
BDNF/TrKB介导的突触丢失对创伤性脑损伤小鼠远期认知障碍的影响及机制
2023-12-21 07:14孙衍昶冯基高欧阳一彬刘达远海南医学院附属第二医院神经外科海南海口570311
中国老年学杂志 2023年24期
孙衍昶 冯基高 欧阳一彬 刘达远 (海南医学院附属第二医院神经外科,海南 海口 570311)
创伤性脑损伤(TBI)是由外部物理力量引起的脑功能损伤或其他脑病理特征〔1〕。在中国,TBI 患者的死亡率为 13/10 万人,虽然近年来TBI发病率有所下降,但由于TBI病理复杂性,中枢神经系统常出现不可逆性损伤,仍会有约30%的颅脑损伤患者死于继发性二次损伤,致残致死率高〔2〕。
研究发现,TBI发生过程中会产生肿块损伤、挫伤、弥漫性轴索损伤及一系列可以造成神经元损伤的机制,如缺血、细胞凋亡、线粒体功能障碍、皮质扩散抑制和微血管血栓形成等,导致不同的临床病理特征〔3〕。尽管对 TBI 机制的大多数研究都集中在神经元损伤上,但有研究〔4〕表明,突触及其内在分子通路在多种脑部疾病中发挥重要作用,这增加了突触与 TBI 相关的可能性。且多项研究发现〔5~7〕,在TBI动物模型中可观察到突触丢失的现象,进一步提示了突触功能在TBI中的重要作用。脑源性神经营养因子(BDNF) 是哺乳动物神经系统中研究最广泛的神经营养因子,可促进感觉神经元的生长和分化及成人的突触调节〔8〕。BDNF与酪氨酸激酶受体(Trk)B作为调控中枢神经系统的重要通路,参与帕金森病、阿尔茨海默病、抑郁症、癫痫和慢性疼痛等〔9〕多种疾病的发生发展过程。并且,许多证据表明了BDNF在TBI中发挥重要作用,如王国强等〔10〕研究发现,腺病毒介导的 BDNF 基因过表达可保护TBI小鼠的海马神经元;康雅琴等〔11〕发现异氟醚处理后通过调节小鼠脑组织胰岛素样生长因子(IGF)-1、BDNF、神经生长因子(NGF)表达对新生小鼠缺血缺氧性脑损伤具有保护作用,可改善小鼠远期认知功能。但是目前还不清楚BDNF-TrkB参与TBI的具体机制,且是否和突触丢失存在联系亦不清楚。故本实验拟在TBI中探究BDNF-TrkB信号通路与突触功能的关系,并阐明其对脑损伤后远期认知功能的影响及其机制,为TBI临床治疗提供理论基础。
