
基于基因分型测序(GBS)技术的堇叶紫金牛遗传结构分析
2023-12-16 12:34:42周鑫洋夏国华马丹丹陈涛梅姚熠涵
植物资源与环境学报 2023年6期
周鑫洋, 饶 盈, 夏国华, 马丹丹, 陈涛梅, 姚熠涵
(1. 浙江农林大学林业与生物技术学院 浙江省森林芳香植物康养功能研究重点实验室, 浙江 杭州 311300; 2. 浙江省杭州市临安区天目山林场, 浙江 杭州 311311; 3. 浙江农林大学暨阳学院, 浙江 绍兴 311800)
堇叶紫金牛〔Ardisiaviolacea(T. Suzuki) W. Z. Fang et K. Yao〕隶属于报春花科(Primulaceae)紫金牛属(ArdisiaSw.),仅分布于浙江、台湾和安徽[1],其种群呈岛屿状间断分布,是中国特有的濒危植物[2]。因果实红艳、挂果时间长,堇叶紫金牛具有很高的观赏和应用价值。此外,堇叶紫金牛对研究中国大陆和台湾植物区系演化、紫金牛属的分组及系统发育具有重要的科研价值;该种还是紫金牛属植物栽培育种及品种改良的野生基因库,对维持紫金牛属植物的遗传多样性具有重要意义。但由于生境片断化以及自身的生理生态学特性,堇叶紫金牛野生居群更新受到严重干扰,居群数量急剧减少,因此,对其开展保护生物学研究已迫在眉睫。目前,对堇叶紫金牛濒危机制的研究主要集中在种群结构特征[3-5]、繁殖技术[6-8]以及生理生态[9-10]方面,而关于遗传结构方面的研究尚未见报道。
基因分型测序(genotyping by sequencing,GBS)技术是一种简化基因组测序技术。虽然GBS技术不能深度覆盖全基因组,但其具有测序通量高、成本低等优点[11-12]。GBS技术自2011年在玉米(ZeamaysLinn.)和大麦(HordeumvulgareLinn.)研究中首次应用以来[13],已经广泛应用于资源分类、遗传分析及基因定位等领域,在不实野燕麦(AvenasterilisLinn.)[14]、茶〔Camelliasinensis(Linn.) O. Ktze.〕[15]、稻(OryzasativaLinn.)[16]、硬粒小麦〔Triticumturgidumsubsp.durum(Desfontaines) Husnot〕[17]等植物中得到了应用。GBS技术结合全基因组关联分析,有助于发现植物的新性状及其相关变异来源,对后续优良品种的选育和改良具有指导意义[18]。
遗传多样性高低对濒危植物能否长期存活具有重要影响[19],因此对濒危植物遗传多样性的研究有助于了解物种遗传背景,揭示物种濒危机制。为此,本研究采用GBS技术对堇叶紫金牛13个野生居群77份样本进行单核苷酸多态性(single nucleotide polymorphism,SNP)位点挖掘,在此基础上,对各居群的遗传多样性、系统发育树、亲缘关系等进行了分析,以期为堇叶紫金牛的物种保护和利用及种质资源保存提供参考依据。
