
乙烯利对结缕草响应低温胁迫的作用
2024-01-11 10:40李进超张智韦张嘉航韩翔劲许立新
草业科学 2023年12期
李进超,张智韦,张嘉航,韩翔劲,杜 洋,许立新
(1.北京林业大学草业与草原学院, 北京 100083;2.中煤科工生态环境科技有限公司, 北京 300450)
乙烯利是一种应用广、效果好、安全且残留少的植物生长调节剂,可调控植物种子休眠、果实成熟、花芽分化等生长过程,能够减少植物受到的逆境伤害,如机械损伤、冷害及干旱胁迫等[1-3]。有研究显示乙烯利有助于提高葡萄(Vitis vinifera)的抗寒性[4]。乙烯利还可增加玉米(Zea mays)、甘蔗(Saccharum officinarum)、草地早熟禾(Poa pratensis)等植物的抗旱性[4-8]。而乙烯利对结缕草耐寒性的影响研究较少。日本结缕草(Zoysia japonica),也称结缕草,原产于中国、韩国、日本等地区,具有耐践踏、耐粗放管理、养护成本较低、病虫害少等优良性状,被广泛应用于绿化草坪、运动场草坪及生态修复草坪的建植中[9-10]。结缕草在我国北方生长绿期较短[11],低温是影响其草坪质量的重要因素之一[12]。因此,研究提升结缕草耐寒性的方法,对其进一步推广和应用具有重要意义。有报道显示结缕草和假俭草(Eremochloa ophiuroides)、海滨雀稗(Paspalum vaginatum)、 以 及‘ 天 堂 草419’(Cynodon dactylon‘Tifgreen 419’)等在低温胁迫下叶片中游离脯氨酸、可溶性糖含量和相对电解质渗透率上升,这种变化和其耐寒性有一定相关性[13-14]。
本研究以日本结缕草为研究对象,分析外施乙烯利对日本结缕草低温胁迫下生理指标的影响,确定乙烯利对结缕草耐寒性的作用,并从渗透调节物质的角度初步分析和研究其影响日本结缕草耐寒性的作用机制。
1 材料与方法
1.1 植物材料
日本结缕草草皮,由南京农业大学白马科研基地提供。
1.2 植株培养
将日本结缕草草皮移栽于直径20 cm 的盆内,种植基质为草炭 ∶ 蛭石 = 3 ∶ 1。置于光照培养箱内进行养护,环境设定为32 ℃、17 600 lx 光照14 h、70%相对湿度(relative humidity,RH),以及28 ℃、无光照10 h、70% RH,养护期间每日浇水,修剪高度为13 cm。
1.3 试验处理
日本结缕草盆栽材料分为乙烯利喷施组和蒸馏水对照组,每组设置低温及常温两个处理,每个处理5 个生物学重复。乙烯利喷施组过量喷施浓度为150 mg·L-1的乙烯利溶液,蒸馏水对照组过量喷施蒸馏水。喷施后对两种植物材料进行低温和常温处理。常温处理条件32 ℃、17 600 lx 14 h、70% RH;28℃、无光照 10 h、70% RH。低温处理条件6 ℃、17 600 lx 14 h、70% RH;4 ℃、无光照 14 h、70% RH。处理前(0 d)和处理后(52 d),进行叶绿素、电解质渗透率、相对含水量、可溶性蛋白、可溶性糖、脯氨酸含量的采样测定。