1 材料与方法
1.1主要材料 定量聚合酶链反应(qPCR)引物合成于金唯智公司;腺病毒过表达/敲低载体由上海吉玛基因公司制备;原位细胞凋亡检测试剂盒购自Roche公司;一抗p-磷脂酰激醇-3激酶(PI3K)、PI3K、蛋白激酶B(AKT)、p-AKT、B细胞淋巴瘤(Bcl)-2、Bcl-2相关X蛋白(Bax)、cleaved-半胱天冬蛋白酶(caspased)3、BDNF、TrkB、p-TrKB(Tyr516)、突触蛋白(SYN)1、SYNA、突触素(Synaptophysin)、突触后致密蛋白(PSD)95、二抗抗兔IgG、辣根过氧化酶(HRP)-相关抗体、二抗抗鼠IgG、HRP相关抗体等均购自CST;荧光二抗山羊抗兔IgG H&L(Alexa Fluor® 488)、山羊抗鼠IgG H&L(Alexa Fluor® 594)购自Abcam公司。
1.2动物分组及模型制备 雄性C57BL/6小鼠240只,8~10周龄,体质量(28.0±2.0)g,由海南医学院实验动物中心提供,动物实验经医院伦理会批准。分为Sham组、TBI组、TBI_OE_NC组、TBI_OE_BDNF组、TBI_Sh_NC组、TBI_Sh_BDNF组,每组40只。采用控制性皮质撞击(CCI)法〔12〕构建TBI小鼠模型。以甲苯噻嗪(10 mg/kg) 和氯胺酮(75 mg/kg)腹腔注射麻醉,麻醉稳定后去除小鼠顶部毛发,俯卧位固定于立体定位仪,碘伏消毒头顶部位,作正中切口,分开头皮剥离骨膜,在右侧冠状缝和人字缝之间、中线旁2 mm处颅骨钻开直径为3 mm的骨窗,将小鼠头部固定于立体定位仪,应用CCI打击仪,设定参数为:打击速度1.5 m/s,深度1.5 mm,停留时间100 ms。打击后,还纳骨瓣,缝合小鼠头皮,无菌棉纱布包扎。手术完成后将动物置于加热垫上,待其恢复意识后,放回笼中。Sham组麻醉后只暴露骨窗,不做打击。