 和次级NPs。初级NPs指在制造时体积微小,通常由聚乙烯或聚苯乙烯组成,主要运用于含有微珠的防水涂料、生物医学产品和个人护理产品,例如油漆、面部磨砂膏、牙膏等(Fendall et al.,2009),另一个用途则用于作为建筑材料。次级NPs 主要来源是大型塑料或微塑料的机械磨损(El Hadri et al.,2020)、光化学氧化(Yousif et al.,2013)、生物降解(Narancic et al.,2018)等过程,导致塑料破碎成较小的NPs 颗粒(Dominguez-Jaimes et al.,2021;Yu et al.,2021a)。
1.2 环境中纳米塑料的赋存特征
许多研究表明,NPs 本身具有毒害性。吸附在细胞上的NPs 可以通过破坏细胞膜而造成毒性(Lian et al.,2021)。例如,聚苯乙烯纳米塑料(PSNPs)可以紧密地粘附在生菜根系的细胞膜上,造成高毒性。原因是PS-NPs 颗粒与细胞膜之间的静电吸引力影响了水通道蛋白的运输过程和膜的疏水性,导致膜内外渗透压失衡,最终导致膜破裂(Sun et al.,2021);其次,当NPs 进入细胞内,会造成细胞损伤、对细胞内分子进行干扰和引起氧化应激(Liu et al.,2022b)。NPs 会使细胞产生活性氧自由基(ROS)。例如,高等植物和藻类细胞会在NPs的诱导下造成氧化应激,ROS 通过降低合成代谢程度来影响植物的能量代谢(Lian et al.,2020b)。ROS的升高也会影响植物的基因毒性。例如,微核(MN)测试显示,PS-NPs(100 nm)增加了绿豆根细胞的MN 频率(Chae et al.,2020),这表明NPs 引起的遗传毒性潜力增加。上调基因本体(GO),包括生物合成和代谢过程,根表皮细胞被NPs 改变,导致根成熟区肿胀和对缺水相关基因的下调,从而降低了植物的鲜重(Chae et al.,2020)。
除了NPs 本身的毒性,从NPs 中浸出的化学添加剂也是引起毒性反应的原因之一(Liu et al.,2020)。在塑料生产过程中会添加各种添加剂,如增塑剂、阻燃剂、抗氧化剂、紫外线稳定剂、热稳定剂、滑爽剂、固化剂、杀菌剂、颜料和其他物质等(Hahladakis et al.,2018)。塑料当中的这些添加剂在受到外力(光、风、雨、机械)作用时会释放出来(Lehner et al.,2019)。这些添加剂可能导致内分泌紊乱,进而影响流动性、繁殖、发育和致癌(陈蕾等,2021;Ding et al.,2022)。此外,这些添加剂增加了环境中的溶解性有机碳(DOC)量,扰乱了环境中的碳循环(Vom Saal et al.,2005;Hahladakis et al.,2018)。
另外,NPs 和其他环境污染物(有机、无机)耦合后会产生复合毒性,如多环芳烃(Bakir et al.,2014;Mai et al.,2018)、抗生素(Hahladakis et al.,2018)、重金属(Yousif et al.,2013)、农药(Tong et al.,2021)等。所复合的化学物质浓度受到NPs 在环境中的暴露时间、NPs 的老化程度、以及环境介质特性的影响(Holmes et al.,2012)。例如,老化的NPs颗粒吸附的微量金属较多。NPs 的老化也可以增强塑料颗粒表面的反应活性,如光氧化风化会产生含氧基团,从而增加NPs 的极性,表面电荷、粗糙度、孔隙度和亲水性等(Tong et al.,2022)。三氯羟基二苯醚(Triclosan)是一种剧毒物质,实验证明,Triclosan 可以富集到NPs 上,从而通过食物链积累,给动物和人类健康带来潜在风险。由此可知,NPs 不利影响可能是由其本身、化学添加剂的释放,吸附的其他化学物质或是这些情况的组合引起的(Fraser et al.,2020)。这就使得NPs 的毒性效应不但受到其本身毒性的影响,还与其内部含有的添加剂和外部吸附的污染物有关,且其吸附特性又受到外界自然环境的影响,因此具有非常复杂的毒性特征。