1 材料和方法
1.1 材料
根据堇叶紫金牛野生资源的分布状况和居群大小,以相邻植株间隔超过20 m作为划分居群的依据,在堇叶紫金牛全分布区内13个野生居群中共采集77份样本,每个居群选取健康的成年植株,每株采集2~3枚健康幼嫩的叶片,液氮速冻后于-80 ℃保存、备用。各居群基本信息见表1。

表1 堇叶紫金牛13个居群的基本信息Table 1 Basic information of 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao
1.2 方法
1.2.1 基因组DNA提取与GBS文库构建 采用Axyprep DNA提取试剂盒〔康宁生命科学(吴江)有限公司〕提取堇叶紫金牛DNA,采用质量体积分数2%的琼脂糖凝胶电泳分析DNA完整性,采用Qubit 2.0荧光定量仪(美国赛默飞世尔科技公司)精确定量DNA浓度;用RsaⅠ和HaeⅢ将检验合格的DNA样品酶切消化处理成小片段,小片段经过末端修复、加A尾、加测序接头、纯化、PCR扩增等过程完成GBS文库制备。根据预设方案对文库进行质量体积分数2%的琼脂糖凝胶电泳后割胶纯化,选择目标区间内相应的文库进入后续测序。文库质检合格后上机进行高通量测序,测序平台为Illumina Nova 6000,测序模式为PE150。
1.2.2 基因组SNP位点挖掘 采用FastP 0.20.0软件对测序得到的所有read进行质控,去除接头,去除含有无法确定碱基信息的比例大于5%的read,去除低质量read(质量值Q小于等于10的碱基占比在20%以上),统计原始测序量、有效测序量、错误率在1%以下的碱基占比(Q20)、错误率在0.1%以下的碱基占比(Q30)、GC含量,并进行综合评价,过滤后获得clean data。之后对所有样本的read进行聚类,将聚在一起的read作为1个标签,并生成一致性序列。采用BWA 0.7.13软件进行序列一致性比对,并使用GATK 3.8.1软件和Samtools软件共同进行SNP检测。按照最小等位基因频率大于0.05、数据完整度大于60%(即不超过40%个体基因型缺失)的标准对检测到的SNP进行过滤。
1.3 数据分析
采用R语言扩展包Genepop v1.0.5计算观测杂合度、多态信息含量、居群间的遗传距离和遗传分化系数;采用vcftools v0.1.14软件计算核苷酸多样性指数、Tajima’s D值、Shannon’s多样性指数;采用MEGA 5.0软件的Neighbor-joining法基于p-distance模型构建系统发育树,自展重复抽样次数为1 000次;采用Admixture 1.3.0软件分析群体结构;采用EIENSOFT 6.0.1软件进行主成分分析;采用SPAeDi 1.4软件进行亲缘关系分析;采用TreeMix v1.01软件进行基因流分析。