1.4 测定指标及方法
叶绿素和脯氨酸含量测定参照李合生方法[15],相对含水量的测定采用烘干法[16],电解质渗透率测定参照Blum 和Ebercon[17]的方法,可溶性糖的测定参照Hendrix[18]的方法,可溶性蛋白测定采用考马斯蓝G-250 染色法[19],脯氨酸代谢途径相关基因采样根据Zhang 等[20]所示依据脯氨酸含量检测而定,当低温下脯氨酸上升,喷施乙烯利脯氨酸显著(P <0.05)上升时采样。Ilumina 测序数据与文库构建采用Zhang 等[20]RT-PCR 验证过的转录组库。
1.5 数据分析
用Excel 对数据处理和作图。利用SPSS 20 软件对试验数据进行单因素方差分析,以 Duncan (邓肯) 法对数据进行多重比较。
2 结果与分析
2.1 叶绿素含量
在本研究中,常温处理52 d 后,乙烯利喷施组叶绿素含量较蒸馏水和同组0 d 无显著差异(图1)。低温处理52 d 后,蒸馏水组日本结缕草叶绿素含量较0 d 显著下降(P< 0.05),下降幅度为30.14%;低温处理52 d 后,乙烯利喷施组日本结缕草叶绿素含量略微下降,和低温处理前相比无显著差异。
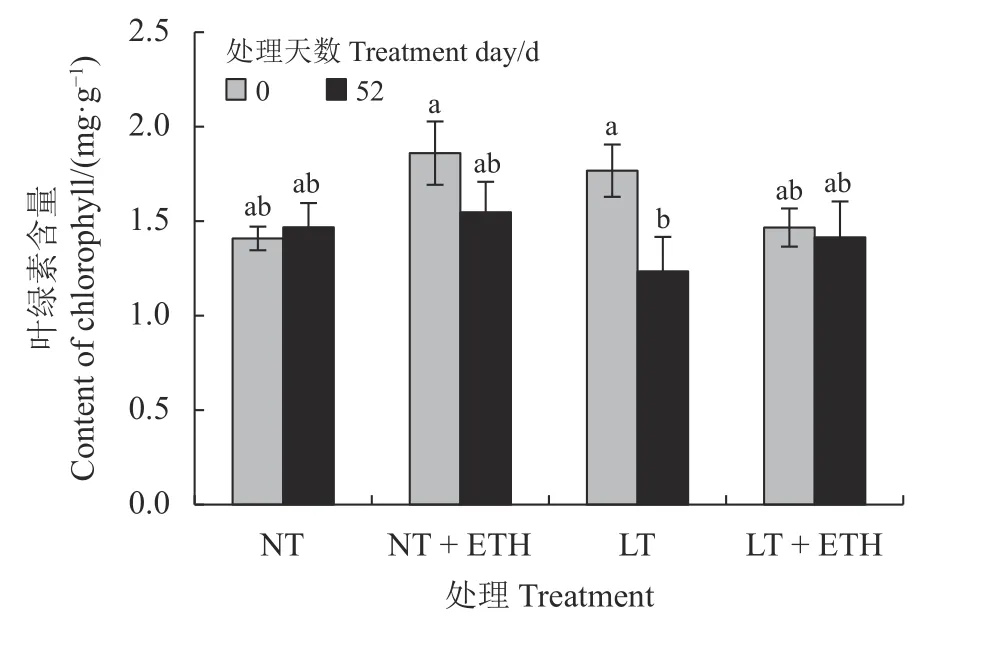
图1 喷施乙烯利对低温胁迫下日本结缕草叶绿素含量的影响Figure 1 Effects of spraying ethephon on Zoysia japonica chlorophyll content under low temperature stress
2.2 相对含水量及电解质渗透率
常温处理52 d 后,乙烯利喷施组日本结缕草相对含水量较蒸馏水和同组0 d 无显著变化(图2)。低温处理52 d 后,蒸馏水组日本结缕草相对含水量较0 d 显著下降(P< 0.05),下降幅度为62.18%;乙烯利喷施组比蒸馏水组相对含水量高。
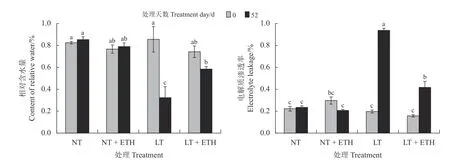
图2 喷施乙烯利对低温胁迫下日本结缕草相对含水量和电解质渗透率的影响Figure 2 Effects of spraying ethephon on Zoysia japonica relative water content and electrical leakage under low temperature stress
在低温处理52 d 后,蒸馏水组日本结缕草电解质渗透率显著上升(P< 0.05) (图2),上升幅度较同组0 d 为376.40%;乙烯利喷施组日本结缕草受低温胁迫影响电解质渗透率也显著上升(P< 0.05),但上升幅度较蒸馏水组低。
2.3 可溶性糖含量
在低温处理52 d 后,蒸馏水组日本结缕草可溶性糖含量较同组0 d 显著下降(P< 0.05) (图3),下降幅度为39.01%;乙烯利喷施组日本结缕草可溶性糖含量未下降,显著高于低温前52 d 处理。
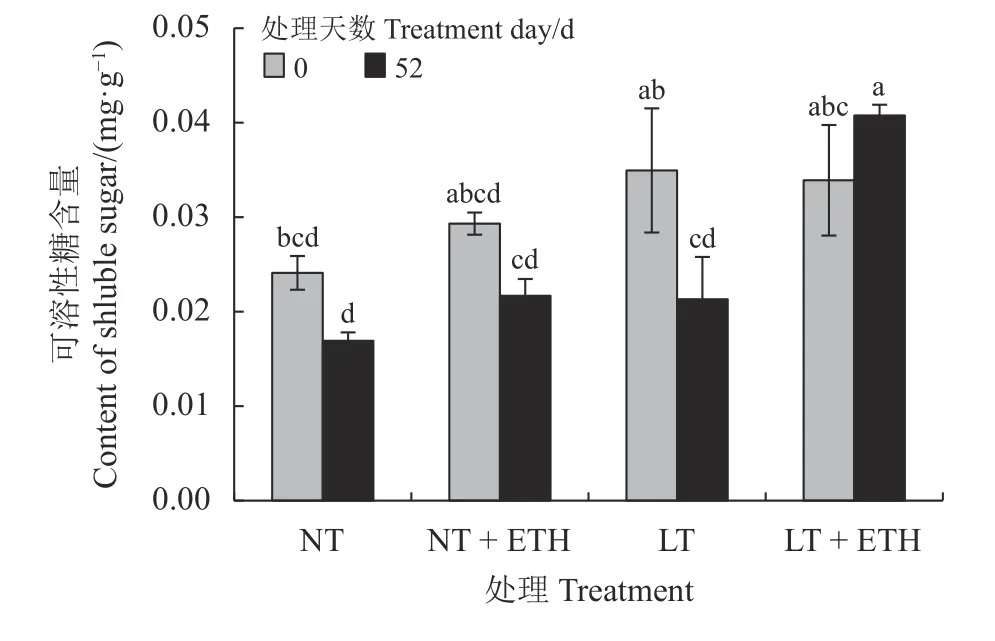