2 纳米塑料的生物毒害效应
2.1 纳米塑料对陆生动物的毒害效应
本文所研究的陆生动物是指包括人类在内的哺乳类、鸟类、爬行类和两栖类动物。NPs 主要通过空气吸入、口腔摄入等途径(Cedervall et al.,2012;Mattsson et al.,2015;Kosuth et al.,2018)进入陆生动物体内,并且会通过体液运输易位至其他的组织和器官,从而对动物体造成不同的毒害效应。本节以NPs “进入”体内和在体内“易位”这两种过程对陆生动物体产生的毒害效应进行分别论述。
2.1.1 纳米塑料进入陆生动物体内产生的毒害效应
2.1.1.1 通过吸入产生的毒害效应
吸入是陆生动物能直接接触到NPs 的最主要途径(Dris et al.,2017)。进入体内的NPs 的一个来源是纺织品(如衣服和床上用品)(韩超然等,2022)。每件衣服每次洗涤约释放1 900 根纤维(属于塑料的一类)(Browne et al.,2011);另外的来源则为建筑物材料、工业排放、颗粒悬浮、交通中释放的颗粒等。这些塑料颗粒很容易通过呼吸进入并累积到人的气管、鼻腔和肺部(Prata,2018)。通过尸体解剖可知,人体肺组织中能检测出NPs 的存在,20 个肺部组织样本中的13 个中观察到聚合物颗粒和聚酯纤维(Amato-Lourenco et al.,2021)。吸入到体内的塑料颗粒会根据生物体结构和颗粒特性,在鼻腔和肺部等位置沉积(Dris et al.,2016)。与其他微米和纳米尺寸的颗粒一样,较小和较低密度的NPs 颗粒将更有可能到达肺部的下气道和肺泡区域,且因为它们的高表面积和高渗透潜力,NPs 难以从肺系统中去除(Deville et al.,2015;Dris et al.,2017;Prata,2018)。而较大的塑料颗粒可以被鼻腔和气管内等粘膜或纤毛清除(Dong et al.,2020),较少到达肺部。其实,达到肺部的NPs 不仅仅局限于特定的肺部区域,而且还被发现分布在整个肺部。使用人类肺上皮细胞来研究肺损伤与PS-NP 之间的联系时发现,随着PS-NPs 浓度的增加,细胞活力显著降低(Yang et al.,2021)。PS-NPs 可以通过耗尽紧密的结蛋白来降低上皮电阻。研究结果还表明(Yang et al.,2021),经过PS-NPs 处理的细胞增加了基质金属肽酶表面活性剂蛋白A 的水平。这表明暴露于PS-NP可能会降低肺部的修复能力并导致组织损伤。另外,空气中的NPs 可以被吸入并通过分支气道沉积在肺泡中,在那里它们首先与肺表面活性剂(LS)相互作用。LS 在降低肺泡空气-水界面的表面张力和参与先天宿主防御粒子入侵方面起着双重作用(Li et al.,2022)。这种局部颗粒沉积引起的氧化应激,可能会导致慢性肺部炎症,甚至可能增加患癌风险(MacNee,2001)。
职业教育的目标是帮助学生掌握一技之长,从而让他们在竞争激烈的社会中占有一席之地。因此, 教学中,教师要结合企业的生产需求,不断引导学生,从而提升学生的实践能力、学习能力等。如果有学生不知道怎样学习,也不知道怎样运用知识,他们学习就变得没有意义,他们也会被竞争激烈的社会淘汰。为了减少这一情况的出现,教师要大胆创新,鼓励学生动脑,从而让学生在动手动脑的过程中提升能力。
