
遮阴干旱对白及幼苗光合特性的影响
2024-01-11 10:40尹若勇陈蕾如邓鹏飞沈德舜徐小牛
草业科学 2023年12期
尹若勇,陈蕾如,邓鹏飞,沈德舜,徐小牛
(安徽农业大学林学与园林学院, 安徽 合肥 230036)
由于植物在生长过程中会遇到各种非生物胁迫,包括重金属、盐、干旱、营养缺乏、光照强度、农药污染以及极端温度等。这些胁迫通过降低气孔导度、引起氧化应激和降低Rubisco 酶的活性进而阻碍植物进行光合作用。此外,非生物胁迫还会破坏光系统(PS Ⅰ和PS Ⅱ)、阻碍光合电子传递和叶绿素生物合成[1]。光照和水分限制是影响植物光合作用的重要因素[2]。植物的光合生理特性会随着外界环境中水分和光照的变化而发生改变。在干旱胁迫初期,气孔限制是引起植物光合作用下降的主要因素[3]。干旱胁迫通过降低植物向叶片供水的能力而阻碍光合作用[4],此时植物通过提高水分利用效率、降低蒸腾作用等生理调节来应对干旱胁迫[5-6]。随着干旱胁迫程度的增加,光合酶的活性降低,光合磷酸化和ATP 的合成也进一步降低。光照是影响植物生长的另一重要环境因素,植物生长的最适光照强度因物种而异,通常弱光下,植物的光合作用受到抑制,会直接导致光合速率下降,此时植物会通过增加光合色素的方式增大光捕获能力来应对光照不足的影响。在强光环境下植物的生长也会受到抑制,光合色素会在强光下降解导致光合作用受限。
自然条件下,植物往往会受到遮阴和干旱的双重胁迫,有关遮阴和干旱双重胁迫对植物光合特性的影响存在不同观点。“权衡理论”认为,遮阴会加重干旱对植物的影响,植物为了应对遮阴环境下光照不足的影响,会增加植物地上部分的生长量以提高光捕获能力和光能利用率,从而导致植物利用地下水分的能力降低,加重干旱的影响[7]。相反,“促进理论”则认为,遮阴可以减轻干旱对植物的负面影响[2]。“相互作用理论”则提出,过高或过低的光照均会加重干旱对植物的负面影响,中度遮阴下植物受干旱的影响会得到缓解[8]。“独立影响理论”指出干旱与遮阴二者彼此独立,互不影响[9]。
白及(Bletilla striata)是兰科白及属多年生药用兼观赏草本植物,其干燥假鳞茎是我国传统珍贵中药材,具有收敛止血、消肿生肌等功效。我国陕西南部、甘肃东南部、江苏、安徽、贵州等地是白及的主产区[10]。自然条件下,白及分布于常绿阔叶林、栋树林或针叶林下、路边草丛或岩石缝中[11-12]。国外学者对白及的研究主要集中在化学成分和药理作用方面[13],国内学者对遗传多样性[14]、种苗繁育和组培快繁技术[15]、白及属植物资源分布与评价[16]等方面做了研究。白及为耐阴植物,高光强和过度遮阴均不利于白及生长发育[17]。吴明开等[18]研究表明适当遮阴有利于白及生长发育,权雪和张石宝[19]表明水分会显著降低白及的光合能力,不利于其生长发育,王莹博等[20]也发现干旱胁迫会抑制白及光合作用。目前,关于遮阴和干旱交互对白及光合生理影响的研究尚不够深入,缺乏对白及相关胁迫下生长发育的理论指导。因此,本研究选用二年生白及幼苗,通过盆栽试验探讨不同遮阴和干旱胁迫环境下白及光合特性及叶绿素荧光参数的变化。并提出以下假设:适当遮阴有利于白及植株应对干旱环境的变化,但重度干旱时的遮阴会加重对白及叶片光合生理的负面影响。本研究旨在揭示白及对不同干旱和遮阴环境的响应规律和适应机制,为白及高效栽培提供理论依据。
