
噬菌肽SEYT4对食品链中常见致病菌的清除效果
2024-01-01 00:00:00杨甜张月邹更周洋宋智勇吴仁蔚李锦铨
华中农业大学学报 2024年5期


关键词 食源性细菌; 致病菌; 噬菌肽; 食品加工链; 消毒剂
食品生产、加工等过程中,致病菌在不锈钢、塑料等材料制成的食品加工设备表面可长时间存活[1];此外,在家庭厨房中,未清洗的蔬菜和水果由于清洁不当或与砧板等厨具交叉污染也会引起疾病的发生[2-4],导致致病菌对食物及其接触表面的污染成为引起食源性疾病的重要原因[5]。为了避免由致病菌引起的食物中毒,清除在食物接触表面污染的和食物来源的致病菌十分重要。
在食品工业中,常用的化学消毒剂如含氯化合物、乙醇、有机酸和表面活性剂等,虽然能控制致病菌的污染[6],但会对加工设备产生不良影响,甚至危害人体健康[7]。此外,近年来对消毒剂产生抗性的细菌的出现[8],也促使学者们寻求安全有效的新型消毒剂。抗菌肽在控制食品中的致病菌和保持其原有的感官品质和营养特性方面展现出良好的应用前景[9],因此受到了广泛关注。早在1969 年,FAO 和WHO就批准乳酸链球菌肽(nisin)作为食品防腐剂在乳制品、肉制品和罐装食品等领域投入使用[10]。近年来有研究者在噬菌体中也发现了具有抗菌活性的多肽类物质,简称噬菌肽。例如在金黄色葡萄球菌噬菌体中发现的能抑制细菌生长的新型噬菌肽家族[11],以及在假单胞菌噬菌体裂解酶中发现的能够杀死细菌的抗菌肽类似物[12]等,但这些噬菌肽尚未在食品工业中应用。
笔者以课题组前期筛选到的沙门氏菌噬菌体LPSEYT[13]为基础,从中筛选出1 条抗菌肽SEYT4,选择在食品加工链中常见的致病细菌进行杀菌试验,探究该噬菌肽发挥杀菌作用的最佳温度、盐浓度和pH 值并评估其安全性。同时,通过模拟被致病菌污染的叶菜和厨具,探究SEYT4 清除食品中致病菌的效果,以期拓宽抗菌肽的来源,并为今后将该噬菌肽开发为新型食品消毒剂提供理论参考。
1 材料与方法
1.1 材料
1)菌株。猪链球菌(Streptococcus suis)SC19、P1/7、LSM102、LSM178 菌株,粪肠球菌(Enterococ⁃cus faecalis)LEFS2 菌株,屎肠球菌(Enterococcus fae⁃cium)LEFM1、LEFM2 菌株,鲍曼不动杆菌(Acineto⁃bacter baumannii)LAB11、LAB12、LAB13 菌株,铜绿假单胞菌(Pseudomonas aeruginosa)LPA3、LPA4、LPA6 菌株及副溶血弧菌(Vibrio parahaemolyticus)LVP1、LVP78 菌株由笔者所在实验室保存;金黄色葡萄球菌(Staphylococcus aureus)LSA139、LSA140、LSA141、LSA144 菌株,粪肠球菌(Enterococcus fae⁃calis)ATCC51299 菌株,大肠杆菌(Escherichia coli)LEC32、LEC33 菌株及沙门氏菌(Salmonella)ATCC13076、ATCC13311 菌株来源于美国菌种保存中心。
所有菌株使用LB 液体培养基并于37 ℃进行振荡培养。使用添加1.5%( m/V)琼脂的LA固体培养基进行单菌落筛选和细菌计数。
2)30 mmol/L HEPES(pH=7.4)缓冲液。使用1 mol/L HEPES 缓冲液和1 mol/L HEPES sodiumsalt 缓冲液调整pH 至7.4,过滤除菌后用无菌水稀释备用。