目前,对NPs 对肺部的影响研究仍具有一定的局限性。虽然吸入是最有可能的暴露途径,但不能排除一些NPs 可能通过全身易位到达肺部。此外,针对NPs 的吸入对陆生动物的危害主要通过体外模型来检测。但NPs 对动物鼻腔、气管、肺部的真实影响尚未可知,而且NPs 如何导致肺部炎症的机理还未曾得知。未来的研究需进行NPs 动物实验,可以通过控制环境中NPs 颗粒浓度、种类等实验去探究NPs 对生物体的真实影响、如何在肺部释放化学物质、炎症等致病机理,以评估NPs 通过吸入对生物体造成的危害。
2.1.1.2 通过摄入产生的毒害效应
NPs 会漂浮或沉积在陆生动物生长的环境中(Mofijur et al.,2021),这会使得陆生动物有摄入NPs 的风险(Reisser et al.,2014)。大多数研究都是使用PS-NPs 完成,以评估其对肠道的损伤和毒性。胃肠道-塑料颗粒的体外模型研究显示,NPs 具有穿透肠道上皮细胞层的能力(Mattsson et al.,2015)。PS-NPs 大多数会附着在肠道黏膜中,其量分别在前肠、中肠和后肠区域有 (216×106±40×106)、(262×106±22×106) 和 (720×106±251×106) 粒,不少NPs 会穿过黏膜到达肌肉层,从而产生更难预计的风险(Clark et al.,2022)。在这些细胞研究中,使用原始和荧光纳米聚苯乙烯颗粒表明没有消化屏障损伤或上皮细胞毒性。然而,羧化(-COOH)或氨基化(-NH2)的NPs 会破坏肠道屏障功能(Della Torre et al.,2014),NPs 更容易通过内吞作用进入上皮细胞,以及对细胞产生更致命的毒性(Rubio et al.,2020)。此外,PS-NPs 与其他环境污染物(如金属)的混合物显著增加了细胞的吸收和毒性(Cedervall et al.,2012)。肠道中的微生物会产生使多糖降解的特异性酶,这使肠道中的微生物具有降解原本动物体不可消化的碳水化合物的功能(Dam et al.,2019),但NPs 扰乱了肠道氧化和炎症平衡,过量的ROS 可能导致肠上皮细胞的脂质过氧化和蛋白质氧化,从而破坏了肠道微生物群、免疫蛋白活性、营养吸收和损害肠道功能(Li et al.,2020a)。
目前,大多数关于NPs 对陆生动物消化系统毒性的研究仅停留于探究肠道上皮细胞的氧化应激影响和NPs 在肠道内的聚集情况,没有更深层次的研究NPs 破坏肠道屏障功能的机理、NPs 使肠道炎症的致病机理等。其次,尚未有针对NPs 在生物体内摄入-消化-排泄整个系统的研究。因此,往后的研究需注重NPs 是否在肠道皮下细胞也产生毒害效果、产生毒害作用的原因以及NPs 在肠道的累积与排泄的定量分析,为探究NPs 对生物体整个消化系统的毒害效应提供理论依据。
2.1.2 纳米塑料在动物体内易位产生的毒害效应
NPs 主要从胃或肺向生物体的其他组织或器官易位(Lu et al.,2016)。例如,在服用PS-NPs 后,小鼠肠道细胞的通透性增加,NPs 更容易进入到动物体的运输系统,从而向身体的各个部位转移(Li et al.,2020a)。在斑马鱼的培育实验中,口服了NPs后能在胚胎中检测到NPs 的存在,进而对后代产生影响(He et al.,2022)。NPs 进入其他器官中会逐渐积聚,包括肝脏,肺,肾,脑和生殖系统。而且,当母体肺部接触NPs 后,母体心脏、脾脏和胎儿胎盘、肝脏、肺、心脏、肾脏和大脑中均有NPs 积累,并且在妊娠晚期从肺部向全身易位(Cai et al.,2019)。