
基于耳石地标点法的南黄海和东海小黄鱼种群关系研究
2023-12-25 00:55宋大德康中杰张琥顺李国东吴晓睿李冬佳王淑艳
南方水产科学 2023年6期
梁 龙,宋大德,康中杰,张琥顺,李国东,吴晓睿,李冬佳,王淑艳,闫 欣,祝 斐,熊 瑛
1.上海海洋大学 海洋科学学院,上海 201306
2.江苏省海洋水产研究所,江苏 南通 226007
3.华东师范大学河口海岸国家重点实验室,上海 200241
小黄鱼 (Larimichthyspolyactis) 属鲈形目、石首鱼科、黄鱼属,广泛分布于我国东海、黄海和渤海海域[1]。小黄鱼曾为我国“四大海产”之一,近十年其全球年产量稳定保持在30 万吨以上[2],是中国、朝鲜和韩国近海海域的重要经济鱼类之一[1,3]。小黄鱼是典型的海洋洄游型鱼类[4],以南黄海为例:每年3—5 月,小黄鱼迁徙至苏北近海(水深10~20 m) 海域产卵;6—9 月,向外洄游至机轮拖网禁渔区线附近 (水深30 m 左右) 海域索饵;10—11 月,开始逐步游向外海越冬;12 月至翌年2 月在济州岛西南、东海中南部海域越冬[5-6],栖息海域温度是驱使其洄游的重要因素[7-8]。正因小黄鱼多样的生境履历且不同种群的洄游路线不尽相同[9],其总体上呈现出复杂的族群结构,不同的地理族群之间具有相对独立的种群动态,但彼此之间也存在个体的迁入与迁出[10]。迄今,针对小黄鱼地理族群划分特别是黄海南部海域的小黄鱼群体划分尚未有统一的定论。林新濯等[11]通过对比不同海域小黄鱼的生物学特征指标,认为中国近海海域的小黄鱼可以分为3 个地理种群,分别是黄-渤海、南黄海和东海群;在21 世纪初之前,这一结论广受学界认可[12-13],但之后,徐兆礼和陈佳杰[9]剖析了1971—1982 年我国十几家渔业公司的渔捞日志,发现南黄海和东海的小黄鱼在地理分布上未表现出明显的地理隔离现象,认为中国近海海域小黄鱼可划分为两个群体,即渤海-北黄海群和东海-南黄海群,后期研究也支撑了这一结果[10,14];Zhang等[15]、Xiao 等[16]和Zheng 等[17]利用基因组学探究黄海和东海北部小黄鱼的种群结构,发现地理族群间的基因差异不显著,且存在高度的基因流现象。造成以上观点分歧的主要原因是所用方法不同,因此,采用当前的鱼类群体划分方法厘清小黄鱼的地理族群,对其资源科学管理及种群恢复具有重要意义。
耳石存在于硬骨鱼类内耳中,是由钙经生物矿化形成的一种碳酸钙结晶,随鱼类个体生长每日沉积且不易代谢[18]。耳石内侧面具有明显的“听沟(Sulcus acusticus)”凹槽,起到控制鱼体平衡和听觉的功能[19]。耳石及其“听沟”的形态具有种间差异,常被用于区分种间的近似种[20-21]和种内的不同地理族群[22-23]。宋大德等[6]利用耳石微化学中的耳石、锶 (Sr)/钙 (Ca) 探究了南黄海小黄鱼早期发育阶段的连通性,对连云港、盐城和南黄海外海的小黄鱼产卵群体进行了有效划分。耳石形态学作为划分鱼类群体的方法,得到了广泛应用。目前,框架分析法 (Framework analysis method) 和几何形态测量法 (Geometric morphometrics) 是耳石形态分析领域两种主要的研究方法。