腺病毒注入方式:TBI_OE_NC组和TBI_OE_BDNF组于CCI打击后立即在骨窗中心处导入10 μl微量注射器针头,深度3 mm,以1 μl/min的速度分别注入AAV_OE_NC和AAV_OE_BDNF病毒缓冲液3 μl,然后进行还纳骨瓣,缝合小鼠头皮,无菌棉纱布包扎。TBI_Sh_NC组、TBI_Sh_BDNF组麻醉后只暴露骨窗,不做打击,以同样的位置和方式分别注射AAV_Sh_NC和AAV_Sh_BDNF病毒缓冲液3 μl。
1.3qPCR检测目的基因mRNA表达 Trizol法提取细胞RNA,1%琼脂糖凝胶电泳检测RNA完整性,Nanodrop2000测定RNA的浓度与纯度,0.8 μg RNA逆转录成cDNA,qPCR体系(20 μl)为1 μl逆转录产物、10 μl SYBR Green Mix、上下游引物(10 μmol/L)各0.4 μl、8.2 μl ddH2O。预变性95 ℃、5 min,94 ℃变性30 s、60 ℃退火30 s、72 ℃延伸 1 min,40个循环后终延伸完成PCR。内参选用β-actin,以2-△△Ct法计算目的基因表达量。实验独立重复3次。 引物序列(3′-5′):ACTB正向引物:CAGCACAGAGCCTCGCCTTT,反向:AGACAAGAG-ACCCCGCCGGTT;BDNF正向引物:ATGTGATGCCCTCGGATTATC,反向:GAGGAAGAGTAGCCAAGTGTG;TrkB正向引物:GCACTCACCTCTTCAGAACG,反向:CATCTTTGGAAGGTTCAGGTTG。
1.4Western印记法检测蛋白表达水平 收集目标小鼠脑部组织,使用冷冻研磨仪进行组织匀浆,14 000 r/min离心10 min收集上清即为总蛋白,二喹啉甲酸(BCA)法测定蛋白浓度。每组样品取30 μg蛋白进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),湿转法将目的蛋白转至聚偏氟乙烯(PVDF)膜上,5%脱脂奶粉室温封闭2 h,一抗4 ℃过夜孵育;次日,磷酸盐吐温缓冲液(PBST)洗涤3次,二抗室温孵育2 h;PBST洗涤后加入电化学发光(ECL)液进行显影。