1 材料与方法
1.1 试验材料
2021 年8 月15 日,选择100 株苗高、长势一致的二年生白及幼苗 (平均株高约35 cm)进行缓苗,采用土培盆栽法单株单盆培养,选用园土 ∶ 草炭土 ∶珍珠岩 = 1 ∶ 1 ∶ 1 (质量比)的混合基质,塑料花盆尺寸为11.6 cm × 10.5 cm × 8.8 cm,每盆基质干质量为220 g。在缓苗过程中,利用定期称重计算含水量,确保基质水分含量均一且充足(达田间最大持水量的75%)。昼夜温控变化在26 ℃/24 ℃ (昼/夜),光照时间12 h,空气湿度为60%,光照强度为60%最大光照强度。
1.2 试验设计
试验于2021 年9 月30 日开始,将完成缓苗的白及幼苗植株放入人工气候培养箱(最大光照强度13 000 lux,德国,CLIMACELL)中控制培养,培养箱相对湿度设定为60%,温度设置为26 ℃/24 ℃ (昼/夜),光照时间12 h。采用双因素裂区试验设计,主因素为光照强度,培养箱光照为20% (重度遮阴)、40% (中度遮阴)和60% (对照),副因素为水分控制,采用Hsiao[21]的水分控制梯度,根据本试验基质用量,称重法控制30% (重度干旱,每盆浇水78 g)、50%(中度干旱,每盆浇水130 g)和75% (正常水分处理,每盆浇水195 g)田间最大持水量,以此模拟干旱及遮阴两种胁迫条件。选择60% 相对光照作为对照是基于野外田间观测。每个处理设置6 株重复,控制试验时长为60 d。试验结束后,测量每株白及幼苗的光合参数、叶绿素荧光参数以及光合色素含量。
1.3 测定指标与方法
1.3.1 光合气体交换参数测定
于上午09:00—11:00 选取从上向下数第2、3 片上部完全展开的白及功能叶片中段,采用CIRAS-3便携式光合测定仪(PP-Systems, USA)测定其光合参数。测量时参数设置为光强600 μmol·(m2·s)-1,叶室温度25 ℃、湿度60%,CO2浓度500 μmol·mol-1。测定指标包括净光合速率(A)、胞间CO2浓度(Ci)、蒸腾速率(E)、气孔导度(gs)、水分利用效率(WUE),计算气孔限制值Ls=1-Ci/Ca(Ca为大气CO2浓度)。
1.3.2 叶绿素荧光参数测定
测定光合作用同时,选用光合测定的同株白及叶片的同一部位,采用JUNIOR-PAM 型便携式调制叶绿素荧光仪(德国WALZ 产),参照Lin 等[22]的方法测定叶片叶绿素荧光参数:叶片暗适应30 min后,记录植株的PS Ⅱ最大光化学效率(Fv/Fm)、潜在活性(Fv/Fo)和实际光化学效率(YⅡ)以及光化学淬灭系数(qP)、和非光化学淬灭系数(NPQ)。其中Fv=Fm-Fo。
1.3.3 叶绿素含量测定