框架分析法主要是通过度量耳石形状参数 (长、宽、最长半径、最短半径和厚度等) 并结合耳石质量和面积,从而获得一些形态指标[24],但该方法不能有效提取耳石的一些关键形态指标信息,因此对种间区分不同地理族群的效果甚微[25-26]。几何形态测量法主要包括基于耳石轮廓的傅里叶分析法 (轮廓法) 和基于笛卡尔地标的地标点法。其中,傅里叶分析法是对耳石二维投影后通过椭圆傅里叶分析提取表征耳石轮廓信息的傅里叶系数,然后通过判别分析和主成分分析等方法区分鱼类种类或种群[24],该方法受限于耳石完整度和轮廓规则度,在耳石轮廓较不规则或有所缺损的情况下,会表现出一定的不适宜性[27];地标点法是通过分析耳石轮廓和听沟等重要区域的形态信息,将形态差异与变化定量描述和可视化,且利用多变量技术来研究多维关系的一种方法,在鱼类种类和种群区分中已得到广泛应用[24-26]。关于小黄鱼个体和耳石形态的已有研究主要采用框架分析法[28-29]和傅里叶分析法[29],尚未见有采用地标点法的研究报道。
本研究基于2022 年采自海州湾、吕泗、长江口和舟山渔场的1 龄小黄鱼样品,利用地标点法分析其矢耳石样本的形态特征,探究了南黄海和东海海域小黄鱼不同地理族群之间的连通性,以期完善小黄鱼的空间分布格局,为其管理单元的合理规划提供参考。
1 材料与方法
1.1 样品采集
为减少鱼体体长和年龄效应引起的耳石形态变化的干扰,避免因小黄鱼种群洄游而造成不同海域样本交叉带来的误差,本研究选取1 龄鱼为研究对象。康中杰等[30]比较了国内近30 年小黄鱼的von Bertalanffy 生长方程退算 (Back-calculate) 鱼体年龄的精度,发现生长方程Lt=366[1-e-0.11777(t+1.76429)][31]的退算年龄效果最佳,退算出1 龄鱼个体的体长范围为100~120 mm。为确保采集的样品历经的洄游周期一致,2022 年分别在海州湾、吕泗、长江口和舟山渔场 (图1),通过捕捞渔获筛选采集了30 尾体长符合以上退算范围的1 龄小黄鱼样品。将冷冻保存的样品运送至陆基实验室,参照GB/T 12763.6—2007《海洋调查规范》中的生物学测定方法对所选样品进行体长 (精确到0.01 mm)、体质量 (精确到0.01 g) 等测定 (表1)。对4 个海域小黄鱼体长进行单因素方差分析 (One-way ANOVA)。
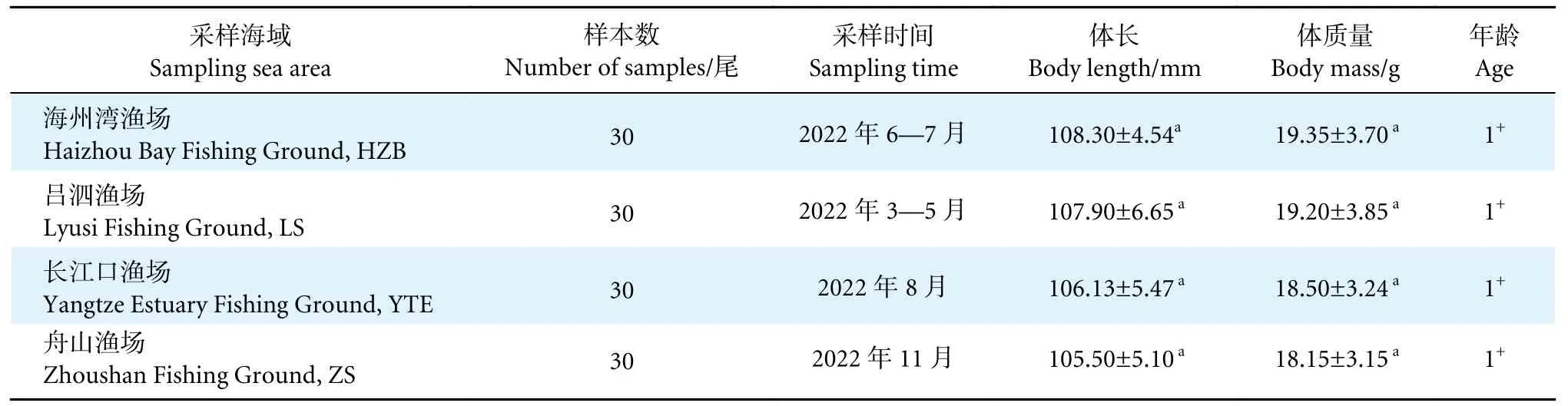
表1 4 个渔场小黄鱼样本详细信息Table 1 Detailed information of L.polyactis sampled from four fishing grounds

图1 小黄鱼采样站点及其洄游路线注:洄游路线图参考文献[32]。Fig.1 Map of sampling sites and migratory routine of L.polyactisNote: Migration route map sees Reference [32].
