
基于山地蜂蜜花粉组成的中华蜜蜂传粉多样性研究
2023-12-26 02:44郭新军孟长军
生态学报 2023年23期
郭新军,孟长军,封 婧
西安文理学院生物与环境工程学院,西安市秦岭天然产物开发与抗癌类创新药物研究重点实验室,西安 710065
中华蜜蜂(Apiscerana)善于应用零星蜜源,能较好地适应山区环境[1—2],是我国分布最广的本土蜂种之一。近年来,随着栖息地丧失、气候变化等[3—5],多地野生中华蜜蜂资源已明显下降,在一些地区甚至已经完全消失[6—8]。作为重要的传粉昆虫之一,中华蜜蜂独特的形态结构和生物学特性在植物传粉方面具有很大的优势[9—10],对维持生物多样性和维护生态系统的稳定等起到重要的作用[11—13]。中华蜜蜂种群数量减少意味着植物的授粉率下降、繁殖效率降低、遗传多样性降低等,进而影响生态系统稳定。因此,有必要进一步提升对中华蜜蜂传粉多样性的认知水平,增强对其传粉功能与生态服务价值重要性的认识。
利用孢粉学方法可通过对花粉形态的鉴定、分类来确定蜜蜂传粉植物的组成特点[14],但需要专业的花粉分类知识与技巧。新兴的DNA条形码和高通量测序等技术可用来快速而准确地鉴定花粉混合物的物种组成[15—18],在一定程度上能克服以往研究中植物分类知识及花粉鉴定基础的欠缺对研究者造成的瓶颈。利用该类技术,研究者对不同地区蓝彩带蜂、意大利蜜蜂、白斑切叶蜂等传粉蜜蜂访问的蜜粉源植物进行了分析[18—20],为传粉昆虫访花植物种类的鉴定提供了新的思路。
为进一步了解中华蜜蜂访问蜜粉源植物的组成及特点,增强人们对中华蜜蜂的传粉服务功能重要性的认识,本研究利用聚合酶链式反应(PCR)、DNA条形码和高通量测序等技术,通过分析蜂巢蜜花粉的来源,明确研究区域蜜粉源植物的组成及其特点,为了解中华蜜蜂对蜜粉源植物选择的偏好性及中华蜜蜂与蜜粉源植物间的进化关系提供理论基础,为中华蜜蜂资源的保护提供一定依据。
1 材料与方法
1.1 样品采集
样地分别选择在陕西省宝鸡市(BJ)、贵州省望谟县(WM)、广西壮族自治区凤山县(FS),于2021年9月—11月在每地随机选择3个蜂箱(相距50 m以内,各箱内蜂蜜积累时间均达1年以上)各收集1份蜂巢蜜样品作为1个重复,每地共3个重复。蜂蜜样本来源地点信息如表1所示。

表1 蜂巢蜜来源地信息
1.2 实验方法
1.2.1样品花粉分离、DNA提取、PCR及高通量测序
称取蜂巢蜜样品各50 g,分别置于烧杯中,各加入100 mL蒸馏水,充分搅拌得到样液。分别取10 mL样液置于离心管中,3000 r/min离心10 min,去除上清液后分别加入蒸馏水至10 mL,再进行第二次离心。去除上清液后保留底部残留物,镜检后作为花粉样本。