采用乙醇提取法[23],从用于测定光合、荧光参数的植株叶片中剪取部分叶片组织,称取约0.1 g 磨碎叶片,加入10 mL 的95%乙醇,于4 ℃下避光浸泡24 h,摇动数次,直至叶片呈白色。将混合物离心静置后,取2 mL 浸提液待用。用分光光度计(Lambda 750S, PerkinElmer)测量待测液在470、649 和665 nm处的吸光度,并根据吸光度计算叶绿素含量。
1.4 可塑性指数
PI=(Pmax-Pmin)/Pmax。式中:Pmax和Pmin为各参数在所有处理中的最大和最小平均值,PI可塑性指数。
1.5 数据处理
采用SPSS 21.0 软件对测定数据进行统计分析,采用LSD 法进行多重比较;利用Origin 2021 软件作图,数据以平均值 ± 标准误表示。
2 结果与分析
2.1 气体交换参数对不同遮阴及干旱的响应
遮阴和干旱处理对白及幼苗的气体交换参数产生不同程度的影响(图1)。根据双因素方差分析结果可知,遮阴和干旱交互作用显著影响Ls、WUE 和Ci(P< 0.05),遮阴极显著影响WUE (P< 0.01),干旱对gs、A 和E 具有极显著影响(P< 0.01)。在60% (对照)和40% (中度遮阴)光强下,随着干旱程度的增加,Ls和WUE 逐渐增加,Ci、A、E 和gs逐渐降低。正常水分处理下,与60% 光照(对照) 相比,Ls在20%光照(重度遮阴)下显著增加40.00% (P< 0.05),gs在重度遮阴下显著降低44.88%。60% 光照(对照)下,与正常水分处理相比,中度干旱下E 和gs显著降低39.08%和41.00%,重度干旱下A、E 和gs显著降低32.80%、51.72% 和54.06%。WUE 在重度遮阴和中度干旱交互下降到最低,A、E 和gs均在重度遮阴和重度干旱处理下降到最低。
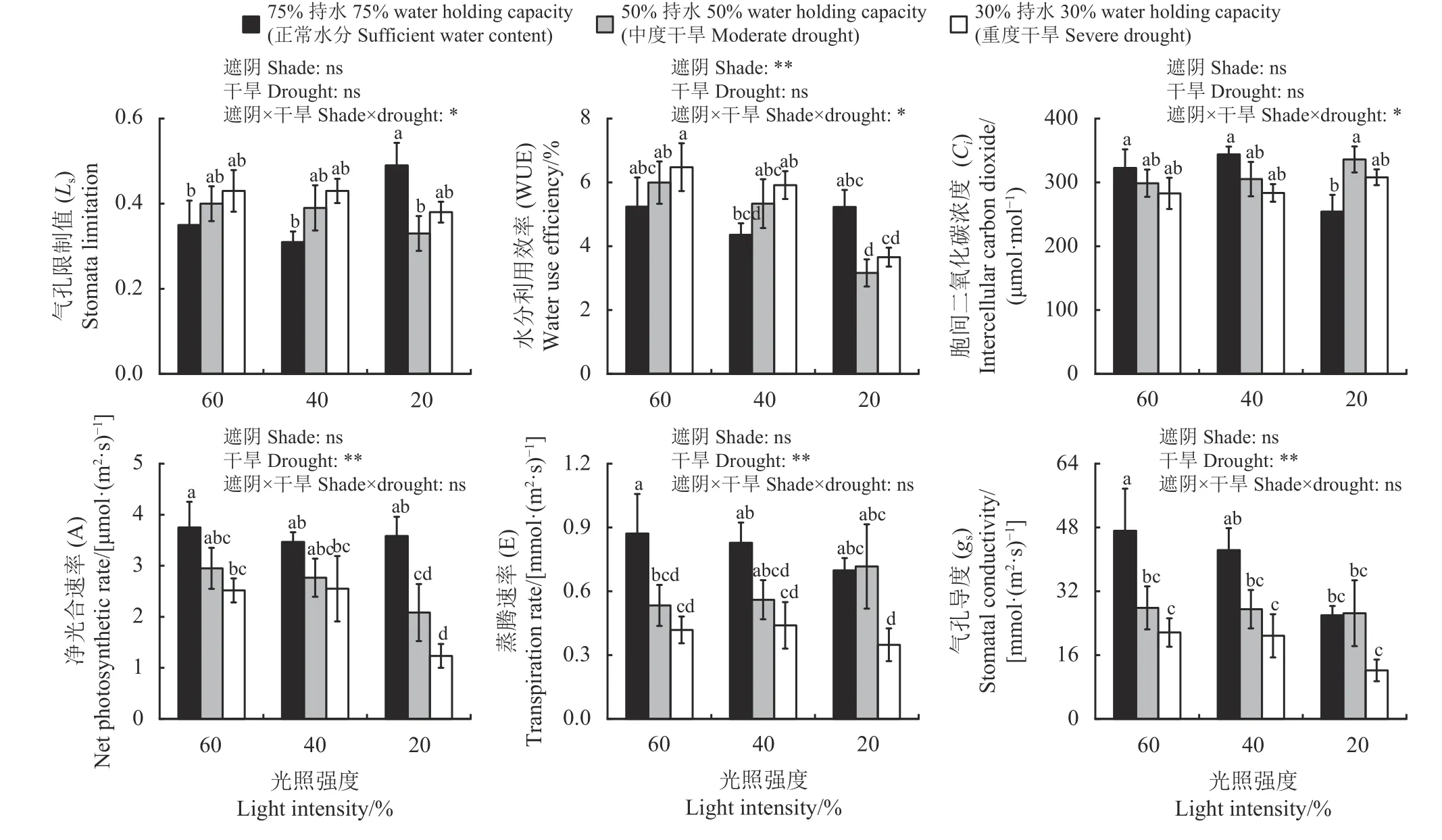
图1 遮阴和干旱处理下白及幼苗光合气体交换参数的差异(n=6)Figure 1 Photosynthetic gas exchange parameters of Bletilla striata seedlings under various shade and drought treatments (n=6)
2.2 遮阴和干旱处理对白及幼苗叶绿素荧光参数的影响
如图2 所示,除qP 外,遮阴对其他叶绿素荧光参数均产生显著影响(P< 0.05)。其中,光照对白及叶片PS Ⅱ的Fv/Fm、Fv/Fo和YⅡ的影响达到极显著水平(P< 0.01)。正常水分下,与60% 光照(对照)相比,Fv/Fo在20%光照(重度遮阴)下降低14.12%。60%光照(对照)下,各荧光参数在不同干旱程度下均未表现出不同。Fv/Fm、Fv/Fo和YⅡ均在重度遮阴和重度干旱交互下降到最低。NPQ 在60%光照和中度干旱交互下最低,qP 在60%光照和重度干旱下最低。