1.2 耳石处理
小黄鱼生物学测定完成后,解剖鱼体并用镊子摘取其矢耳石,然后去除表面有机质,乙醇清洗,烘箱干燥,最后编号保存于离心管待拍照。本研究统一选取小黄鱼左矢耳石为研究对象,首先将耳石具听沟的一侧 (内侧面)朝上放置并沿耳石最长轴水平固定,然后用体式显微镜 [Nikon-SMZ800N,尼康精机 (上海) 有限公司;配套软件NIS-Elements D 5.20.00 64-bit] 在同一放大倍率下对耳石形态进行拍摄和图像采集 (图2)。

图2 海州湾、吕泗、长江口和舟山渔场的小黄鱼左矢耳石内侧面11 个地标点位置图Fig.2 Positions of 11 landmarks on inner profile of left otolith morphology of L.polyactis sampled in Haizhou Bay (HZB),Lyusi (LS),Yangtze Estuary (YTE) and Zhoushan (ZS) fishing grounds
1.3 地标点法分析
本研究地标点软件的使用参考姜涛等[33]。1) 利用tpsdig 2.12 软件,建立小黄鱼耳石照片上地标点的坐标位置 (图2),选取小黄鱼耳石形态上的耳石轮廓端点7 个 (1—7) 和内测面“听沟”轮廓拐点4 个 (8—11),并生成相应的坐标数据文件;2) 使用tipsmall 1.2 软件对获取的数据文件进行最小平方和法则回归分析,以检验所选取地标点的有效性;3) 利用tpsRelw 1.46 软件进行普式叠加,并对地标点的坐标值进行一系列旋转和放大,计算平均形和相对扭曲得分;4) 利用tpsRegr 1.36 软件进行薄板样条分析,生成小黄鱼耳石形态网格图,并比较分析其平均形态之间的形扭曲和变化差异。
1.4 数据分析
使用Origin 2023 软件进行数据处理、主成分分析和聚类分析,对样本的体长、体质量作单因素方差分析,数据以“平均值±标准误 ()”表示,显著性水平为P<0.05。
2 结果
2.1 矢耳石形态地标点分析
对矢耳石地标点的数据文件进行最小平方回归分析,结果显示,其切空间距离 (Distance in tangent space,y轴) 与普式距离 (Procrustes distance,x轴) 之间的回归系数是0.99,验证了所选取地标点的有效性。通过tpsRelw 软件对4 个渔场120 个样本的小黄鱼矢耳石地标点分析,可以计算出11 个地标点的平均形和4 个渔场小黄鱼矢耳石地标点的重叠图。相对扭曲分析显示 (表2),在所选11 个地标点中,小黄鱼耳石听沟“尾端部”和“口端部”与中间“颈状部”的交点 (地标点8—11) 的相对扭曲贡献率占比较大 (91.88%)。其中,地标点11 的贡献率最高 (45.40%),其次为地标点10 (36.86%),表明听沟区域的地标点在4 个渔场小黄鱼的矢耳石形态区分中作用最大。
2.2 几何形态分析
基于相对扭曲的方法对4 个渔场的小黄鱼矢耳石形态地标点进行主成分分析后,共提取了18 个主成分 (表3)。其中,前3 个主成分对耳石形态总体变异的贡献率分别为89.93%、2.21%、1.96%,累计贡献率为94.10%。从二维主成分析图 (图3)可以看出,第1、第2 主成分散点图中部分海州湾渔场的群体被显著地隔离为一个单独类群,剩余部分海州湾渔场群体和其他渔场 (吕泗、长江口和舟山) 的群体高度重叠为一个类群,无法区分。从前3 个主成分的三维分析图 (图4) 可以看出,第1、第2 主成分的效果与二维散点图一致,并且部分海州湾渔场的小黄鱼和其余3 个渔场高度重叠的部分从第2、第3 主成分也未得到明显的区分。