1.5TUNEL法凋亡检测 严格参照原位细胞凋亡检测试剂盒操作说明书进行实验,简言之,对小鼠脑部组织切片进行透化,并在37 ℃下与 TUNEL反应混合物一起孵育60 min。磷酸盐缓冲液(PBS)洗涤3 min后,将样品在 0.1%4′,6-二脒基-2-苯基吲哚(DAPI)中孵育5 min,然后在 PBS 中洗涤。使用蔡司LSM700激光共聚焦显微镜观察并拍照。
1.6免疫荧光检测 将脑片放到12孔板,加3%双氧水封闭10 min,中杉金桥超敏二步法试剂后PBS 洗两遍,将脑片周围的水用吸水纸或卫生纸擦净,一抗Synaptophysin(1∶100)、BDNF(1∶200)稀释,加入稀释后的一抗,4 ℃过夜,次日PBS 洗净两遍后参照中杉金桥超敏二步法试剂盒说明书完成。使用蔡司LSM700激光共聚焦显微镜观察并拍照。
1.7Morris 水迷宫实验 各组分别于造模后28 d进行定位航行实验。将圆形池分为4个象限。跟踪摄像机放置游泳池上方,用于记录小鼠运动轨迹。随机选择象限将小鼠面向池壁放入水迷宫,计算机跟踪分析小鼠寻找目标平台(第一象限中水面下方2 cm处)的潜伏期,定90 s为最长时间,若90 s还没到达,人为将小鼠置于平台20 s,记录逃避潜伏期为90 s。每只动物训练5次/d,每次训练之间间隔10 min,连续训练5 d。于造模后32 d进行空间探索实验,最后一次训练结束后移去平台,将小鼠面向池壁放在第三象限,计算机记录小鼠90 s内的游泳速度及路径,计算目标象限穿越次数。
1.8统计学处理 采用SPSS20软件进行t检验、单因素方差分析,GraphPad8.0软件绘制相关图片。
2 结 果
2.1BDNF/TrkB在TBI中表达降低 透射电镜结果显示,与Sham组相比,TBI组神经元突触后致密物厚度(5、7 d)和突触体密度(3、5、7 d)显著降低(P<0.05)。见表1。qPCR及Western印迹结果显示,TBI组脑组织中BDNF和TrkB mRNA和蛋白水平随时间逐渐降低(P<0.05)。见表2、图1。