2 结果和分析
2.1 SNP位点挖掘
分析结果显示:77份堇叶紫金牛样本共获得421.37 G有效高质量测序数据,平均每份样本获得5.47 G有效数据,有效数据率98.9%。Q20值为96.0%~98.1%,均值为97.5%,Q30值为89.6%~94.0%,均值为92.7%;GC含量为40.5%~46.0%,均值为43.1%,表明测序数据质量较高,可以用于后续分析。77份样本中获得高质量序列最多的是浙江省杭州市临安区顺溪坞的1个居群(HLS2)的第1份样本,有54 816 017条序列,最少的是安徽省黄山市祁门县牯牛降(HQG)居群的第3份样本,有21 527 449条序列。平均每份样本获得高质量序列36 557 933条,平均比对率为99.1%。
对测序数据进行分析,共获得有效SNP位点246 307个,每份样本检测到SNP位点1 154~3 789个(均值为3 199)。浙江省杭州市临安区顺溪坞的另1个居群(HLS1)的平均SNP位点数最多,有3 652个,浙江省舟山市定海区蔡家岙(ZDC)居群的平均SNP位点数最少,仅有2 138个。77份样本中有效SNP位点的杂合率为11.7%~37.0%,均值为26.6%;有效SNP位点的纯合率为63.0%~88.3%,均值为73.5%。
2.2 遗传多样性分析
对筛选获得的SNP位点进行统计分析,评价不同居群的遗传多样性,结果见表2。结果显示:13个居群的观测杂合度为0.156 9~0.428 9,均值为0.353 7,表明堇叶紫金牛基因整体稳定性较差;13个居群的多态信息含量为0.078 5~0.324 4,均值为0.263 4;核苷酸多样性指数为0.000 2~0.000 7,均值为0.000 3,表明堇叶紫金牛的核苷酸多样性较低;13个居群的Tajima’s D值为0.224 7~1.093 6,均大于0,表明堇叶紫金牛各居群受平衡选择程度较高;13个居群的Shannon’s多样性指数为0.217 5~0.664 9,均值为0.549 4,其中安徽省黄山市祁门县牯牛降(HQG)、浙江省舟山市定海区蔡家岙(ZDC)和浙江省宁波市象山县屠家园村(NXT)居群的Shannon’s多样性指数较大,表明这3个居群的遗传多样性相对较高。总体来看,堇叶紫金牛遗传多样性水平偏低,各居群间遗传多样性存在一定差异。
2.3 系统发育树分析
利用鉴定到的高质量SNP对堇叶紫金牛13个居群77份样本进行系统发育树分析,结果见图1。结果显示:77份样本可分为6组,Ⅰ组包含浙江省杭州市临安区顺溪坞的2个居群(HLS1、HLS2)和浙江省杭州市西湖区云栖竹径的2个居群(HXY1、HXY2)的所有样本;Ⅱ组包含浙江省舟山市定海区蔡家岙(ZDC)居群的第1和第4份样本及浙江省金华市永康市方山口村(JYF)居群的所有样本;Ⅲ组包含安徽省黄山市祁门县牯牛降(HQG)居群的所有样本;Ⅳ组包含浙江省建德市寿昌林场(JSL)居群、浙江省杭州市淳安县向阳村(HCX)居群和浙江省宁波市宁海县归云洞(NNG)居群的所有样本;Ⅴ组包含浙江省舟山市定海区毛陈周(ZDM)居群的所有样本;Ⅵ组包含浙江省宁波市象山县屠家园村(NXT)居群和台湾省台北市北投区阳明山(TBY)居群的所有样本及ZDC居群的第2、第3、第5份样本。