图2 遮阴和干旱处理下白及幼苗叶绿素荧光参数的差异(n=6)Figure 2 Chlorophyll fluorescence parameters of Bletilla striata seedlings under various shade and drought treatments (n=6)
2.3 遮阴和干旱对白及幼苗光合色素的影响
不同程度的遮阴和干旱处理对白及光合色素含量的影响不同(图3)。遮阴对Chla、Chlb、Car、总叶绿素含量和Chla/b 均有极显著影响(P< 0.01);干旱对Chla/b和总叶绿素影响达到极显著水平;所有光合色素含量均受二者交互作用产生极显著影响。正常水分处理下,与60%光照(对照)相比,Chla、Chlb、Car 和总叶绿素在40%(中度)和20%(重度遮阴)光照下均显著增加59.49%和52.38%、59.09%和59.60%、53.16%和66.67%、31.82% 和56.57%,Chla/b 在重度遮阴下显著降低了7.63%。60% 光照(对照) 下,与正常水分相比,Chla、Chlb、Car 和总叶绿素在中度干旱下显著增加36.71%、38.10%、36.36%、39.39% ,Chla、Car 和总叶绿素在重度干旱下显著增加31.65%、31.82%和31.31%。所有的光合色素均在重度遮阴和中度干旱处理下最低。

图3 遮阴和干旱处理下白及幼苗光合色素含量的差异(n=6)Figure 3 Photosynthetic pigment contents of Bletilla striata seedlings under various shade and drought treatments (n=6)
2.4 白及幼苗受遮阴和干旱影响下的可塑性变化
不同遮阴和干旱处理下的白及幼苗可塑性能力基本一致(PI水分总平均= 0.38,PI光照总平均= 0.40),但不同光合生理特征受遮阴和干旱影响的可塑性能力存在差异(图4)。其中,气体交换参数受遮阴和干旱影响的可塑性能力最高(PI水分= 0.59、PI光照= 0.61),受遮阴影响下的可塑性能力比水分高3.4%。其次为光合色素:PI水分= 0.25、PI光照= 0.32;叶绿素荧光参数:PI水分= 0.27、PI光照= 0.24 (图4)。由可塑性指数大小可知,叶绿素荧光参数可塑性指数受干旱影响较高,而光合色素可塑性指数受遮阴影响较高。

图4 遮阴和干旱影响下白及可塑性指数的双变量示图Figure 4 Bivariate plot of plasticity indices of Bletilla striata under varying shade and drought stresses
3 讨论