这表明NPs 毒性可能不仅限于初始接触部位(Lehner et al.,2019)。表1 总结了NPs 对生物体各个器官的毒性效应。从表1 可以看出,NPs 很可能造成血管堵塞;诱使精子头畸形、无尾、小头、杂交体丢失、卷曲或精子肿胀;诱使碱性磷酸酶(ACP)、酸性磷酸酶(AKP)、苯并氧化酶(PO)等免疫相关酶的活性下降;NPs 还会显著降低细胞内钙水平、线粒体膜电位、细胞代谢和心肌细胞的收缩力,这最终可能导致体内致命的心力衰竭。
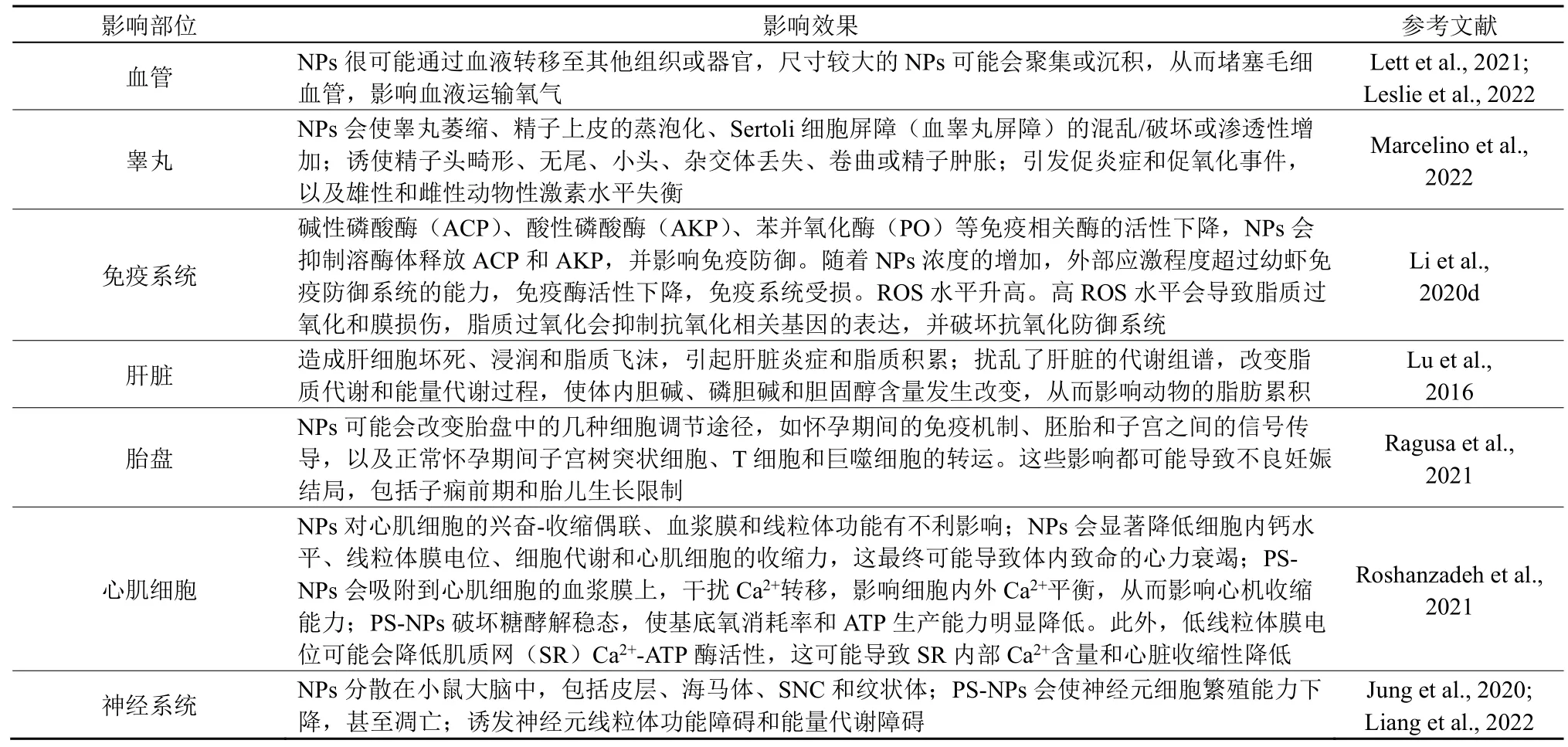
表1 NPs 的易位及对各个器官的毒理效应Table 1 Translocation of NPs and toxic effects on organs
目前,尽管NPs 对陆生动物各个部位所造成毒害作用已有充足证据,但有关NPs 如何对各组织和器官造成毒害影响的机理、NPs 在各组织和器官累积量等研究尚且不足,对NPs 如何引发生物的应激反应,NPs 通过何种途径在生物体内运输的过程均未被探明。据此,今后的研究需要更深入地从细胞层面上探究NPs 的影响机制、定量分析NPs 的积累量、不同浓度、种类NPs 在不同组织和器官造成的差异等问题。
2.2 纳米塑料对植物的毒害效应
NPs 对植物的影响主要体现在使生物量降低、植株高度降低、含水量降低、叶绿素含量降低、光合作用系数下降等方面(Sridharan et al.,2021;Zhouet al.,2021a;Liu et al.,2022a)。表2 总结了NPs对各类植物的毒理效应的研究,分析了不同类型、不同尺寸的NPs 对各种植物的影响效果。例如,NPs可以抑制拟南芥的生长(Yin et al.,2021),抑制程度取决于NPs 颗粒的电荷和浓度。带正电荷的NPs颗粒对拟南芥的毒性更大,带正电荷的NPs 会明显降低拟南芥体内的4 种过氧化物酶浓度,导致过氧化物产量增加,最终导致氧化损伤(Sun et al.,2020)。NPs 附着在生菜根表面会导致根毛孔堵塞(Sun et al.,2021;Lian et al.,2021)。当环境中存在邻苯二丁酯(DBP)时,PS-NPs 和DBP 通过van der Waals 力相互作用,增加了对生菜的负面影响。PSNPs 和DBP 的组合不仅影响了生菜的生长,还降低了光合作用参数,使得微量营养素和必需氨基酸的量显著减少。在9 种必需氨基酸中,亮氨酸、异亮氨酸和纈氨酸属于支链氨基酸(BCAA)(Wu et al.,2020),这可能会影响蛋白质的生物合成和能量产生,这3 种氨基酸的下降能够阻碍相应蛋白质的生物功能,进而导致生菜整体的营养质量下降(Lian et al.,2021)。在NPs 的影响下,植物体也会产生氧化应激效应,植物体的ROS 含量和抗氧化系统会发生变化,通过抗氧化酶活性反映出植物氧化受损的情况(Li et al.,2021a)。ROS 和抗氧化酶是植物诱导的相对防御基因表达的信号分子。在相关研究中,通过检测抗氧化酶活性和丙二醛(MDA)含量来评估NPs 对植物的氧化损伤(Lian et al.,2020b)。低浓度(0.1 mg·L-1)的聚苯乙烯纳米塑料(PS-NPs)会增强ROS 刺激酶编码基因的表达,从而增加各种抗氧化酶的活性(朱旭等,2022)。MDA 浓度的微小变化反映了植物抗氧化系统的有效性(Liu et al.,2022b)。