图3 喷施乙烯利对低温胁迫下日本结缕草可溶性糖含量的影响Figure 3 Effects of spraying ethephon on Zoysia japonica soluble sugar content under low temperature stress
2.4 可溶性蛋白含量
常温处理52 d 后,乙烯利喷施组日本结缕草可溶性蛋白含量较同组0 d 显著上升(P< 0.05) (图4),上升幅度为49.75%。在低温处理52 d 后,蒸馏水组日本结缕草可溶性蛋白含量显著下降,下降了31.29%;乙烯利喷施组日本结缕草可溶性蛋白含量略微下降,与蒸馏水组无显著差异。
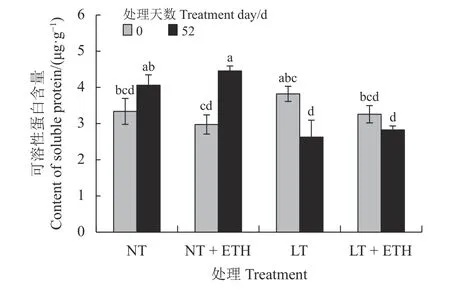
图4 喷施乙烯利对低温胁迫下日本结缕草可溶性蛋白含量的影响Figure 4 Effects of spraying ethephon on Zoysia japonica soluble protein content under low temperature stress
2.5 游离脯氨酸含量
在低温处理52 d 后,蒸馏水组日本结缕草脯氨酸含量较同组0 d 显著上升(P< 0.05) (图5),上升幅度为592.97%;乙烯利喷施组日本结缕草52 d 后脯氨酸含量也显著上升(P< 0.05),是蒸馏水组脯氨酸含量的3 倍多。
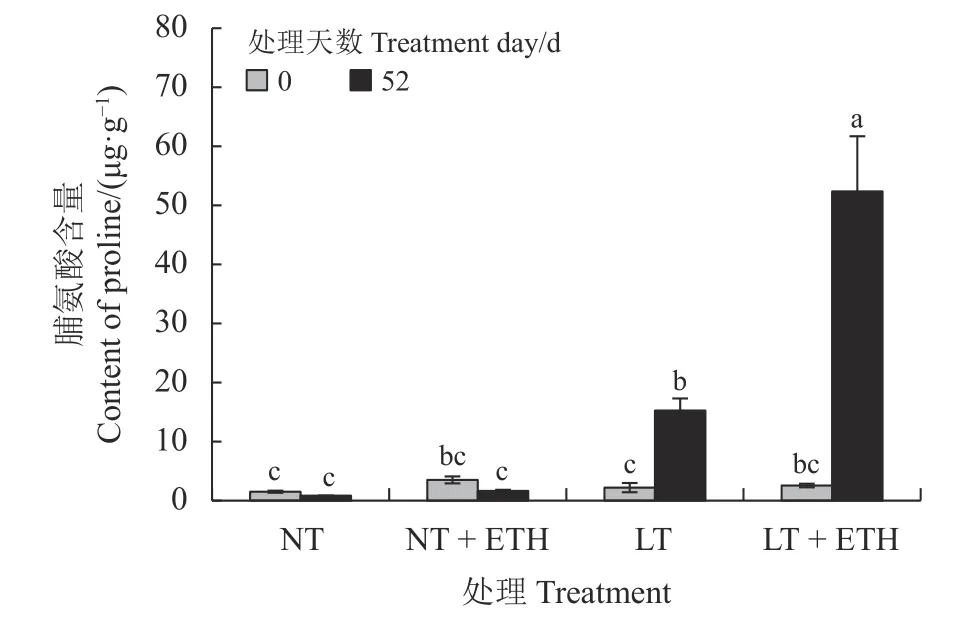
图5 喷施乙烯利对低温胁迫下日本结缕草游离脯氨酸含量的影响Figure 5 Effects of spraying ethephon on Zoysia japonica proline content under low temperature stress
2.6 脯氨酸代谢途径的相关基因变化
在高等植物中,脯氨酸的生物合成存在两条途径(图6 a)。P5CS和P5CR基因参与脯氨酸合成谷氨酸(glutamic acid,Glu) 途径,Glu 首先由Δ1-吡咯琳-5-羧酸合成酶(Δ1-Proline-5-carboxylate s-ynthase,P5CS)作用直接生成γ- 谷氨酸半醛(Glutamic semialdehyde,GSA),GSA 自发转化Δ1-吡咯琳-5-羧酸(Pyrroline-5-carboxylate,P5C),P5C 中间体在Δ1-吡咯琳-5-羧酸还原酶(Δ1-Proline-5-carboxylate reductase,P5CR)催化下产生脯氨酸(Proline,Pro)。OAT和P5CR基因参与脯氨酸合成鸟氨酸(Ornithine,Orn) 途径,鸟氨酸被鸟氨酸-δ-转氨酶(Ornithine-oxo-acid transaminase, OAT)转按氨生成GSA,GSA 自发环化成P5C,然后通过 P5CR 转化为脯氨酸。ProDH和P5CDH基因参与脯氨酸分解途径,Pro 先被脯氨酸脱氢酶(Proline d-ehydrogenase,ProDH)生成 P5C,然后在Δ1 -吡咯琳-5-羧酸脱氢酶(Delta-1-pyrroline-5-carboxylate dehydrogenase,P5CDH)的催化下转化为 Glu。