利用E.Z.N.ATMMag-Bind Soil DNA Kit试剂盒提取花粉样本DNA,采用Qubit试剂盒定量检测DNA浓度以确定PCR反应中其用量。PCR扩增rbcL基因目的片段的引物为rbcL-F(CTTACCAGYCTTGATCG TTACAAAGG)和rbcL-R(GTAAAATCAAGTCCACCRCG)[21—22]。采用2×Hieff® Robust PCR Master Mix PCR预混液进行两轮PCR反应,并在第二轮扩增中引入Illumina桥式PCR兼容引物,构建文库。
文库大小通过琼脂糖凝胶电泳检测,扩增产物委托生工生物工程(上海)股份有限公司进行高通量测序。
1.2.2数据处理
利用Usearch软件将获得的序列进行聚类[23],得到具有代表性的可操作分类单元(OTU)序列。计算各样本OTU水平上蜜粉源植物Alpha多样性数,并利用SparCC计算样本间的相关系数和P值,使用R语言的gplots包绘制样本相关性热图。通过核酸序列比对工具(BLASTN)与基因库(GenBank)中的数据进行比较(https://blast.ncbi.nlm.nih.gov/Blast.cgi),筛选最佳比对结果并获得相应的植物名称,植物分类系统参考中国植物物种信息(http://db.kib.ac.cn/)和中国科学院植物科学数据中心(https://www.plantplus.cn/cn)等。对蜜粉源植物的分布及生物地理特点、花序类型信息进行统计、分析。各蜜粉源植物属的区系特征参考吴征镒等[24—25]。
1.2.3各地样本蜜粉源植物优势种分析
根据样本中各OTU的序列数,计算各种蜜粉源植物在样本中的相对丰度,统计样本中的优势物种,比较样本间物种丰度的差异。
1.2.4蜜粉源植物系统进化关系分析
使用MEGA11对获得的各种蜜粉源植物rbcL基因部分序列进行比对,并采用邻接法(NJ)构建系统树[26—27],对蜜粉源植物的系统发育关系进行分析,并与被子植物系统发育研究组(APG)建立的被子植物分类系统第四版(APG Ⅳ)进行比较[28]。
2 结果与分析
2.1 蜜粉源植物Alpha多样性与地理相关性
各样本文库的覆盖率为1.00±0.00,说明样品中序列未被检测出的概率很低,反映了本次测序结果可代表样品的真实情况。各样本序列数均在40000条以上,包含的蜜粉源植物物种丰富度较高。Chao和Ace指数反映了各样本中的物种丰富度,Shannon和Simpson指数反映了各样本物种多样性,Shannoneven指数反映了各样本物种均匀度。各样本间OTU水平上蜜粉源植物Alpha多样性指数无显著差异(表2)。

表2 样本蜜粉源植物组成Alpha多样性指数