然而,高浓度(100 mg·L-1)的PS-NPs 可能会削弱抗氧化防御系统,导致失衡,造成对植物细胞的损害(Mittler et al.,2011);在NPs 对水稻培养影响的实验中,随着NPs 浓度的增加,抗氧化酶的活性先升高后降低,MDA 也会随着浓度的增加而积累(Li et al.,2020b)。此外,NPs 还可以进入植物细胞,使用NPs 的荧光标记技术来测量它们被植物根的吸收。实验证明,烟草BY-2 细胞通过内吞作用吸收PS-NPs,进而造成植物细胞壁破裂等问题(Zhou et al.,2021a);NPs 对植物的部分遗传指标也有重要影响,包括抗氧化剂、RNA、基因转录、蛋白质表达和代谢物积累(Li et al.,2020e)等。相关研究表明,PS-NPs 会通过调节植物的RNA 转录和蛋白质表达来调节萜类和类黄酮生物合成途径,使这些抗氧化类化合物的合成量降低,造成植物的氧化损伤(Lian et al.,2020b)。由此可见,NPs对植物的生长具有不同程度的抑制效果,并且NPs被吸收转移到植物根系的现象表明,NPs 可以沿食物链迁移。如果是农产品类的植物,可能最终对人类健康构成威胁(Wang et al.,2021)。

表2 NPs 对各类植物的毒理效应Table 2 Toxic effects of NPs on plants
值得关注的是,尽管已有研究表明NPs 对植物体生长具有抑制作用(Lian et al.,2021;Sun et al.,2021;Liu et al.,2022b),但也有研究发现NPs 能够降低其他共存毒性污染物(如重金属)对植物的抑制作用或是对个别植物的生长具有明显的促进效果。例如,NPs 可以吸附重金属,使其在环境中的浓度减小,从而降低该重金属对植物的毒害作用。另外,NPs 本身可以参与植物体内的代谢过程,改善光合作用,因而促进植物的生长。Lian et al.(2020b)的研究表明,Cd 和PS-NPs-Cd 处理都明显导致小麦幼苗生长迟缓。但是,单独使用Cd 处理时,小麦幼苗的干生物量、叶绿素含量和根冠比等指标降低较快。相比之下,当PS-NPs 和Cd 共存时,这些指标降低的效果却变得不明显了。单独Cd 处理对小麦生长的抑制比联合处理(Cd-PS-NPs)更严重。这表明PS-NPs 的存在部分缓解了Cd 对小麦的毒性。NPs 能够减轻小麦中Cd 毒性的原因可能是NPs 将Cd 吸附,使环境中的Cd 浓度降低(Lian et al.,2020b)。而当Cu 和NPs 共存时,小麦植株高度比单独有Cu存在时有所增长。在之前的研究中发现内化的PSNPs 可以显著提高小麦叶绿素含量和氮同化效率,这可能是 NPs 使 Cu 的毒性缓解的重要原因(Davarpanah et al.,2015)。在植物体内,PS-NPs 可参与3 个过程,即碳代谢途径、磷酸戊糖途径和光合作用天线蛋白的合成。这3 个过程是植物进行光合作用过程中至关重要的碳固定途径(Moore et al.,2016)。此外,NPs 的存在也使得氧化磷酸化过程被显著促进。这是一种将太阳能转化为三磷酸腺苷(ATP)和还原型辅酶(NADPH)的过程(梁嘉伟等,2022)。这表明NPs 可以促进光系统期间的光转换速率(Lian et al.,2020b)。此外,植物在暴露到PSNPs 后,碳水化合物分解代谢和单生物碳水化合物分解代谢过程等能量代谢途径被激活,通过糖酵解,柠檬酸循环,糖原分解,磷酸戊糖和糖醛酸途径为植物生长提供必要的能量(Pidatala et al.,2016)。因此,NPs 能够改善光合作用过程并加速能量代谢途径,这可能是NPs 促进植物生长的重要原因。