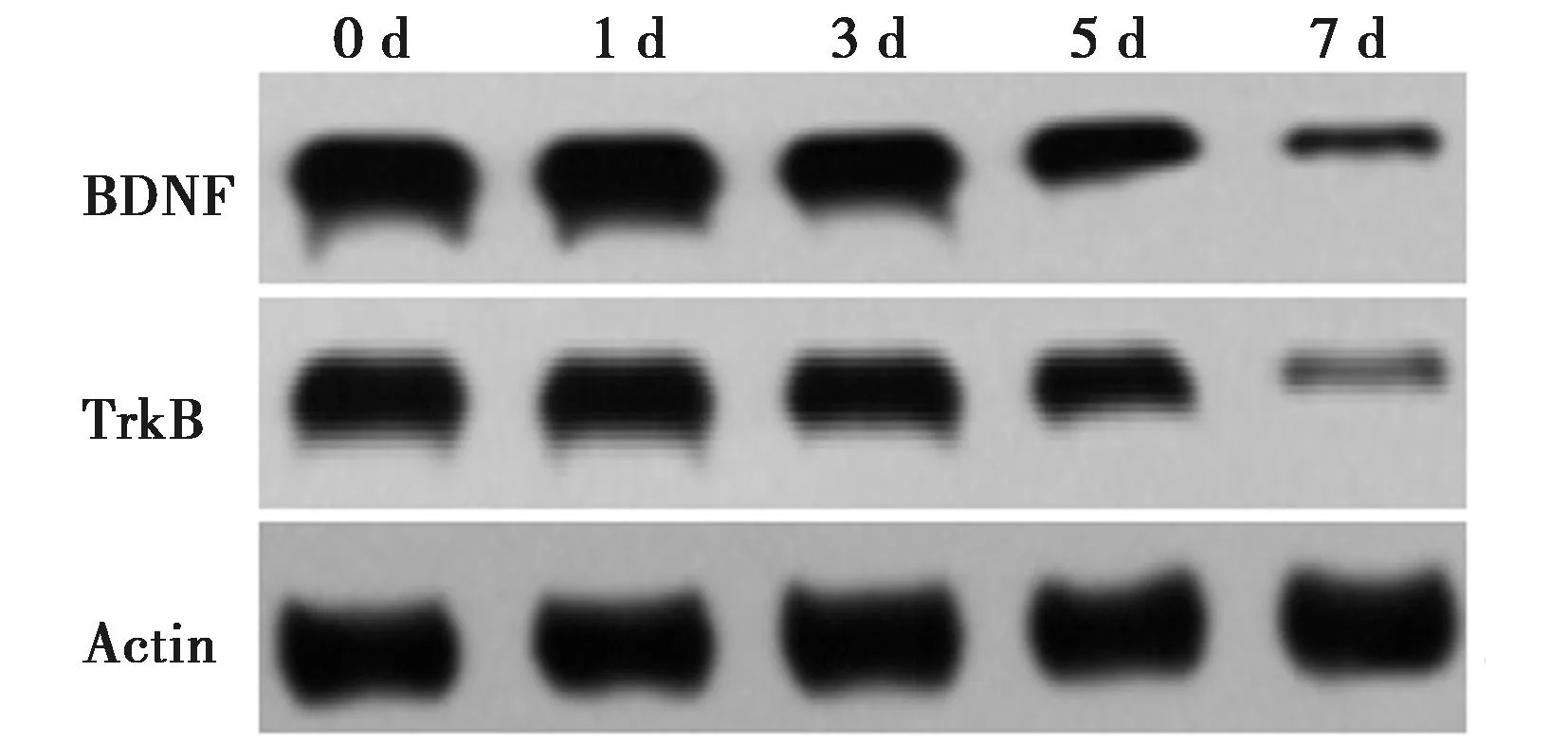
图1 Western印迹检测TBI组BDNF/TrkB蛋白表达

表1 两组神经元突触后致密物厚度及突触体密度
表2 TBI组脑组织中BDNF、TrkB相对mRNA表达
2.2过表达BDNF增加TBI小鼠模型海马体中突触数量 与Sham组比较,TBI_OE_NC组海马体BDNF表达显著减少;而TBI_OE_BDNF组BDNF蛋白表达水平显著高于TBI_OE_NC组(P<0.05)。TBI_OE_BDNF组突触数量显著多于TBI_OE_NC组(P<0.05),但与Sham组无统计学差异(P>0.05)。相较于TBI_OE_NC组,TBI_OE_BDNF组脑海马体组织中TrkB、p-TrkB、SYN1、SYNA、PSD95蛋白表达水平显著提高(P<0.05),与Sham组水平相当(P>0.05)。见表3、图2、图3。
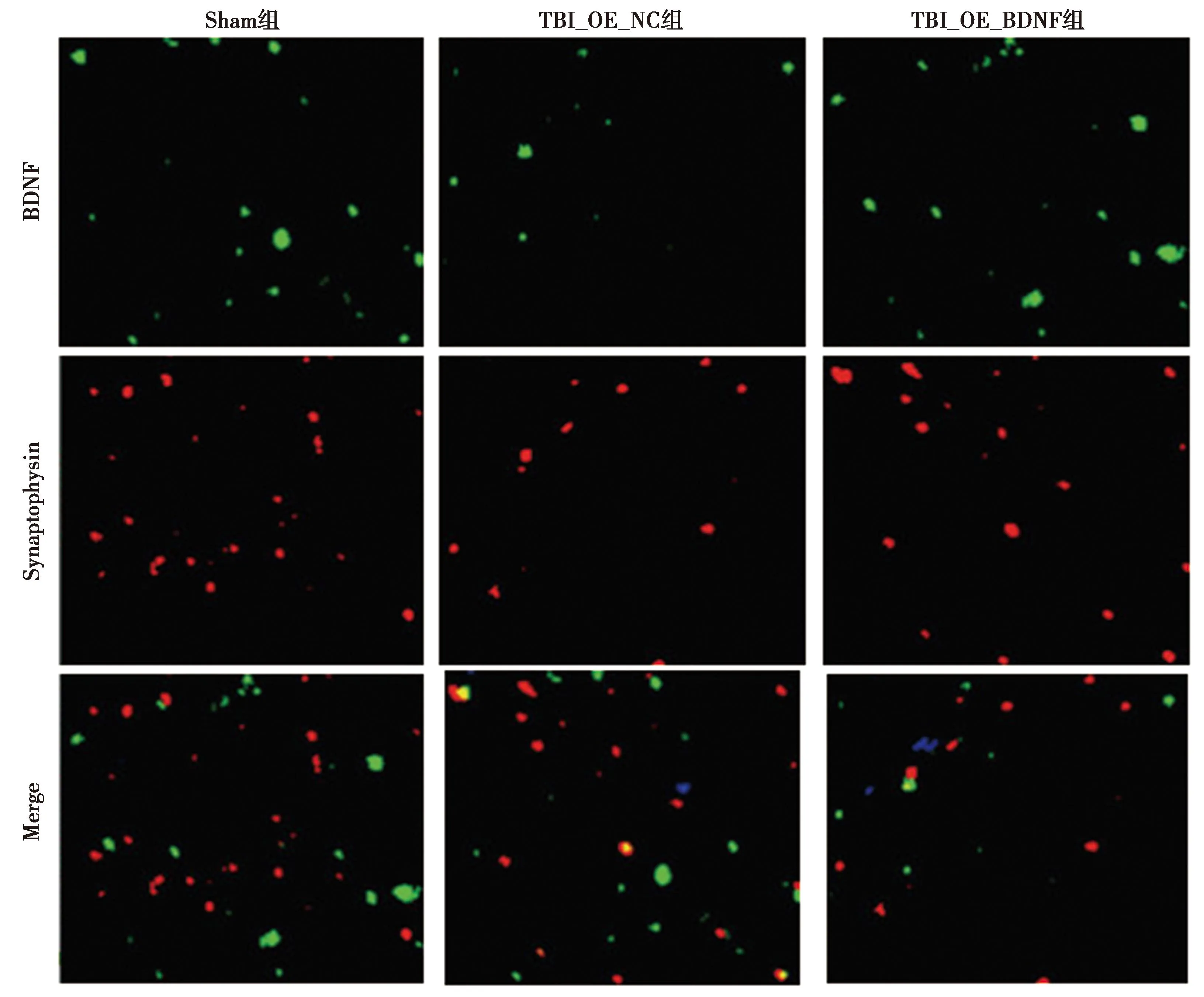
图2 免疫荧光检测各组海马体中突触数量(×200)

1~3:Sham组、TBI_OE_NC组、TBI_OE_BDNF组;图6同图3 Western印迹检测各组BDNF/TrkB及突触相关蛋白表达
表3 各组突触数量、TrkB、p-TrkB、SYN1、SYNA、PSD95蛋白表达比较
2.3过表达BDNF提高TBI小鼠模型远期认知能力 与Sham组比较,TBI_OE_NC组逃避潜伏期(3、4、5 d)显著升高,目标平台驻留时间、平台穿越次数及游泳速度显著降低(P<0.05)。与TBI_OE_NC组比较,TBI_OE_BDNF组逃避潜伏期(2、3、4、5 d)显著降低,目标平台驻留时间、平台穿越次数及游泳速度显著升高(P<0.05)。见表4。
表4 过表达BDNF对各组水迷宫实验的影响