表3 18 个相对扭曲得分的特征值和贡献率Table 3 Eigenvalues and contribution rates of 18 relative warps scores

图3 相对扭曲分数第1、第2 主成分散点图 (主成分得分×102) 和网格变形图 (变异扩大10 倍)注:HZB.海州湾渔场;LS.吕泗渔场;YTE.长江口渔场;ZS.舟山渔场。图4 同此。Fig.3 Scatterplots of relative warp scores on 1st and 2nd principal component (Principal component scores are enlarged 102 times)and grid deformation of otolith (Variation are enlarged 10 times)Note: HZB.Haizhou Bay fishing ground; LS.Lyusi fishing ground; YTE.Yangtze Estuary fishing ground;ZS.Zhoushan fishing ground.The same case in Fig.4.

图4 相对扭曲分数第1、第2、第3 主成分3D 散点图Fig.4 3D scatterplots of relative warp scores on 1st,2nd and 3rd principal component
利用tpsRegr 软件对4 个渔场的小黄鱼矢耳石形态进行地标点可视化分析,并在此基础上将结果放大10 倍后,可以清晰地看出每个地标点在网格中的变化趋势和程度 (图3)。其中,变化最明显的为耳石外轮廓上的地标点3、4、6 和耳石听沟区域的地标点8、9、10,具体表现为海州湾渔场和吕泗渔场耳石外轮廓有向外变化的趋势,且海州湾渔场更加明显。耳石听沟的整体区域有向右变化的趋势。长江口渔场和舟山渔场耳石形态向内变化,且舟山渔场变化更加明显。
2.3 聚类分析
基于耳石地标点的相对扭曲得分进行的聚类分析结果显示,4 个渔场的小黄鱼群体聚为两个类群,即吕泗、长江口和舟山渔场聚为一个类群,海州湾渔场单独聚为一个类群,其个体的欧式距离为0.9 (图5)。

图5 海州湾、吕泗、长江口和舟山渔场小黄矢耳石形态聚类分析结果Fig.5 Dendrogram for hierarchical cluster analysis by otolith morphology of L.polyactis population in Haizhou Bay,Lyusi,Yangtze Estuary and Zhoushan fishing grounds
结合主成分分析的结果,本研究表明海州湾渔场存在2 个小黄鱼种群,即海州湾渔场代表的北黄海-渤海群,吕泗、长江口和舟山渔场代表的南黄海-东海群。
3 讨论
3.1 耳石形态在种间及种内识别能力分析
鱼类种间和种内不同群体的识别是种群生物学研究的基础,而随着鱼体的生长,其耳石形态趋于稳定。自从耳石形态应用于鱼类种群划分之后,其逐步发展成为鱼类种群研究的重要手段之一[24]。同时,鱼类形态分析方法也得到长足发展,且对鱼类群体的识别能力不断提升。据Xiong 等[34]报道当小黄鱼体长达到130 mm 时,其矢耳石形态趋于稳定,推测耳石形态可能于首次性成熟时发生了显著变化。Jiang 等[35]研究指出小黄鱼第一次性成熟发生在1 龄左右,根据本文所用公式反演退算出1 龄小黄鱼体长为100~120 mm,而体长为130 mm 时小黄鱼已经达到2 龄。因此,笔者认为小黄鱼耳石形态的显著变化可能发生于2 龄左右,并非在第一次性成熟时。究其原因,可能是以往报道中体长约130 mm 的1 龄小黄鱼使用了不同的年龄鉴定方法及生长方程所致。