植物的生理特性会响应外界环境变化,影响植物的生长和发育。本研究中干旱和遮阴均增加了白及幼苗Chla 和Car 含量,这是由于在干旱和遮阴环境下,植物的生长受到限制。白及幼苗通过增加Chla 和Car 含量提高光捕获能力以应对胁迫环境,但重度遮阴和中度干旱交互对白及植株色素含量的消极影响增加;白及植株整体受遮阴和干旱影响的可塑性指数变化也表明对不同遮阴和干旱处理的环境变化适应性较高。与此同时,环境胁迫对植物生长的影响机制因不同的环境因子存在差别,通常认为,在干旱环境中,植物的生长和生产可能会由于光合作用不足而受到限制[24],通过吸收大量水分、减少气孔数量、降低蒸腾作用来适应干旱环境[25]。“气孔限制”[26]和“非气孔限制”[27]是导致植物光合速率下降的主要原因。根据Farquhar 和Sharkey[28]的光合控制理论,植物受到气孔限制的表现主要为gs和Ci降低,同时Ls增大;而“非气孔限制”的影响主要为Ci上升,同时Ls降低。在本研究中,除重度遮阴外,导致白及幼苗A 下降的主要因素均为“气孔限制”,二氧化碳进入叶片受阻,引起白及幼苗进行光合作用时所需的原料减少。植株气孔关闭会导致WUE 增加,同时E 减小[3],这与本研究结果一致。有研究表明,干旱胁迫可使叶绿素的合成受阻或加速叶绿素的降解[29-30],因此会降低植物对光能的吸收和利用能力,引起植株光合功能的改变。而在本研究中,干旱处理对白及幼苗的总叶绿色含量和叶绿素荧光参数的影响表明植株对光能的吸收和利用能力并未因干旱而受到显著影响。
光是主要环境因子之一[31-32],适当的光照有利于植物的生长,当光照过高或过低时,会对植物的生长产生极大的影响。本研究中,正常水分处理时,不同遮阴条件下白及叶片E 随着gs的降低而下降,且Ci也随着gs的降低而降低,但白及叶片A 并未发生显著变化,表明此时白及的光合同化能力仍保持相对协调。此外,植物光合色素具有释放多余能量、防止光损伤、促进光系统修复和缩短光抑制时间的能力,在不同的环境中提高光合作用效率。Chla/b 与环境变化有关,包括光照强度、养分供应、温度和干旱等胁迫[33-35]。若植物的Chla/b较高,则表明其对高光强有更强的适应能力,且增强了光合作用的电子传递效率[36],而Chla/b较低则表明其耐阴性较强[37]。因此,可以通过改变植物光合色素组成从而维持其在不同环境中的光捕获能力来提高植物的光合作用效率[36]。在弱光环境中,植株可通过增加光合色素含量来增大对光能的捕获机会和对光能的吸收和利用能力,以适应弱光环境[38-40]。当植物对光的吸收超过其光合作用能力时,需要耗散多余的能量,否则会对光系统造成破坏,降低光合作用能力。叶绿素是光的收集器,而Car 除了参与光收集外,还具有光保护作用[34]。Car 可以吸收400~550 nm 范围内的光,其存在于所有的光合作用有机体中[41],具有稳定脂膜[40-42]和保护光系统免受活性氧的损害等作用[43-45]。在本研究中,正常水分处理时,白及幼苗在弱光环境下会增加光合色素含量,从而增强对光能的捕获和利用能力;重度遮阴下的Chla/b 低于其他两组遮阴处理,说明Chlb 的增长量相对更高,薛思雷等[46]的研究认为,Chlb 相对含量的增高可提高对弱光的捕获能力。由此可见,白及通过对光合色素含量的调节增强了对弱光环境的适应。Fv/Fm是PS Ⅱ的最大光化学效率,常用于说明植物叶片PS Ⅱ原初光能转换效率,当植物处于正常生长状态时,Fv/Fm值一般为0.75~0.85[47]。而在本研究中,尽管Fv/Fm均随着光照强度的增加而增加,但所有处理下白及的Fv/Fm始终维持在0.7 以上,这说明干旱和遮阴胁迫并未破坏白及叶片的PSⅡ系统,其仍能维持较高的原初光能转换效率,可适应较大的光强幅度。