图6 脯氨酸代谢途径基因间的相互关系模式图及表达量的变化Figure 6 Intergene relationship patterns and changes in proline metabolic pathway expression
蒸馏水组低温条件与常温条件相比日本结缕草P5CR和P5CDH基因无表达(图6 b),OAT和P5CS基因表达上升,ProDH基因表达下降;乙烯利喷施组低温条件与常温条件相比P5CS基因表达上升,OAT基因表达无变化,ProDH基因表达下降。
3 讨论
3.1 乙烯利对叶绿素含量的影响
叶绿素是高等植物吸收、传递光能必不可少的色素分子,叶绿素含量在一定程度上与光合速率显著相关,因此影响植物生长状况,进而间接影响植物的抗逆性[21]。已有研究表明,低温胁迫抑制裸燕麦(Avena nuda)和水稻(Oryza sativa)叶绿素生物合成,导致叶绿体功能紊乱,叶绿素积累减少[22-23]。与此一致,本研究中低温胁迫造成蒸馏水组日本结缕草叶绿素含量大幅减少,喷施乙烯利可缓解低温胁迫下叶绿素的下降幅度,表明乙烯利可能减缓低温诱导的叶绿素降解过程,增加叶绿素积累,Zhang 等[20]对叶绿素合成与降解的基因分析,也印证了这一观点。冯仁军等[24]研究香蕉(Musa paradisiaca)低温胁迫下的表现发现,乙烯利对叶绿素同样的作用,可能原因是乙烯利通过保护植物光合器官增强了有机物积累。而丁久玲等[21]发现低温胁迫下喷施乙烯利对铁皮石斛(Dendrobium officinale)叶绿素含量无显著变化。说明乙烯利对低温胁迫下植物叶绿素的影响不一致,乙烯利浓度以及植物发育阶段和种类等都可能是重要因素。
3.2 乙烯利对相对含水量及电解质渗透率的影响
植物叶片相对含水量是体现植物生长发育状况和抗逆性的重要指标,它能够评估植物吸水能力和体内水分变化,同时反映细胞代谢活性。植物在受到胁迫时细胞膜功能受损,细胞膜透性增大,细胞内物质外渗,电解质渗透率上升。因此电解质渗透率是植物受到胁迫程度的重要指标[19]。本研究表明,低温胁迫下,蒸馏水组日本结缕草相对含水量下降,电解质渗透率上升,这与阳妮[25]等在柑橘(Citrus reticulata)低温胁迫下的研究结果一致。低温下,乙烯利喷施组日本结缕草相对含水量的下降幅度减小,说明喷施乙烯利能够提高日本结缕草在低温胁迫下的保水能力,抗寒性得到提升。喷施乙烯利后抑制了低温胁迫下日本结缕草电解质渗透率的上升,可能是乙烯利能够促进逆境胁迫下抗氧化酶的活性,为细胞膜提供了更好的抗氧化保护[26],从而减轻了膜系统受损程度,提升了细胞膜的完整性,其具体作用机制有待进一步研究确定。
3.3 乙烯利对可溶性糖的影响