1.2 噬菌肽合成及生物信息学分析
噬菌肽SEYT4(RKQNWTEMCNRITDWD⁃MGGKYRGLTIRKQSQSRESQC)采用固相肽技术合成(南京GenScript 公司)并使用高效液相色谱法测定肽的纯度为90.60%。将合成的噬菌肽溶于纯水中,制备质量浓度为1 mg/mL 的原液,-80 ℃保存备用。使用ProtParam(https://web. expasy. org/prot⁃param/)和HeliQuest 服务器(http://heliquest.ipmc.cnrs.fr/)分别预测噬菌肽的各种物理和化学参数。使用I-TASSER 在线服务器(https://zhanglab.ccmb.med.umich.edu/I-TASSER/)预测噬菌肽的结构。
1.3 噬菌肽二级结构测定
分别使用无菌水(水环境)、50% 三氟乙醇(TFE,模拟微生物膜的疏水环境)和30 mmol/LSDS 胶束(模拟带负电的原核膜的类似环境)制备200 μg/mL 的肽溶液以测定在不同条件下肽的状态。将上述肽溶液置于石英比色皿中,使用圆二色谱仪记录其在室温下的波长变化,其中扫描速度为100nm/min,光谱带宽为0.1 nm,带宽为1 nm,扫描波长范围为190~250 nm,每个波长下扫描3 次取平均值,作图后根据扫描出的谱带判断噬菌肽的二级结构。
1.4 噬菌肽杀菌谱测定
测定方法如Euler 等[14]所述,略有修改。挑取供试菌株的单克隆分别接种于5 mL LB 液体培养基中,37 ℃下过夜培养后按比例转接并继续培养至对数期(OD600 nm=0.5, 108 CFU/mL)。离心收集细菌,使用30 mmol/L HEPES 缓冲液重悬并稀释至105 CFU/mL。将上述菌液加入96 孔板,并加入100μg/mL 等体积的肽,阴性对照为加入等体积的HEPES 缓冲液。37 ℃振荡培养1 h 后稀释并涂布LA 固体平板上,过夜培养后计数。试验设置2 组平行,重复3 次,取平均值。平板上细菌菌落数越少即表明噬菌肽的杀菌效果越好。
1.5 噬菌肽杀菌效率测定
细菌处理方法同本文“1.4”,在96 孔板中加入等体积的菌液(105 CFU/mL)和100 μg/mL 噬菌肽,阴性对照为加入等体积的HEPES 缓冲液,置于37 ℃下振荡培养。在预定时间点吸取反应液,稀释至合适梯度后涂布于LA 固体板上,过夜培养后计数。试验设置2 组平行,重复3 次,取平均值。
1.6 噬菌肽杀菌特性测定
细菌处理方法同本文“1.4”,处理后的细菌(105CFU/mL)分别在不同温度、不同浓度盐溶液和不同pH 缓冲液中与100 μg/mL 的噬菌肽反应(除在不同温度下反应的试验外,其余试验反应温度均为37 ℃),振荡培养1 h 后,将反应液稀释后在LA 平板上涂布,过夜培养后计数。阴性对照为加入等体积的HEPES 缓冲液。试验设置2 组平行,重复3 次,取平均值。
1.7 噬菌肽溶血活性测定
取1 mL 5 周龄C57BL/6 雌性小鼠的全血于肝素钠抗凝管中,4 ℃下以3 500 r/min 离心10 min,弃上清后使用PBS 洗涤2~3 次以除去血清和其他杂质,获得100% 红细胞。将红细胞用PBS 稀释至4%(V/V)并吸取100 μL 分装至不同离心管中,加入100 μL 不同质量浓度的噬菌肽,分别以PBS 和1%(V/V)Triton X-100 作为阴性对照和阳性对照,混匀后置于37 ℃孵育1 h,然后将混合物在4 ℃下以3 500r/min 离心10 min,吸取150 μL 上清至96 孔板,用酶标仪测定在540 nm 处的吸光值,试验设置2 组平行,重复3 次,取平均值。试验组与阴性对照吸光值越接近即表明噬菌肽的溶血活性越低,即噬菌肽安全性越高。