各地样本的不同组样品间存在较高的相关性,而不同地区样本间相关性较低,但WM和FS个别样品具有相对较高的相关性(图1),可能与两地距离相对较近有关。样本间的相关性反映出结果具有一定重复性,且不同地域样本具有明显区别。
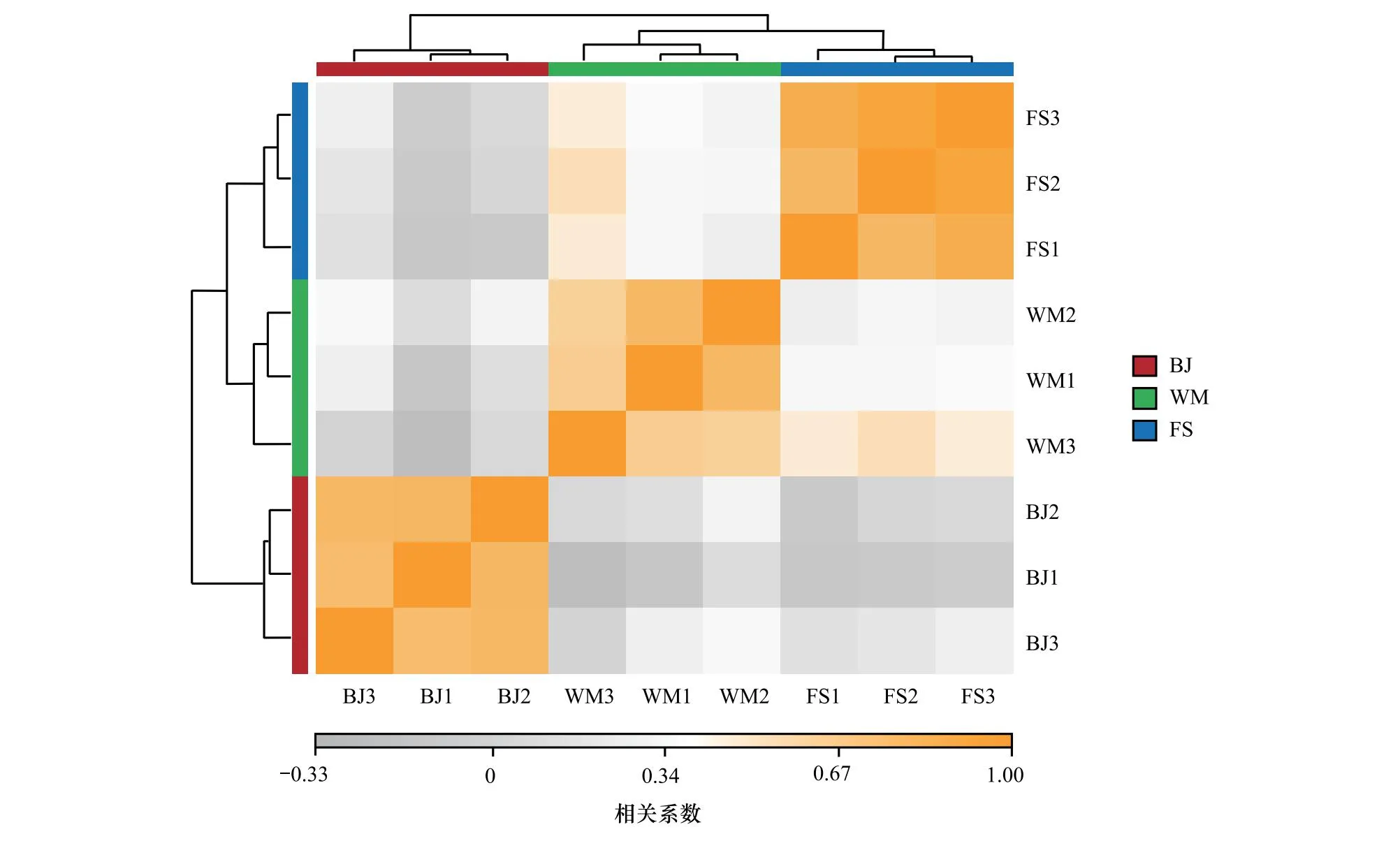
图1 样本相关性热图
2.2 各地样品中蜜粉源植物物种组成
通过对3地9个样品测序获得的序列进行聚类,共获得241个OTU,基于各OTU序列比对分析,共获得蜜粉源植物82种(其中19种只鉴定到属)(表3),隶属于27目46科71属。各样本中,鉴定出最多蜜粉源植物的是陕西省宝鸡市样本(BJ),有57种;贵州省望谟县样本(WM)中有30种,广西壮族自治区凤山县样本(FS)中有40种。BJ和WM共有种20个,BJ和FS共有种24个,WM和FS共有种19个,其中三地共有种18个,占22.0%,主要有红足蒿(Artemisiarubripes)、芜青(Brassicarapa)、翅柃(Euryaalata)等。仅在一个地区的样本中出现的共有55种,占67.1%,其中有31种出现在BJ样本中,有9种出现在WM样本中,有15种出现在FS样本中。由此可见,三地样品中蜜粉源植物的物种组成存在较大差异。

表3 鉴定出的中华蜜蜂访问蜜粉源植物及其分布
鉴定出的蜜粉源植物中,菊目最多,有12种,毛茛目有7种,蔷薇目、禾本目、唇形目各有6种。草本植物和木本植物(含乔木、灌木和木质藤本等)分别为44种和38种。这些植物有少数是花单生(如蛇莓Duchesneaindica),多数形成花序,花序类型包括伞形花序(如蒜Alliumsativum)、头状花序(如鬼针草Bidenspilosa)、总状花序(如芜青Brassicarapa)、穗状花序(如野草香Elsholtziacyprianii)、圆锥花序(如漆树Toxicodendronvernicifluum)等多种类型,但研究结果不能反映中华蜜蜂对花序选择的偏好性。
结果显示,中华蜜蜂采集的蜜粉源植物属的区系复杂(表3,表4),其中包含世界分布区类型(类型1)属共12个,如铁线莲属(Clematis)、鬼针草属(Bidens)等;热带分布区类型(包括类型2—7)属共26个,其中最多的是泛热带分布区类型(类型2)属,共9个;温带分布区类型(包括类型8—14)属共33个,其中最多的是北温带分布区类型(类型8)属。样本中未见分布区类型有温带亚洲分布区类型(类型11)、中亚分布区类型(类型13)和中国特有分布区类型(类型15)三种。根据属的区系成分分析,温带分布区类型的植物属最多,其次为热带分布区类型,所占比例分别为46.5%和36.6%,其中BJ和WM样本中属的分布区类型均是温带分布区类型多于热带分布区类型,而FS样本中热带分布区类型多于温带分布区类型。