可溶性糖是一种重要的渗透调节物质,可通过调节细胞液浓度,提高细胞的吸水和与脂质相互作用能力,增强糖分的积累,从而减少呼吸作用对植物的伤害。大量研究表明,适宜的低温胁迫导致植物可溶性糖含量上升,而过度低温会使植株受到迫害,可溶性糖含量下降。低温胁迫下植物可溶性糖的含量可以反映其抗寒性的强弱,抗寒性强的植株在低温下能迅速提高可溶糖含量,可调节细胞的渗透势,降低细胞冰点温度,避免细胞因温度过低而凝固[27]。本研究中,低温下日本结缕草可溶性糖含量降低,表明植株受到胁迫的时间过长或程度较大,糖分无法积累。外施乙烯利可以通过促进日本结缕草糖分的积累,提高细胞吸水能力,减缓植株受到的胁迫损伤。
3.4 乙烯利对可溶性蛋白的影响
可溶性蛋白通过提高植物的保水能力,保护细胞的生命物质及生物膜,是衡量植物抗寒性的重要指标。可溶性蛋白的积累与植物低温耐受性有关,在一定的低温胁迫下,可溶性蛋白含量会增加,而温度过低或低温持续时间长,使其保水能力下降,可溶性蛋白积累减少[27-28]。本研究中,在常温下喷施乙烯利日本结缕草可溶性蛋白升高,在低温下可溶性蛋白下降,喷施乙烯利使可溶性蛋白维持基本不变,说明乙烯利可能影响了细胞内蛋白质合成、变性及降解等过程,从而促进了可溶性蛋白的积累。
3.5 乙烯利对游离脯氨酸含量的影响
渗透调节物质主要在调节细胞溶液浓度中发挥作用,脯氨酸被认为是一种重要的渗透调节物质之一,可作为稳定蛋白质结构的分子伴侣,细胞氧化还原电位的调节剂以及控制自由基水平的抗氧化剂。脯氨酸主要通过促进渗透压调节,降低植物细胞的水势,使植物一定程度免受胁迫引起的脱水,提高植物对胁迫的耐受能力[29-30]。大量研究表明,在低温胁迫下,植物体内脯氨酸含量上升,植物体内积累的脯氨酸可以提高植物的耐受性。在本研究中,低温胁迫下蒸馏水对照组日本结缕草脯氨酸的升高,在裸燕麦[22]低温胁迫处理中也有同样的研究结果。在低温条件下喷施乙烯利日本结缕草脯氨酸含量升高,且上升幅度高于未喷施乙烯利对照组,说明喷施乙烯利可以促进日本结缕草低温胁迫下脯氨酸的积累,乙烯利可能通过积累脯氨酸增强渗透势调节能力,减少自身受到的胁迫损伤[21]。
3.6 乙烯利对脯氨酸代谢途径相关基因的影响
细胞中的脯氨酸水平取决于脯氨酸合成代谢和分解代谢之间的平衡。谷氨酸和鸟氨酸途径是植物中脯氨酸合成的主要两种途径,P5CS 是脯氨酸合成途径的限速酶,OAT 是鸟氨酸合成脯氨酸途径的主要催化合成酶[31]。脯氨酸分解代谢过程是合成过程的逆过程,ProDH 是脯氨酸脱氢酶,负调控脯氨酸积累,催化脯氨酸脱氢生成延丙酮酸,是脯氨酸降解过程中的限速酶[32]。
日本结缕草低温胁迫下,P5CS基因的表达增强, 而ProDH基因的表达受到抑制,OAT同时高表达(图6b),说明在基因调控水平上,结缕草响应低温胁迫,促进脯氨酸合成的同时抑制其降解。这与其他植物在胁迫条件下,P5CS基因的表达增强,而ProDH基因的表达受到抑制的研究结果一致[32-33]。例如,杨婷等[34]发现盐胁迫可导致小桐(Jatropha curcas)幼苗脯氨酸含量的大幅上升,上调小桐子P5CS和小桐子OAT基因的表达水平增强,同时盐胁迫下小桐子ProDH基因的表达水平下降。