2.4过表达BNDF抑制小鼠海马体组织细胞凋亡水平 相较于Sham组〔(2.31±0.13)%〕,TBI_OE_NC组细胞凋亡水平〔(32.3±3.78)%〕显著升高;而TBI_OE_BDNF组细胞凋亡水平〔(9.33±1.53)%〕显著低于TBI_OE_NC组。见图4。Western印迹结果显示,相较于TBI_OE_NC组,TBI_OE_BDNF组脑海马体组织中PI3K、AKT蛋白表达水平无明显差异(P>0.05),p-PI3K、p-AKT、Bcl-2蛋白表达显著升高(P<0.05),Bax、cleaved-caspased3蛋白水平显著降低(P<0.05)。见表5、图5。

图4 TUNEL法检测各组海马组织细胞凋亡(×400)
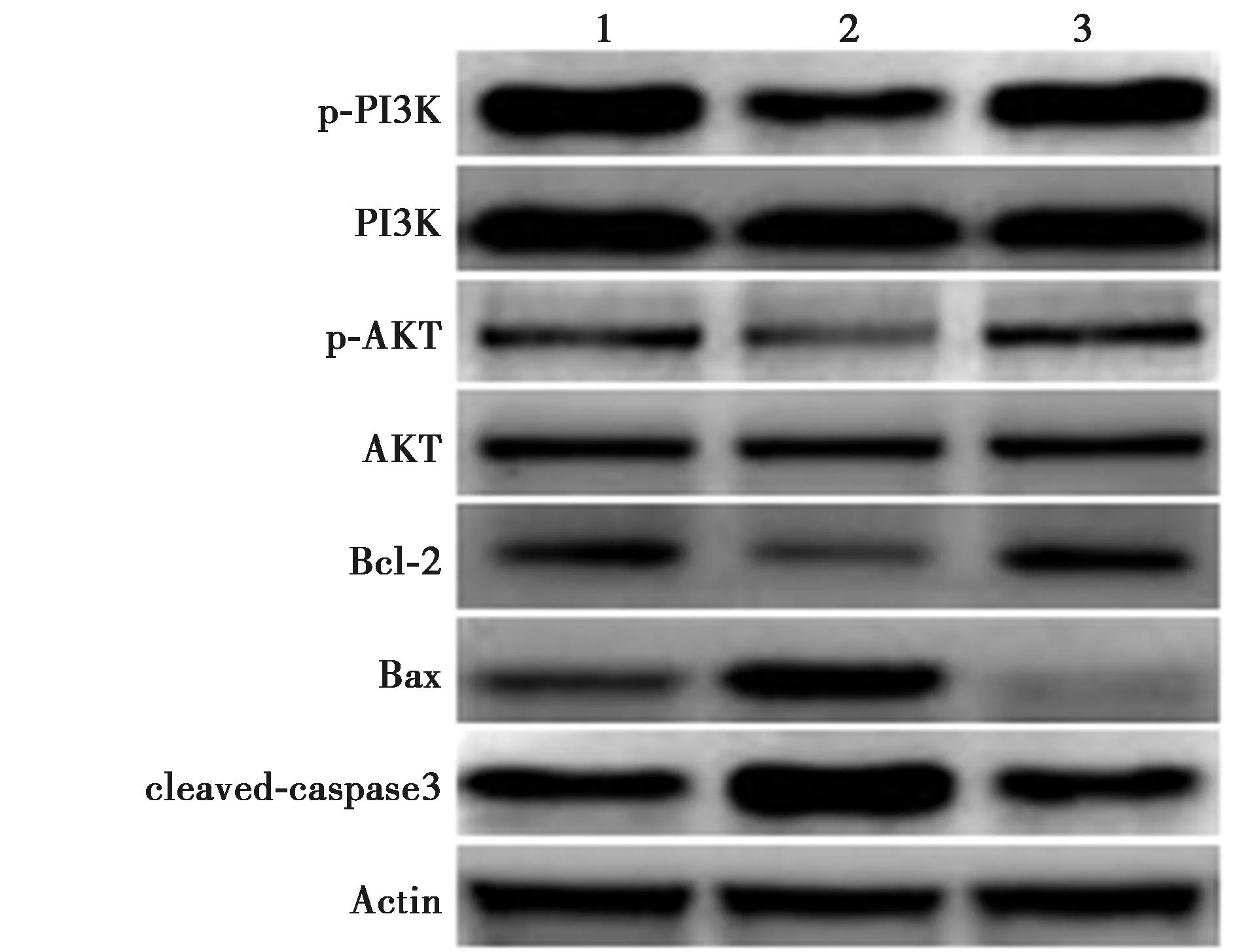
图5 Western印迹检测各组PI3K/AKT相关蛋白表达
表5 各组PI3K、p-PI3K、AKT、p-AKT、Bcl-2、Bax、Cleaved-caspase3蛋白表达比较
2.5敲低BDNF损伤小鼠的远期认知能力 相较于TBI_Sh_NC组(164.00±13.11),TBI_Sh_BDNF组海马体中突触数量(84.00±13.15)显著降低(t=7.47,P=0.002)。Morris 水迷宫结果显示,相较于TBI_Sh_NC组,TBI_Sh_BDNF组逃避潜伏期(3、4、5 d)显著增长(P<0.05),目标平台驻留时间、平台穿越次数及游泳速度显著降低(P<0.05)。见图6、表6。
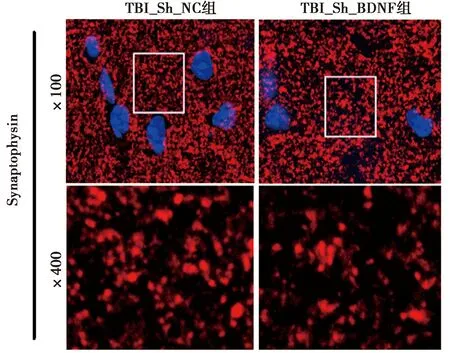
图6 共聚焦显微镜观察两组海马Synaptophysin
表6 敲低BDNF对两组水迷宫实验的影响
3 讨 论
TBI是晚年痴呆的危险因素,关于TBI的具体机制,之前的研究大多侧重于神经元损伤方面,随着突触及其内部分子通路研究的增多,越来越多的证据表明,突触在脑部疾病中发挥重要作用〔13〕。对发育、精神和神经系统疾病患者的突触蛋白质组和基因组测序的研究表明〔14~16〕,超过 130 种脑部疾病是由突触蛋白突变引起的。