目前,关于NPs 对植物生长影响方面的研究主要关注的是NPs 本身的毒害效应,但针对其降解过程及环境中与其他污染物的协同毒性(Yin et al.,2021)相关研究尚未深入开展。相比之下,环境中的NPs 经过光照或是生物降解,可能成为重金属,细菌和病原体良好的载体(Dominguez-Jaimes et al.,2021)。据此,未来的研究需更加关注降解后的NPs在实际环境中对植物的毒害效应。另外,目前NPs对植物毒性效应的研究大多数局限于NPs 对植物的直接影响,未考虑到NPs 对土壤的污染,从而对植物产生的间接毒害作用(Huang et al.,2022)。因此,通过研究NPs 对土壤环境-陆生植物体系的综合效应而非单独地研究NPs 对植物体的影响,能更全面地评估NPs 的毒害效应;此外,目前研究对生物体影响的实验中所使用的塑料种类大多为聚苯乙烯(PS)塑料颗粒(Sun et al.,2021;Spano et al.,2022;Zhang et al.,2022a),而其他塑料种类对植物的毒害作用研究严重不足,往后的研究需针对更多的NPs 种类,如聚氯乙烯(PVC)、聚乙烯(PE)等以评定各类NPs 的毒理效应。
3 纳米塑料对土壤环境的影响
3.1 纳米塑料在土壤中的环境行为和迁移
尽管目前针对NPs 在土壤中的浓度水平等相关研究较少(Nam et al.,2022),但针对NPs 在土壤中的环境行为和迁移等已有较多报道。土壤中的NPs 颗粒粒径使其有穿过土壤的能力,容易向地下运输(Yu et al.,2021b)。此外,与微生物和溶解有机物的相互作用可能会使NPs 更具亲水性,从而促进其向地下运输。O’Connor et al.(2019)研究了初级NPs 的垂直迁移。他们发现,NPs 的运输受颗粒大小和聚合物类型的影响,最小的聚乙烯颗粒移动最远。而与颗粒尺寸相似的聚丙烯相比,聚乙烯的运输能力也较强。同时,NPs 在土壤中也可通过地下水流进行迁移(Li et al.,2020c)。另外,经过降解的NPs 比原始NPs 具有更强的迁移能力。Liu et al.(2019)将原始的PS-NPs 暴露在紫外线(UV)辐射和臭氧下,并观察到UV 辐射和臭氧导致NPs微球表面形成环氧/羟基、羰基和羧基。这些被氧化的NPs 在土壤的迁移过程中,比原始NPs 具有更强的移动性。Yan et al.(2020)使用了自然老化的NPs进行了土壤浸出测试,他们发现这些NPs 被高度光氧化,并且与土壤中的矿物质杂凝聚,这增强了NPs在土壤中的运输。此外,土壤中存在的腐殖酸也大大促进了NPs 在土壤中的迁移(Yan et al.,2020)。这些结果证实,NPs 的降解、与土壤中矿物质凝聚和腐殖酸的存在等因素都会使得NPs 在土壤环境中的流动性增强。
3.2 纳米塑料对土壤的危害风险
NPs 可以改变土壤物理性质,包括土壤容重、松软程度和保水性能等(Zhou et al.,2021b)。例如,在聚醚砜(PES)、聚对苯二甲酸乙二醇酯(PET)和聚丙烯(PP)等塑料种类的存在下,土壤容重降低和土壤孔隙度增加,这会导致植物根部更好地渗透到土壤基质中,最终增加了根系生物量(de Souza Machado et al.,2019;陈瑶等,2022)。土壤中的这种变化可能会促进根系渗透并最终刺激根系生长。聚乳酸(PLA)会对土壤特性产生更大的影响,主要体现在PLA 降解产生乳酸导致了土壤pH 值的降低,pH 的降低会进一步干扰叶片中叶绿素合成所必需的营养物质的运输(Qi et al.,2018)。此外,NPs 的疏水表面会与疏水化合物相互作用。