1.8 在香菜中的杀菌活性检测
新鲜的香菜(Coriandrum sativum L.)购自华中农业大学农贸市场。使用自来水彻底清洗未受损的香菜叶片,在75% 乙醇中浸泡1 min 后用无菌水彻底冲洗,再在无菌水中浸泡30 min。将大小相近的叶片在HEPES 缓冲液中浸泡10 min 后转移至无菌培养皿中晾干。细菌培养方法同本文“1.4”,吸取10 μL细菌(105 CFU/mL)滴于叶片中央并晾干,将晾干后的叶片转移至加有500 μg/mL 噬菌肽的24 孔板中,37 ℃孵育1 h 后,使用HEPES 缓冲液冲洗叶片后转移至离心管,加入300 μL HEPES 缓冲液并研磨至完全破碎。吸取以上均质液至96 孔板中,稀释至合适梯度涂布于LA 固体平板上,37 ℃下过夜培养后计数;同时将24 孔板中剩余的溶液稀释至合适梯度,在LA 平板上涂布计数。试验设置6 组平行,重复2 次,取平均值。
1.9 在食品接触面上的杀菌活性检测
使用2 cm×2 cm 的无菌塑料板和硅胶片模拟不同材质的厨具。按照本文“1.4”所述处理细菌。随后将100 μL 处理后的细菌(105 CFU/mL)滴加于不同食品接触面上,无菌条件下干燥1 h。接着在滴加细菌的区域滴加等体积的噬菌肽(100 μg/mL),滴加等体积HEPES 缓冲液为阴性对照,并于室温下反应1 h。为了恢复细菌,用无菌棉签彻底擦拭塑料板或硅胶片表面并将棉签浸入含有500 μL HEPES 缓冲液的试管中,将棉签上的细菌全部刮入缓冲液中。悬浮液连续稀释后涂布于LA 固体平板上,37 °C 过夜培养后计数。试验设置6 组平行,重复2 次,取平均值。
1.10 数据分析
使用GraphPad Prism(6.0 版)进行统计分析并作图。检验数据采用双尾t 检验进行分析,Plt;0.05 被认为有统计学意义。
2 结果与分析
2.1 SEYT4 特性分析
根据噬菌体LPSEYT 的基因组(GenBank:MH181876.1),筛选出含有37 个氨基酸的噬菌肽SEYT4。I-TASSER 的预测结果显示SEYT4是1个发夹状的双α-螺旋结构(图1A),分子质量约为4.5ku,净电荷为+4,理论等电点为9.69,疏水指数为0.109,疏水力矩为0.166。SEYT4 是1 个两亲螺旋(图1B),SEYT4 中精氨酸含量最丰富(13.50%),其次是谷氨酰胺(10.80%)。使用圆二色谱法测定噬菌肽SEYT4 在细胞外水环境、疏水膜环境和带负电荷的膜环境中的结构特征,结果如图1C 所示,在水环境中,噬菌肽SEYT4 的结构呈随机卷曲形式,但在疏水膜环境和带负电荷的膜环境中,SEYT4 的圆二色谱在192 nm 附近有1 条正谱带,在208 nm 和222nm 附近分别有1 条负谱带,说明SEYT4 在以上2 种环境中均呈典型的α 螺旋结构。
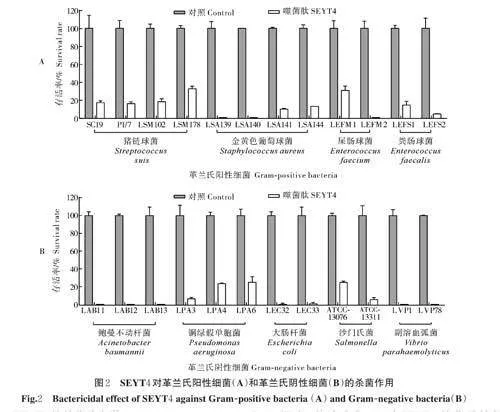
2.2 SEYT4 的杀菌谱
如图2A 所示,SEYT4 对大多数革兰氏阳性细菌均有良好的杀菌活性,对于4 株金黄色葡萄球菌(LSA139、LSA140、LSA141 和LSA144),噬菌肽SEYT4 在1 h 内最多可杀灭约99.99%;对于猪链球菌,噬菌肽SEYT4 在1 h 内最多可杀灭约80.00%;对于粪肠球菌和屎肠球菌,噬菌肽SEYT4 同样有良好的杀菌活性,在1 h 内最多可杀灭约99.99%。