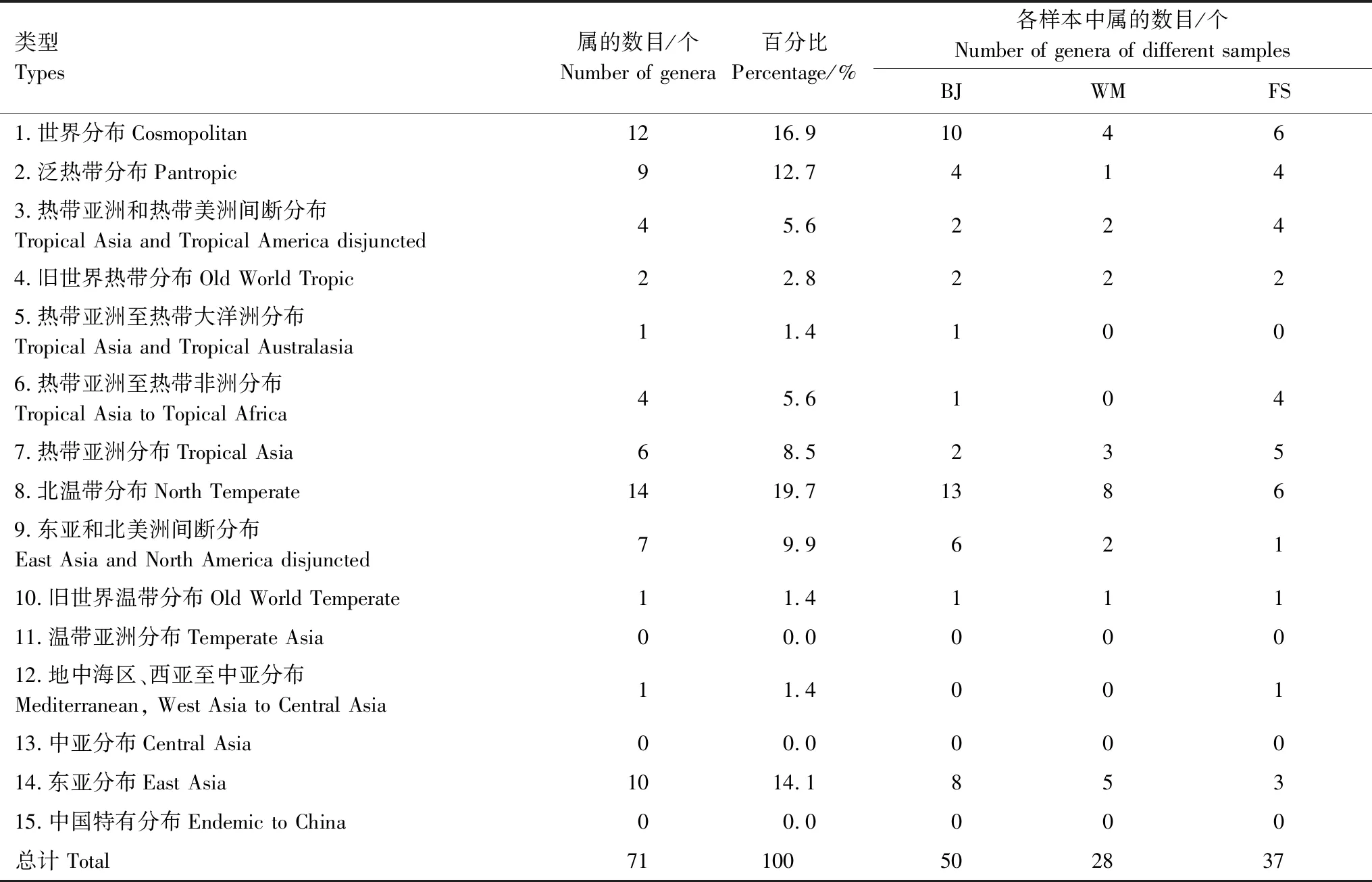
表4 鉴定出的蜜粉源植物属的分布区类型
2.3 各地样本中蜜粉源植物优势种分析
由图2可知,不同样本中优势物种组成情况及其所占比例不同,其中BJ样本中有红足蒿(Artemisiarubripes)、向日葵(Helianthusannuus)等,WM样本中有芜青(Brassicarapa)、粗糠柴(Mallotusphilippensis)等,FS样本中有芜青(Brassicarapa)、枇杷(Eriobotryajaponica)等。

图2 各地样本中蜜粉源植物优势物种及其相对丰度
2.4 蜜粉源植物系统进化关系
中华蜜蜂访问的蜜粉源植物处于APG IV系统的不同分支,包括基部支群、木兰类分支、单子叶植物、真双子叶植物等(图3),说明中华蜜蜂对蜜粉源植物的选择具有一定广泛性,其中在核心双子叶植物菊类分支和蔷薇类分支中某些位置比较集中。
3 讨论
蜜蜂是重要的传粉昆虫类群之一,对其蜜粉源植物多样性的研究可为其资源的保护提供一定依据[18]。中华蜜蜂适应于山地生态环境[29],善于利用零星蜜源[9—10],传统中华蜜蜂养殖取蜜时间往往间隔一年以上,其蜂蜜中包含不同花期植物来源的花粉。本研究在三地样本中,分别鉴定出蜜粉源植物57种(陕西省宝鸡市)、30种(贵州省望谟县)和40种(广西壮族自治区凤山县)。这些蜜粉源植物中,除有刺槐、龙眼、白刺花、向日葵、枇杷、柃等主要蜜粉源植物以及漆树、葎草、紫穗槐、臭椿等重要辅助蜜粉源植物[30]外,还有一些鲜见文献报道的物种。这些山区中蜜粉源植物非常丰富,中华蜜蜂在各地实际访问的植物种类数远大于此。若集约化单一性作物的季节性种植是传粉昆虫种类及数目骤减的主要原因之一[31],那么山区里丰富的蜜粉源植物多样性就是中华蜜蜂等传粉昆虫种群得以存续的重要保障;同样地,中华蜜蜂等传粉昆虫也为山区蜜粉源植物多样性提供了保障。栖息地的减少对中华蜜蜂种群的生存产生重大的影响,目前中华蜜蜂的分布区已经有一定缩减[6—8],而相对来说,山区由于环境受到人类干扰较小,成为中华蜜蜂生存的重要阵地。另外,我国近些年封山育林等工作将对中华蜜蜂资源的保护起到一定积极作用。
本研究选择了不同省份的三个山区,从全球气候分布来看,位于陕西秦岭地区的宝鸡市属于温带季风气候区,位于云贵高原边缘地带的望谟县和凤山县属于亚热带季风气候区。植被特点与气候条件相关,从植物区系划分来看,三个研究区域均属于泛北极植物区,其中宝鸡市在中国-日本森林植物亚区华北植物地区和华中植物地区交界处,植被以北温带分布为主[32—34],鉴定出的蜜粉源植物中温带分布区类型属最多;望谟县和凤山县在中国-日本森林植物亚区滇、黔、桂地区与中国-喜马拉雅森林植物亚区云南高原地区的交界处[32],植被具有温带分布向热带分布过渡的特点,在凤山样本中鉴定出的蜜粉源植物出现了热带分布区类型属多于温带分布区类型属的特点,与文献中地理位置上比较接近的贵州茂兰蜜粉源植物区系特点基本一致[35]。受样本量限制,相关数据不能全面反映蜜粉源植物构成的复杂性,仅为定性分析中华蜜蜂对蜜粉源植物的选择及了解局部地区中华蜜蜂传粉多样性提供一定参考。