在土壤中,许多疏水性和两亲性化合物能调节物种交流和生态系统过程(Wu et al.,2021)。例如,疏水蛋白是土壤中普遍存在的由真菌分泌的两亲蛋白。实验表明,NPs 会抑制氢化酶的自组装,将表面从疏水性改为亲水性。使疏水蛋白不能扩散到环境中,进而影响土壤的稳定性(Huang et al.,2019)。这些富含半胱氨酸的多肽在土壤疏水性和土壤团聚体稳定性中发挥重要作用,对土壤侵蚀和生物地球化学循环具有直接的影响(Pidatala et al.,2016)。
NPs 对土壤中微生物会产生影响,从而影响到植物的生长(Yu et al.,2021a)。NPs 会导致土壤微生物死亡率和组织损伤的增加,以及细菌群落丰富度和多样性的降低,因此土壤微生物活性受到显著影响(van Weert et al.,2019)。PE 和PP 增加了芽单胞菌目的丰度,促进了不可用磷的溶解,从而导致植物有效磷的增加(Qi et al.,2020)。PE 和PVC 刺激了与固氮相关的细菌,直接影响植物对氮的利用(Berruti et al.,2015)。因此,土壤中的氮循环和磷循环在一定程度上受到NPs 的影响(Chen et al.,2015),这可能导致土壤质量的改变和植物生长指标的改变(Fincheira et al.,2018)。NPs 对土壤微生物影响的另一个机制是调节根际丛枝菌根真菌(AMF)的活性。AMF 从植物中获取必需的碳水化合物和其他营养物质,同时促进植物根系对水分和养分的吸收,与植物形成协同关系(Yi et al.,2021)。事实上,PES 显著增加了AMF 菌丝的丰度(王英成等,2022),刺激了葱的生长。NPs 可能也会影响土壤养分和水分的比例。一些NPs 携带丰富的碳(例如PS、PE),这可能间接改变了土壤的碳含量(de Souza Machado et al.,2018)。NPs 引起的碳含量变化会改变微生物群落与植物菌根的共生关系(Fei et al.,2020;胡靓达等,2022),影响菌落碳、氮、磷相关酶的活性,最终影响植物生长(Yi et al.,2021)。尽管与碳相比,NPs 中氮和磷的含量可以忽略不计,但它们也可以改变土壤微生物介导的养分转化。此外,由于塑料污染,在土壤中观察到高C∶N 比,导致营养元素的微生物固定增加(Błońska et al.,2021)。
目前,在实际土壤环境中,NPs 的浓度还尚未可知。尽管有实验研究模拟了高NPs 浓度的土壤环境,但在高浓度的土壤-植物系统中发生的反应可能与在低浓度水平上有所不同(Wu et al.,2021)。此外,目前大多数研究都是在短周期,如几周或几个月的实验期内进行,这些实验结果仅反映了NPs对土壤-植物系统的短期影响。据此,往后的研究需要注重监测实际土壤中NPs 浓度以及对土壤理化性质的影响。而且,针对土壤中NPs 对土壤-植物系统的长期影响研究也亟待开展。在研究NPs 对土壤毒害效应时,没有考虑NPs、植物体、土壤、土壤中的微生物、土壤中的污染物的协同作用,未来的研究需要多方面考虑NPs 与其他环境污染物的联合污染对土壤的潜在影响,开发准确的方法进行土壤样本中的NPs 定量和定性分析,揭示NPs 在土壤中的移动和累积机制,探索NPs 与环境污染物的吸附机理等,为保护土壤环境提供坚实的理论基础和技术支持。
4 研究现状和不足
目前,尽管针对NPs 对动植物的影响机制及对土壤的影响规律有了一定的研究。然而,关于NPs在食物链中如何运输、NPs-植物-环境-污染物交互耦合机制、动植物和土壤样本中的NPs 定量分析方法等研究尚未深入开展。表3 总结了NPs 领域的研究现状,并提出了目前研究的不足。这些内容能为当前的NPs 研究建立全面的知识库,而且也为未来的研究提出了建议。