如图2B 所示,SEYT4 能有效杀灭多数革兰氏阴性细菌:噬菌肽SEYT4 能够将鲍曼不动杆菌、大肠杆菌和副溶血弧菌减少约99.99%;可将铜绿假单胞菌在1 h 内最多杀灭约95.00%;对于沙门氏菌,噬菌肽SEYT4 在1 h 内最多可杀灭约95.00%。
2.3 SEYT4 的杀菌动力学
噬菌肽的杀菌动力学结果显示,噬菌肽在2 min内能够将金黄色葡萄球菌减少约99.99%,且在1 h 内经噬菌肽处理后的细菌未见增长,杀菌活性依旧良好(图3A);对于鲍曼不动杆菌,噬菌肽SEYT4 在20min 内能杀灭约99.99%,同样在1 h 内保持良好的杀菌活性(图3B)。
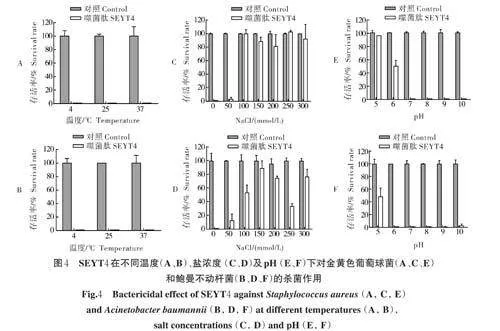
2.4 温度、盐浓度和pH 对SEYT4 杀菌活性的影响
噬菌肽SEYT4 在不同温度(4 ℃、25 ℃和37 ℃)下均具有较高的活性,可将105 CFU/mL 的金黄色葡萄球菌和鲍曼不动杆菌的活菌数量减少约99.99%。而在没有SEYT4 作用时,以上温度下未观察到细菌数量的减少(图4A 和4B)。
在不同浓度的NaCl 溶液中,噬菌肽SEYT4 的杀菌活性受到影响。在50 mmol/L NaCl 溶液中,噬菌肽SEYT4 能够将金黄色葡萄球菌减少约99.99%,能将鲍曼不动杆菌减少约90.00%;但当NaCl 溶液浓度超过50 mmol/L 时,噬菌肽SEYT4 对金黄色葡萄球菌的杀菌活性完全被抑制,对于鲍曼不动杆菌最多可杀灭约60.00%(图4C 和4D)。
在不同pH 下,噬菌肽SEYT4 对金黄色葡萄球菌LSA140(图4E)和鲍曼不动杆菌LAB12(图4F)的杀菌效果不同:在pH=5 和pH=6 时,SEYT4 对金黄色葡萄球菌几乎无杀菌作用,在pH=7~10 时,SEYT4 能将金黄色葡萄球菌减少约99.99%;对于鲍曼不动杆菌,在pH=5 时,SEYT4 几乎无杀菌作用,在pH=7 和pH=8 时,SEYT4 能将细菌减少约99.99%,在pH=6、pH=9 和pH=10 时,SEYT4 可将细菌减少约99.00%。由此可知,该肽在中性和弱碱性条件下杀菌效果更好。
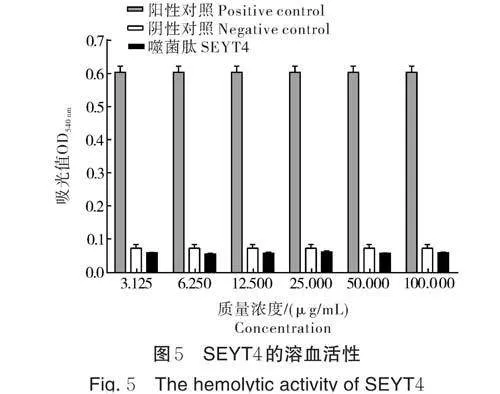
2.5 SEYT4 对红细胞的安全性
SEYT4 的安全性试验结果显示,不同质量浓度的噬菌肽(3.125~100 μg/mL)均不会破坏小鼠红细胞的完整性,表明SEYT4 对小鼠是安全的(图5)。
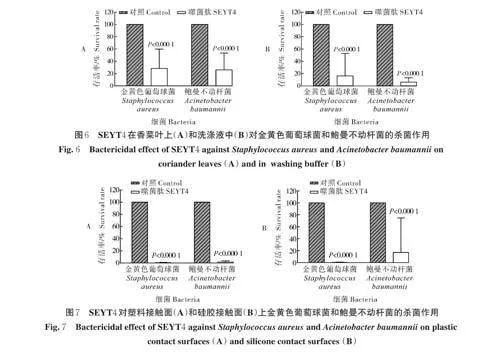
2.6 SEYT4 在香菜上的杀菌作用
在37 ℃ 条件下,用质量浓度为500 μg/mL 的SEYT4 处理香菜叶片1 h 后,观察到受污染香菜叶片上的金黄色葡萄球菌和鲍曼不动杆菌的数量减少约80.00%(图6A),而洗涤液中细菌的数量则可减少约85.00%~99.00%(图6B)。由此可见,SEYT4 能够有效清除叶菜中污染的致病菌。
2.7 SEYT4 在食品接触面上的杀菌作用
在模拟的塑料接触面上,噬菌肽SEYT4 最多可清除约99.00% 以上的金黄色葡萄球菌和鲍曼不动杆菌(图7A),同样,SEYT4 对硅胶接触面上的细菌也有显著的杀菌效果,最多可将细菌数量减少约99.99%(图7B)。表明SEYT4 可作为一种理想的消毒剂,用于清除塑料和硅胶等食品接触表面上的金黄色葡萄球菌和鲍曼不动杆菌。