遮阴和干旱交互作用对白及叶片光合特性和叶绿素荧光参数产生了不同的影响,遮阴提高了干旱下Ci,同时也加重了干旱对WUE 的抑制作用,符合“权衡理论”的观点;而干旱时植株的Ls在遮阴下得到了缓解,Ls降低,符合“促进理论”的观点,这与刘翠菊等[48]对耐冬山茶(Camellia japonica)的研究中得出的结论相同,此时A 下降且在重度遮阴下表现出显著差异,说明此时交互作用可能使植株受到了非气孔限制影响。干旱和遮阴胁迫通过抑制Chla、Chlb 的合成,并加快其分解速度,从而使叶绿素含量降低。本研究中,重度遮阴和中度干旱交互作用对白及光合色素含量产生了负面影响,降低了白及植株对光能的捕获和利用能力,与邓秀秀等[49]对马尾松(Pinus massoniana)的研究结果相同,这也解释了A 在交互作用下呈下降趋势的原因,此时干旱和遮阴互作的影响更符合“相互作用理论”。干旱和遮阴互作对白及植株叶绿素荧光参数并未表现出显著影响,且Fv/Fm均保持在0.7 以上,这与王莹博等[20]对白及的研究结果相似,说明白及植株在本研究条件下具有较强的适应能力,但在重度遮阴和重度干旱交互下白及植株的Fv/Fm和Fv/Fo出现明显下降,表明白及PSⅡ此时受到破坏较为严重,更倾向于“相互作用理论”,若继续施加胁迫可能会造成PSⅡ的破坏,尽管各参数出现波动,但总体较为稳定,若此时解除胁迫,白及的生长将逐渐恢复正常。本研究中不同遮阴处理并未缓解干旱胁迫对白及幼苗的负面影响,这与先前提出的假设不完全相符,可能是由于白及作为阴生植物,对环境光照强度的变化较为敏感,因此对其施加遮阴不会缓解干旱胁迫的负面影响。
4 结论
本研究中,不同干旱和遮阴交互处理下白及幼苗的光合特性呈现出不同的响应和适应机制,重度遮阴和重度干旱交互降低了白及植株的光合能力,影响了白及的正常生长发育,在重度遮阴和中度干旱下光合色素减少,降低了白及的光合作用强度,但叶绿素荧光参数均维持在较为稳定的状态,表明白及未受到不可逆的损伤。因此,在白及实际生产栽培时,需要关注干旱和遮阴对幼苗的影响。
猜你喜欢
科学(2022年4期)2022-10-25
娃娃乐园·综合智能(2022年9期)2022-08-16
中国机械工程(2022年8期)2022-05-09
科学大众(2021年9期)2021-07-16
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
中国调味品(2017年2期)2017-03-20
Coco薇(2016年5期)2016-06-03
现代食品(2016年24期)2016-04-28
考试周刊(2016年6期)2016-03-11