其中HCX、JSL、NNG、HQG、ZDC和TBY居群内样本间的亲缘关系较近,均能聚在一起;HLS1、HLS2居群间的个别样本有嵌套,但作为一个整体,这2个居群的样本也能聚在一起。ZDC居群5份样本中的3份与NXT和TBY居群的样本聚在一起,而ZDC居群的另2份样本与JYF居群的样本聚在一起,说明ZDC居群内遗传分化比较大。
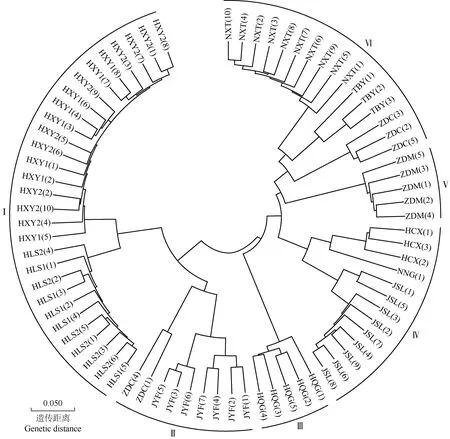
HLS: 浙江省杭州市临安区顺溪坞Shunxiwu, Lin’an District, Hangzhou City, Zhejiang Province; HXY: 浙江省杭州市西湖区云栖竹径Bamboo-lined path at Yunqi, Xihu District, Hangzhou City, Zhejiang Province; JSL: 浙江省建德市寿昌林场绿荷塘林区Lühetang Forest District, Shouchang Forest Farm, Jiande City, Zhejiang Province; HCX: 浙江省杭州市淳安县向阳村Xiangyang Village, Chun’an County, Hangzhou City, Zhejiang Province; HQG: 安徽省黄山市祁门县牯牛降Guniujiang, Qimen County, Huangshan City, Anhui Province; JYF: 浙江省金华市永康市方山口村Fangshankou Village, Yongkang City, Jinhua City, Zhejiang Province; NNG: 浙江省宁波市宁海县归云洞Guiyun Cave, Ninghai County, Ningbo City, Zhejiang Province; NXT: 浙江省宁波市象山县屠家园村Tujiayuan Village, Xiangshan County, Ningbo City, Zhejiang Province; ZDC: 浙江省舟山市定海区蔡家岙Caijia’ao, Dinghai District, Zhoushan City, Zhejiang Province; ZDM: 浙江省舟山市定海区毛陈周Maochenzhou, Dinghai District, Zhoushan City, Zhejiang Province; TBY: 台湾省台北市北投区阳明山Yangmingshan, Beitou District, Taipei City, Taiwan Province. 