3 讨论
噬菌肽SEYT4 是笔者所在研究室从沙门氏菌噬菌体LPSEYT 中筛选出1 条活性良好的抗菌肽[13],其杀菌能力及安全性尚不清楚。本研究结果显示噬菌肽SEYT4 可高效清除对数生长期的细菌,杀菌能力在90.00%~99.99%,其中包括许多食源性病原菌和腐败细菌,表明该肽杀菌的广谱性。在肽的杀菌动力学试验中,SEYT4 能够快速清除金黄色葡萄球菌和鲍曼不动杆菌,表明该肽杀菌的高效性。SEYT4 在不同温度下仍保持近99.99% 的杀菌活性,室温下可以有效防止食品加工过程中金黄色葡萄球菌和鲍曼不动杆菌的交叉污染;在较高的pH下,SEYT4 活性较强,但在弱酸性环境中其活性被抑制;当氯化钠浓度大于50 mmol/L 时,SEYT4 会失活,这在一定程度上限制了SEYT4 在偏酸性环境和高盐环境中的应用。有研究表明可从替换氨基酸入手对肽进行改造,改造后的肽在盐存在的情况下对致病菌有强大的杀菌作用[15-16]。因此后续可通过对噬菌肽进行改造以提高其耐受性。随着分子设计和优化策略的发展,以及纳米技术的迅速崛起,有更多方法可以用来改善抗菌肽的生物和化学性质,以拓宽噬菌肽在食品加工链中的应用[17-18]。
噬菌肽在实际生产应用中使用需考虑其安全性,而通过噬菌肽与真核细胞相互作用可以评判噬菌肽应用于食品中的潜力。本研究中安全性试验结果表明,噬菌肽SEYT4 不会损害哺乳动物小鼠红细胞。另外,细胞毒性也是评判多肽类抗菌药物安全性的一个重要指标。已有研究表明抗菌肽CPF-C1质量浓度达到128 μg/mL 时,肽对人胚肾细胞系未显示出细胞毒性[19]。后续可探究噬菌肽SEYT4 的细胞毒性以进一步评估其安全性。
金黄色葡萄球菌、不动杆菌和大肠杆菌等常见的致病菌多存在于食品加工各个环节,在食品加工和储存期间的不良卫生状况可能会导致它们进入食物链。在本研究的香菜叶片模型中,SEYT4 被证明是一种有效的消毒剂,能够明显清除叶片上细菌的同时也能将洗涤液中约99.00% 的细菌清除,大大降低了洗涤过程中交叉污染的风险;在接触面模型中,SEYT4 能清除食品接触面上约99.99% 的细菌,表明噬菌肽在食品加工链中具有很大的应用潜力。然而与杀菌试验相比,噬菌肽在香菜中的杀菌能力相对降低,推测可能是由于噬菌肽与叶片表面未充分接触或香菜中维生素和醇类等物质的存在导致噬菌肽在香菜叶片上的杀菌能力有所降低。
综上所述,噬菌肽SEYT4 可有效控制食品加工链中致病菌的交叉污染,有望成为一种食品加工链中的实用消毒剂。本研究在拓宽抗菌肽来源的同时,也为食品安全领域提供了一种新的防控技术。
猜你喜欢
当代水产(2021年10期)2022-01-12 06:20:40
恋爱婚姻家庭·养生版(2020年11期)2020-12-17 03:26:48
大众科学(2020年3期)2020-06-09 12:27:54
上海农业学报(2017年4期)2017-04-10 12:40:40
现代食品(2016年24期)2016-04-28 08:11:58
中华老年口腔医学杂志(2016年5期)2016-03-01 02:24:24
湖南农业科学(2015年5期)2015-02-27 14:33:56
中国动物保健(2014年1期)2014-04-29 09:05:48
中国质量与标准导报(2014年4期)2014-03-11 19:54:35
哈尔滨医药(2014年6期)2014-02-27 13:35:50