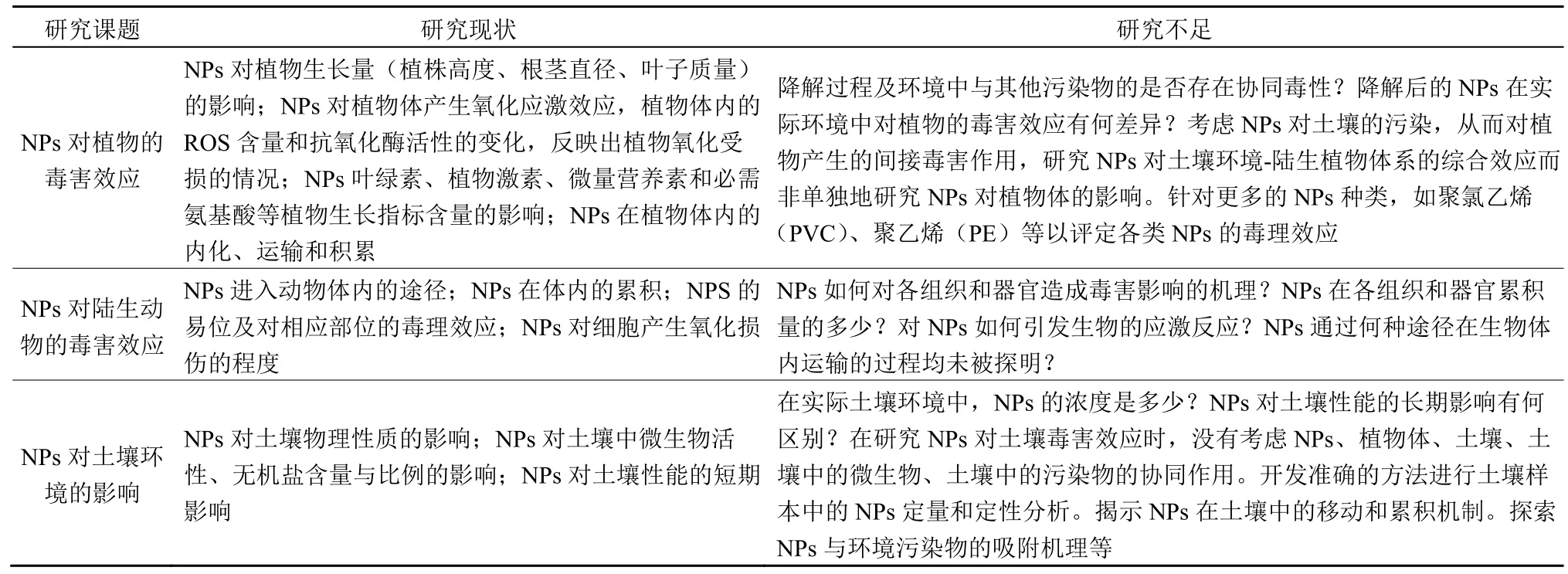
表3 NPs 领域的研究现状和不足Table 3 Current research trends and knowledge gaps of NPs
5 研究展望
NPs 广泛地存在于陆地生态环境中,对陆生动物的各组织和器官都会造成不同程度的损伤。对于植物而言,目前大多数研究都表明NPs 对陆生植物的植株高度、根系生长、叶绿素含量、碳水化合物含量均产生抑制作用。此外,土壤环境中的微生物群落、土壤的结构也会受到NPs 的破坏,从而影响到植物根部对营养物质吸收。本文针对NPs 对陆生生态环境中动植物的影响机制及对土壤的影响规律进行了全面综述,分析了目前NPs 的研究进展。根据综述的结果,本文总结了在相关领域的研究不足和未来研究方向,具体如下:
1)目前针对NPs 的研究主要针对初级NPs,而在环境中经过降解等过程形成的次级NPs 的毒性效应研究较少。但经降解的次级NPs 具有更大的比表面积,更有利于吸附其它污染物和导致NPs 内部化学物质的释放,因此可能具有更高的毒性。此外,由于PS-NPs 实验原材料易于获取、表面功能基团稳定且容易被荧光染料染色,所以目前有关NPs 对植物体影响的研究大多使用PS-NPs 来开展,但其他的塑料,如聚氯乙烯、聚乙烯、聚丙烯等常见塑料仍未进行深入研究。因此,未来的研究需要重点关注经过降解后的次级NPs 毒性效应以及针对更多NPs 种类;
2)目前NPs 对生物体的危害研究主要通过体外模型来检测,但对生物体真实影响还有待进一步探究,所以仍具有一定的局限性。而且NPs 如何导致炎症的机理及其与氧化应激效应机理之间的相关性仍需研究。未来的研究可以通过蛋白质组学、转录组学、代谢组学等生物信息分析手段,探讨NPs 对生物体基因、蛋白质、酶之间的相互作用,通过基因表达分析反映生物体代谢状态,深入探究NPs 对生物体的致病机理,以评估NPs 对生物体造成的危害;
3)目前的研究倾向于单独研究NPs 对土壤环境的理化性质影响、NPs 对土壤中微生物群落的毒害作用、NPs 对环境中无机盐成分和比例的影响,但尚未综合所有因素,从一个整体的角度看待NPs对土壤环境的毒害效应。由于土壤环境及植物生长等过程会受到各种因素的影响,单独研究NPs、微生物、无机盐成分、环境污染物等的影响不能准确解释NPs 对土壤环境及植物生长等过程的毒害效应。因此,往后的研究需着力于阐明环境-NPs-生物-污染物的交互耦合机制。
猜你喜欢
科学导报·学术(2020年19期)2020-07-09 11:21:37
小哥白尼(野生动物)(2019年5期)2019-08-27 00:53:38
Coco薇(2016年8期)2016-10-09 16:58:11
汉语世界(The World of Chinese)(2016年3期)2016-01-10 12:15:40
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:10
少儿科学周刊·少年版(2015年3期)2015-07-07 20:57:25
发明与创新(2015年33期)2015-02-27 10:40:02
声学技术(2014年2期)2014-06-21 06:59:06
养殖与饲料(2014年10期)2014-02-28 22:15:02
养殖与饲料(2014年10期)2014-02-28 22:14:58