括号中数字为样本编号The numbers in brackets are sample numbers.图1 堇叶紫金牛13个居群77份样本的系统发育树Fig.. 1 Phylogenetic tree of 77 samples from 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao
2.4 主成分分析
基于SNP的主成分分析结果获得堇叶紫金牛13个居群77份样本的聚类情况,结果见图2。结果显示:根据PC1、PC2和PC3这3个主成分,可以将77份堇叶紫金牛样本分为6组,浙江省杭州市临安区顺溪坞2个居群(HLS1、HLS2)和浙江省杭州市西湖区云栖竹径2个居群(HXY1、HXY2)的所有样本为一组;安徽省黄山市祁门县牯牛降(HQG)居群的所有样本和浙江省舟山市定海区蔡家岙(ZDC)居群的2份样本为一组;浙江省金华市永康市方山口村(JYF)、浙江省建德市寿昌林场绿荷塘林区(JSL)、浙江省宁波市象山县屠家园村(NXT)居群的样本各单独为一组;浙江省宁波市宁海县归云洞(NNG)、浙江省杭州市淳安县向阳村(HCX)、浙江省舟山市定海区毛陈周(ZDM)和台湾省台北市北投区阳明山(TBY)居群的所有样本及ZDC居群的3份样本为一组。这与系统发育树分析结果相近,但也存在一定的差异。
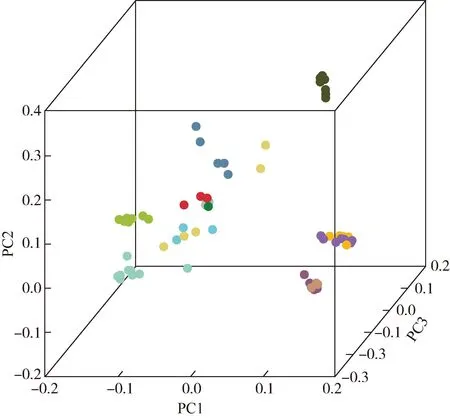
: HQG; : HCX; : HXY1; : HXY2; : JSL; : HLS1; : HLS2; : NNG; : TBY; : NXT; : JYF; : ZDC; : ZDM.HLS: 浙江省杭州市临安区顺溪坞Shunxiwu, Lin’an District, Hangzhou City, Zhejiang Province; HXY: 浙江省杭州市西湖区云栖竹径Bamboo-lined path at Yunqi, Xihu District, Hangzhou City, Zhejiang Province; JSL: 浙江省建德市寿昌林场绿荷塘林区Lühetang Forest District, Shouchang Forest Farm, Jiande City, Zhejiang Province; HCX: 浙江省杭州市淳安县向阳村Xiangyang Village, Chun’an County, Hangzhou City, Zhejiang Province; HQG: 安徽省黄山市祁门县牯牛降Guniujiang, Qimen County, Huangshan City, Anhui Province; JYF: 浙江省金华市永康市方山口村Fangshankou