本文利用地标点法对南黄海和东海小黄鱼耳石形态进行分析,结果发现在海州湾渔场存在两个耳石形态差异明显的小黄鱼群体,即北黄海-渤海群体和南黄海-东海群体,且发现其耳石形态差异主要来源于耳石听沟区域,对比这两个小黄鱼群体的耳石形态发现,北黄海-渤海群体小黄鱼耳石听沟上的地标点8、9、11 均有向耳石“颈状部”内部变化的趋势,说明该群体小黄鱼耳石听沟的“颈状部”相较于南黄海-东海群体有变短的趋势。Jiang等[10]通过比较海州湾渔场的小黄鱼成鱼的耳石形态和鱼卵大小发现,在海州湾渔场存在两个小黄鱼种群,且耳石形态总判别率达91.7%。耳石形态不仅在种内具有高识别能力,对鱼类种类也具有较好的识别效果。姜涛等[33]采用地标点法分析九龙江口和珠江口的凤鲚 (Coiliamystus) 与七丝鲚(C.grayii) 的耳石形态差异,结果显示两个鱼种的总正确判别率高达100.0%。张申增等[36]对红鳍笛鲷 (Lutjanuserythopterus) 和紫红笛鲷 (L.argentimaculatus) 的耳石形态进行分析,其物种总判别率达82.8%。不同群体之间显著的耳石形态差异是取得较高识别效果的基础,耳石形态差异除了与耳石不同的年轮宽度增量和轮廓因子有关外,还与耳石内侧面的听沟区域形态有较大关系。侯刚等[37]在基于地标点法对北部湾4 种白姑鱼矢耳石形态识别中,选取位于听沟区域的地标点总贡献率 (95.08%)远高于外轮廓上地标点总贡献率(4.12%),其矢耳石形态差异主要表现在听沟的“颈部”、“头部”和“尾部”。本文中所选取听沟区域地标点8—11 的总体贡献率 (91.88%) 也远高于耳石轮廓上地标点1—7 的总体贡献率 (8.12%),其矢耳石形态差异主要来源于听沟两端“口端部”和“尾端部”与中间“颈状部”的交点 (地标点8—11)。
耳石形态受到遗传和环境的共同作用[38]。目前,利用分子生物学手段来区分鱼类物种已被广泛认可和使用,但是对遗传分化不明显的同一物种不同地理群体,采用分子生物学手段来划分群体有一定的局限性[16-17,39],而对这类群体利用鱼类耳石形态学来划分种群则相对容易且更具优势[24-26]。地标点法可以有效提取耳石的大部分形态信息,目前鱼类种内和种间的识别效果可基本满足需求。然而,该方法只是分析鱼类耳石具有听沟区域的一个面,限制于二维平面,对耳石整体形态信息无法提取和分析。随着现代高精度仪器的涌现和3D 技术的逐步成熟,提取鱼类耳石整体形态信息已成为可能。Gu 等[40]基于X 射线微计算机断层扫描技术 (Micro-CT) 和扫描电子显微镜 (SEM) 分析了黄唇鱼(Bahabataipingensis) 矢耳石,发现其背面具有许多细长的棱柱晶体构成晶体簇,初步揭示了黄唇鱼耳石的声学功能结构。Martí-Puig 等[41]使用三维耳石图像分析方法,将耳石的整体形态信息以数据集的形式呈现,形态学与三维模型的结合将为鱼类形态学研究提供深层次的分析和更有力的依据。
3.2 南黄海和东海的小黄鱼种群划分
本研究基于在耳石外轮廓和听沟上建立的11 个地标点提取耳石形态特征信息,经分析显示南黄海和东海海域存在两个在耳石内侧面听沟区域形态差异显著的小黄鱼种群,即北黄海-渤海群和南黄海-东海群,且在海州湾渔场存在这两个种群的混栖群体。