中华蜜蜂是我国的本土蜂种,其在长期的进化过程中与蜜粉源植物形成了复杂的进化关系。中华蜜蜂传粉植物种类丰富,其对蜜源植物采集喜好性差异小[9]。有报道称蜜粉源植物的花部性状等可能会影响蜜蜂的选择[36—38],但根据鉴定出的蜜粉源植物花序特点,并未发现相关性状对中华蜜蜂选择的影响。研究发现中华蜜蜂选择的蜜粉源植物中一些占优势的种类主要集中在进化树的菊类分支和蔷薇类分支中,但其是否对进化地位在这些植物附近的类群有所偏好,有待于进一步研究。另外,通过对各地区样本鉴定出的20种主要优势蜜粉源植物分布区域(https://www.gbif.org/)分析发现,有12种与中华蜜蜂主要分布区域呈现出一定的一致性,这将对研究中华蜜蜂起源与其与蜜粉源植物协同进化提供一定依据。
研究中采用的DNA条形码、高通量测序等现代技术为研究花粉组成的多样性提供了方便[16—18],测序结果可为相关研究提供物种多样性、系统进化关系等信息,与孢粉学方法相比,具有分析快速、不完全依靠专业分类知识等优点[14]。但在实际分析中仍存在一些问题,如有部分序列只能鉴定到属,不能准确判断其物种来源,其原因可能是数据库资料不全或数据库中的数据所用分类系统混乱;也可能是由于高通量测序基因片段太短,所得到的序列不足以进行近缘种的区分。随着测序技术的发展及数据库的完善,相关研究方法必将进一步为研究相关问题提供便利,但在实际应用中,种的确定、开花特点分析等还应结合传统分类学知识和实地调查等技术。
综上所述,中华蜜蜂选择蜜粉源植物多样性丰富,相关研究有助于理解中华蜜蜂与蜜粉源植物的进化关系及其在某些植物传粉过程中的重要性,为分析中华蜜蜂对蜜粉源植物的选择机制提供基础,为中华蜜蜂资源的保护提供依据。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
安徽农学通报(2021年22期)2021-12-11
安徽农业科学(2021年14期)2021-09-03
中国蜂业(2021年5期)2021-05-22
中国蜂业(2021年10期)2021-03-02
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
炎黄地理(2019年5期)2019-10-20
中国蜂业(2017年8期)2017-09-05
安徽农业科学(2016年9期)2016-10-14