Village, Yongkang City, Jinhua City, Zhejiang Province; NNG: 浙江省宁波市宁海县归云洞Guiyun Cave, Ninghai County, Ningbo City, Zhejiang Province; NXT: 浙江省宁波市象山县屠家园村Tujiayuan Village, Xiangshan County, Ningbo City, Zhejiang Province; ZDC: 浙江省舟山市定海区蔡家岙Caijia’ao, Dinghai District, Zhoushan City, Zhejiang Province; ZDM: 浙江省舟山市定海区毛陈周Maochenzhou, Dinghai District, Zhoushan City, Zhejiang Province; TBY: 台湾省台北市北投区阳明山Yangmingshan, Beitou District, Taipei City, Taiwan Province.图2 基于SNP主成分分析结果的堇叶紫金牛13个居群77份样本的聚类Fig.. 2 Clustering of 77 samples from 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao based on SNP principal component analysis result
2.5 遗传结构分析
遗传结构分析结果(图3)显示:K=6时,交叉验证误差最小,且此时77份样本的遗传结构聚类清晰,表明堇叶紫金牛13个居群77份样本的最优分组数为6。
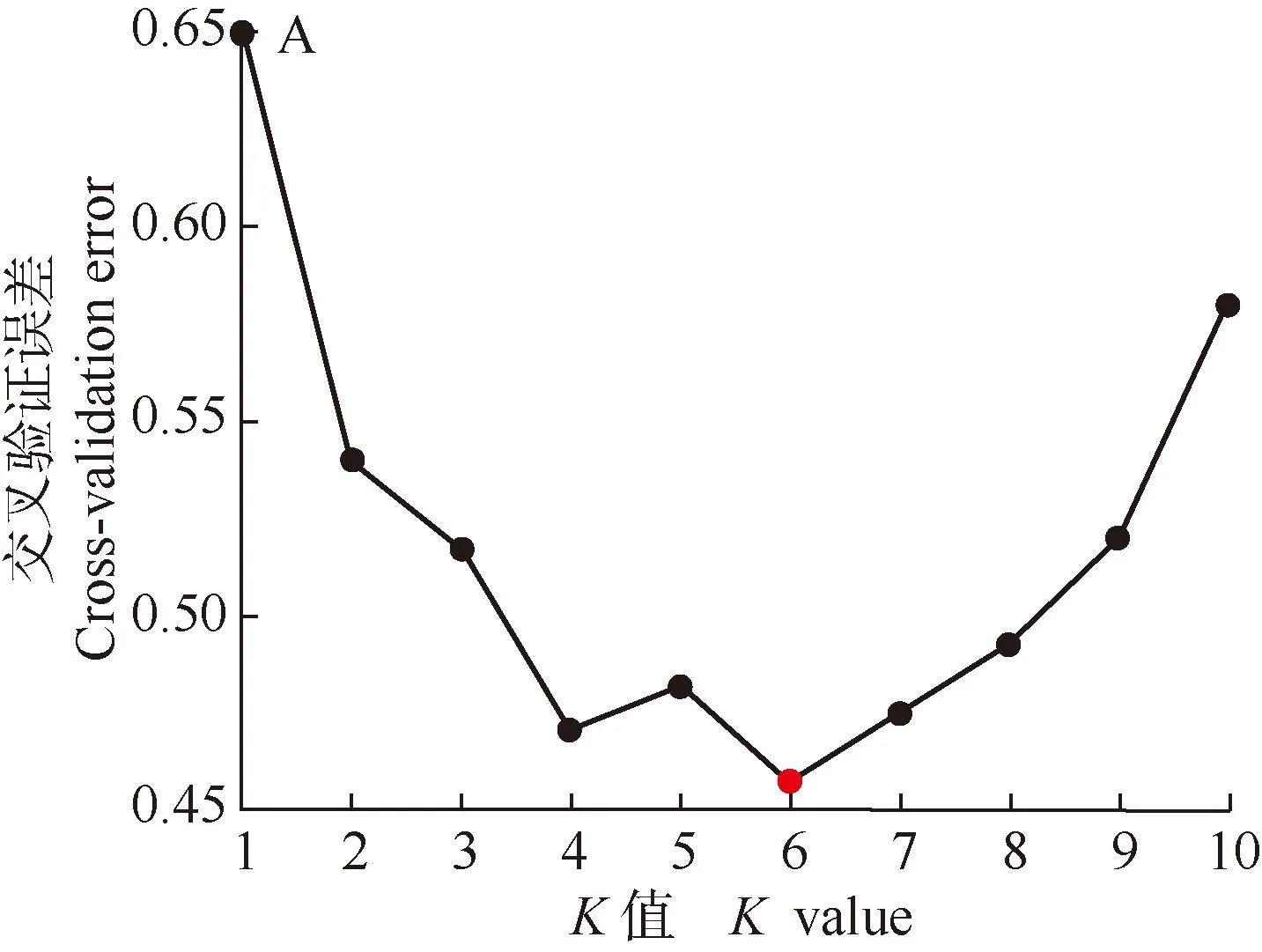
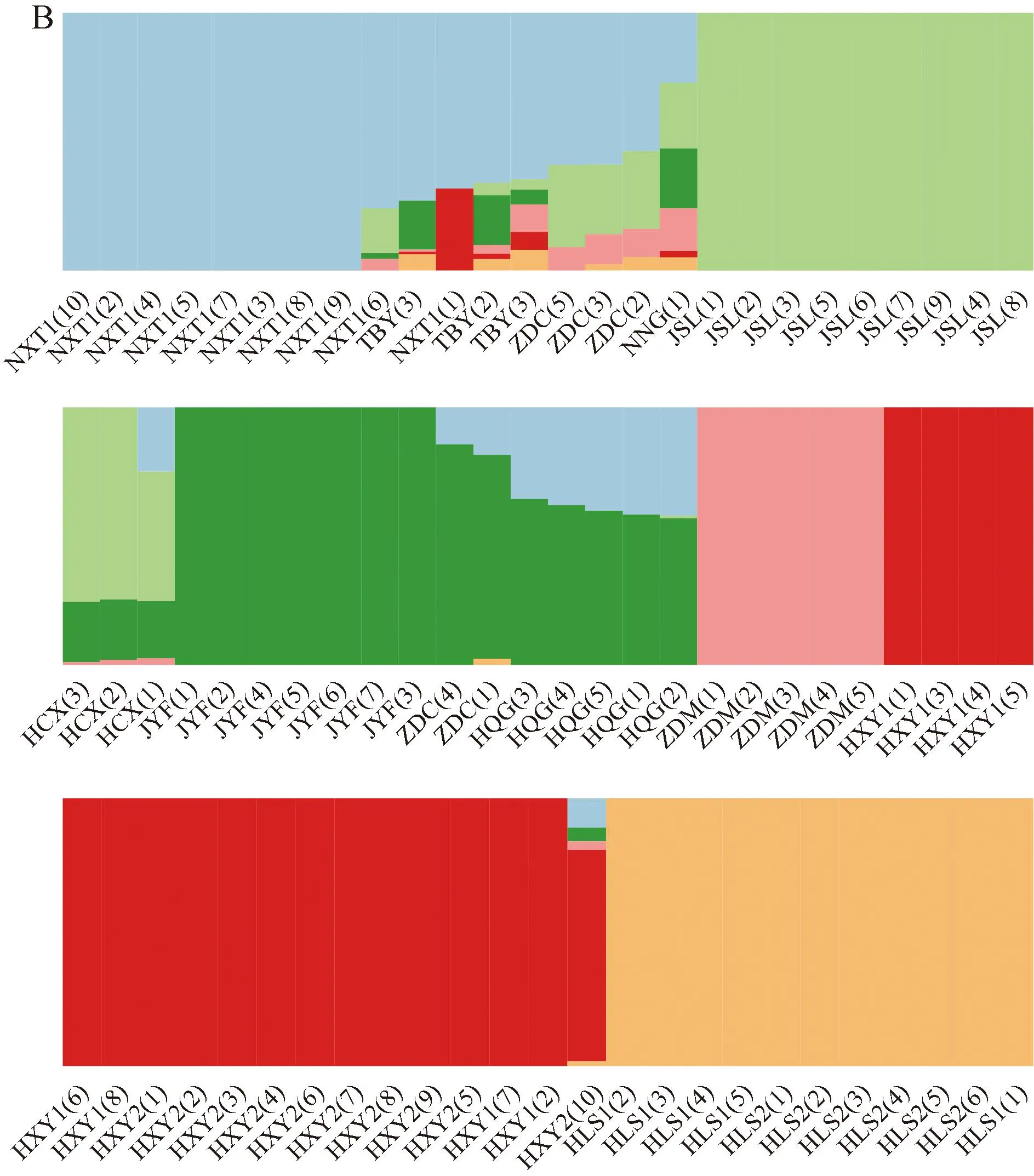
HLS: 浙江省杭州市临安区顺溪坞Shunxiwu, Lin’an District, Hangzhou City, Zhejiang Province; HXY: 浙江省杭州市西湖区云栖竹径Bamboo-lined path at Yunqi, Xihu District, Hangzhou City, Zhejiang Province; JSL: 浙江省建德市寿昌林场绿荷塘林区Lühetang Forest District, Shouchang Forest Farm, Jiande City, Zhejiang Province; HCX: 浙江省杭州市淳安县向阳村Xiangyang Village, Chun’an County, Hangzhou City, Zhejiang Province; HQG: 安徽省黄山市祁门县牯牛降Guniujiang, Qimen County, Huangshan