张丹等[35]发现中国沙棘(Hippophae rhamnoidessubsp.sinensis)受到干旱胁迫时,P5CS基因表达量上调、ProDH基因表达量下调共同影响中国沙棘叶片中脯氨酸含量的积累。高表达P5CS基因对脯氨酸的积累以及抗逆性的形成有重要的意义,过表达P5CS转基因植物的脯氨酸含量得到大幅提升。例如Zhao 等[36]发现拟南芥(Arabidopsis thaliana)中异源表达互花米草(Spartina alterniflora)SaP5CS2基因,提升了转基因拟南芥脯氨酸的合成,从而提高了抗盐胁迫的能力。朱虹等[29]发现在拟南芥中,拟南芥P5CS1基因响应各类胁迫诱导表达,与植物体内脯氨酸的含量有着显著的关系。汪忠杰[37]发现,茶树(Camellia sinensis)在3 种胁迫处理下,CsP5CS基因表达显著提高,游离脯氨酸含量增加。
乙烯利处理日本结缕草植株在低温胁迫下和对照植株呈现同样的表达模式,P5CS基因上调表达,ProDH基因下调表达,但与对照相比,P5CS基因表达量的上升幅度减小,ProDH基因抑制表达的幅度也减少。这说明,在低温胁迫下,乙烯利的负调控脯氨酸合成。已有研究证明P5CS 酶受脯氨酸反馈抑制调控[31],乙烯利抑制了P5CS基因上调表达幅度,可能正是受到过高含量脯氨酸的负反馈调节作用,侧面证明此时结缕草植株中大量累积了脯氨酸,这个结果也得到印证(图5)。ProDH可能也受到植物体内脯氨酸积累增加而带来的负反馈调控作用。李欣[38]发现,PEG-6000 模拟干旱逆境下,马铃薯(Solanum tuberosum)试管苗的ProDH基因表达量随着脯氨酸含量的增加而下降;且试管苗的脯氨酸含量与ProDH基因表达量极显著负相关。说明乙烯利处理提升了结缕草低温胁迫下的脯氨酸含量,脯氨酸一定程度通过负反馈抑制脯氨酸合成相关基因的表达。
4 结论
喷施乙烯利可以提升结缕草低温胁迫的耐受性,具体表现为可以缓解低温胁迫下结缕草叶绿素含量损失,抑制叶片相对含水量的下降,提高日本结缕草叶片持水能力;减轻细胞膜受损、保持细胞膜的完整性,抑制电解质渗透率的上升。乙烯利处理可能通过调控渗透调节物质,如可溶性蛋白、可溶性糖、脯氨酸,降低细胞液渗透势,提高结缕草的低温胁迫适应性。低温胁迫下,日本结缕草通过基因表达层面“促进合成、抑制降解”的策略,促进叶片脯氨酸的积累。乙烯利处理大幅提升低温胁迫下脯氨酸含量,从而负反馈抑制其合成相关基因表达。
猜你喜欢
中老年保健(2022年3期)2022-11-21
科学与财富(2019年15期)2019-10-21
石油化工建设(2018年2期)2018-07-11
发酵科技通讯(2018年2期)2018-07-06
湖南林业科技(2017年1期)2017-02-06
自动化博览(2014年9期)2014-02-28
无机化学学报(2014年9期)2014-02-28
中国氯碱(2014年12期)2014-02-28
机电信息(2014年35期)2014-02-27