每年随着水温的上升,小黄鱼便逐渐向近岸产卵洄游,产卵高峰期存在着自南向北的纬向滞后差异,例如舟山渔场小黄鱼的产卵高峰期一般在3 月,吕泗渔场则在4 月[32]。小黄鱼产卵后会向外海洄游至机轮拖网禁渔区线附近索饵,小黄鱼鱼卵此时完全不具备自主游泳能力,其鱼卵—仔鱼阶段基本是处于“随波逐流”的状态[42],因此,分析当时的海洋动力环境对深入理解小黄鱼种群结构具有重要意义。2018 年3—7 月,对长江口邻近海域的小黄鱼鱼卵、仔稚鱼的调查结果显示:3 月其鱼卵主要分布在机轮拖网禁渔区线内舟山渔场北部水域 (水深10~30 m),4 月鱼卵遍布禁渔区线内长江口渔场,5 月鱼卵和仔鱼重心偏移至吕泗渔场,从整体看小黄鱼鱼卵、仔鱼的分布重心是逐渐北移的[43]。物理海洋动力机制分析可知:3 月,高温高盐的台湾暖流北上至舟山渔场,为小黄鱼产卵创造了良好的环境[44];4 月,随着长江口渔场水温的上升,受台湾暖流输送影响,小黄鱼鱼卵、仔鱼遍布在禁渔区线内的长江口渔场[45];5 月,在强劲的东海潮汐和盛行的东亚季风共同作用下,一部分长江冲淡水会持续北上至苏北浅滩邻近海域,此时禁渔区线内的长江口渔场和吕泗渔场便成了南黄海-东海小黄鱼种群的主要育幼场[46]。综上可知,南黄海-东海小黄鱼种群鱼卵、仔鱼阶段主要受台湾暖流和长江冲淡水等流系控制,并不存在长江冲淡水会在此形成一道隔离东海和南黄海小黄鱼群体的屏障。
每年的4 和5 月分别是吕泗渔场和海州湾渔场小黄鱼的产卵高峰期[34],此时,在北向的潮致余流输运下[47-48],吕泗渔场会有部分小黄鱼鱼卵、仔鱼被输送至海州湾邻近水域,在海州湾渔场形成两地的小黄鱼混栖群体。黄康康等[49]通过水动力物理模型模拟吕泗渔场小黄鱼鱼卵输运路径发现,在吕泗渔场不同年份释放的粒子,其输运走向基本偏北朝向海州湾渔场;Zhang 等[50]通过比较海州湾渔场小黄鱼耳石年轮宽度变量和耳石轮廓等因子发现,在海州湾渔场存在两个来自不同越冬场的小黄鱼群体。由此可知,北黄海-渤海种群和南黄海-东海种群之间并未形成真正的地理隔离,且存在较高的个体交流现象,说明海州湾渔场是小黄鱼不同群体之间个体交流的主要海域之一。这与众多探究小黄鱼种群基因结构的发现一致,即各小黄鱼地理群体间不存在显著的基因差异[16-17,38]。除了两个种群栖息水域环境造成的影响外,笔者认为,两个种群摄食特征差异也可能是导致其耳石形态存在显著性差异的原因。例如:在长江口渔场,小黄鱼主要以甲壳类和鱼类为食,优势种包括糠虾、磷虾、七星底灯鱼 (Benthosemptterotum)、脊腹褐虾 (Crangon affinis) 和鹰爪糙对虾 (Trachysalambriacurvirostris)等;而在海州湾渔场,小黄鱼主要以太平洋磷虾(Euphausiapacifica)、脊腹褐虾 (C.affinis)、细螯虾(Leptochelagracilis) 和赤鼻棱鳀 (Thryssakammalensis) 等为食[51]。
猜你喜欢
保健与生活(2023年23期)2023-12-26
中国水产(2021年12期)2021-12-06
大自然探索(2021年12期)2021-02-07
中华养生保健(2020年7期)2020-11-16
小读者(2020年4期)2020-06-16
当代水产(2019年4期)2019-05-16
老友(2018年7期)2018-08-08
科学生活(2017年9期)2017-10-16
作文与考试·小学高年级版(2017年9期)2017-05-16
渔业致富指南(2016年12期)2016-11-11