BDNF在中枢神经系统中广泛表达,可在发育过程中促进某些神经元群的存活和分化。在成年期,BDNF 可以调节神经元突触强度,并与学习和记忆的海马机制和疼痛的脊髓机制有关,是一种有效的神经保护剂,因此被认为是理想的脑部疾病药物靶点。BDNF 与高亲和力受体 TrkB 结合后,受体二聚化、自身磷酸化〔17〕,进而促进多种细胞内途径的激活:ras-丝裂原活化蛋白激酶〔18,19〕、磷脂酶 Cγ〔20~22〕、 磷脂酰肌醇 3' 激酶〔23,24〕、钙/钙调蛋白依赖性蛋白激酶〔25〕和环磷酸腺苷(cAMP)反应元件结合蛋白磷酸化,诱导部分蛋白质合成及突触可塑性,影响学习和记忆的形成。本研究发现,BDNF-TrkB信号通路与TBI后发生的突触丢失有着密切联系,进一步研究发现,BDNF-TrkB信号通路对突触内部蛋白调控作用及可突触重塑中的作用。
本研究Morris 水迷宫结果表明,BDNF对小鼠海马体突触密度及学习和记忆能力具有支持作用。突触丢失的主要原因是PSD中蛋白质失调引起的,而BDNF-TrkB通路可调控miRNA表达、局部蛋白质合成来调控突触的可塑性。BDNF-TrkB可激活PI3K/Akt 相关通路发挥抗细胞凋亡和促存活活性,并调节 N-甲基-d-天冬氨酸受体(NMDAR) 依赖性突触可塑性。本研究表明,BDNF过表达后可激活下游PI3K/AKT通路,促进抗凋亡相关蛋白表达,从而抑制细胞凋亡,保护突触功能。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
作文周刊·小学二年级版(2022年20期)2022-05-05
自然杂志(2021年6期)2021-12-23
南京林业大学学报(自然科学版)(2021年5期)2021-10-13
创新作文(小学版)(2019年10期)2019-09-25
现代装饰(2018年5期)2018-05-26
小学生学习指导(低年级)(2017年5期)2017-05-04
广西林业科学(2016年3期)2016-03-16
电源技术(2015年5期)2015-08-22