City, Anhui Province; JYF: 浙江省金华市永康市方山口村Fangshankou Village, Yongkang City, Jinhua City, Zhejiang Province; NNG: 浙江省宁波市宁海县归云洞Guiyun Cave, Ninghai County, Ningbo City, Zhejiang Province; NXT: 浙江省宁波市象山县屠家园村Tujiayuan Village, Xiangshan County, Ningbo City, Zhejiang Province; ZDC: 浙江省舟山市定海区蔡家岙Caijia’ao, Dinghai District, Zhoushan City, Zhejiang Province; ZDM: 浙江省舟山市定海区毛陈周Maochenzhou, Dinghai District, Zhoushan City, Zhejiang Province; TBY: 台湾省台北市北投区阳明山Yangmingshan, Beitou District, Taipei City, Taiwan Province. 括号中数字为样本编号The numbers in brackets are sample numbers.图3 堇叶紫金牛13个居群77份样本的分组(A)及K=6时的遗传结构分析(B)Fig.. 3 Cluster(A) and genetic structure analysis when K=6 (B) of 77 samples from 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K.
2.6 亲缘关系分析
堇叶紫金牛77份样本间的亲缘关系分析结果见图4。颜色越红代表样本间的亲缘关系越近,同一居群内多个样本间热图呈红色说明这些样本可能构成了一个亲缘关系紧密的家系群体;反之,颜色越蓝说明样本间的亲缘关系越远。结果显示:堇叶紫金牛居群内个体间的亲缘关系较近。其中安徽省黄山市祁门县牯牛降(HQG)居群内样本间的亲缘关系最近,浙江省宁波市象山县屠家园村(NXT)、台湾省台北市北投区阳明山(TBY)、浙江省建德市寿昌林场绿荷塘林区(JSL)、浙江省舟山市定海区毛陈周(ZDM)居群内样本间的亲缘关系较近,而浙江省杭州市西湖区云栖竹径2个居群(HXY1、HXY2)和浙江省杭州市临安区顺溪坞2个居群(HLS1、HLS2)居群内样本间的亲缘关系较远。居群间的亲缘关系与地理距离有一定的相关性,HLS1、HLS2、HXY1、HXY2居群间的亲缘关系较近,但这4个居群与地理位置邻近的HQG、浙江省杭州市淳安县向阳村(HCX)、JSL居群的亲缘关系均较远。NXT、TBY和浙江省舟山市定海区蔡家岙(ZDC)居群间亲缘关系较近;HCX居群与TBY、JYF和ZDC居群间亲缘关系较远。
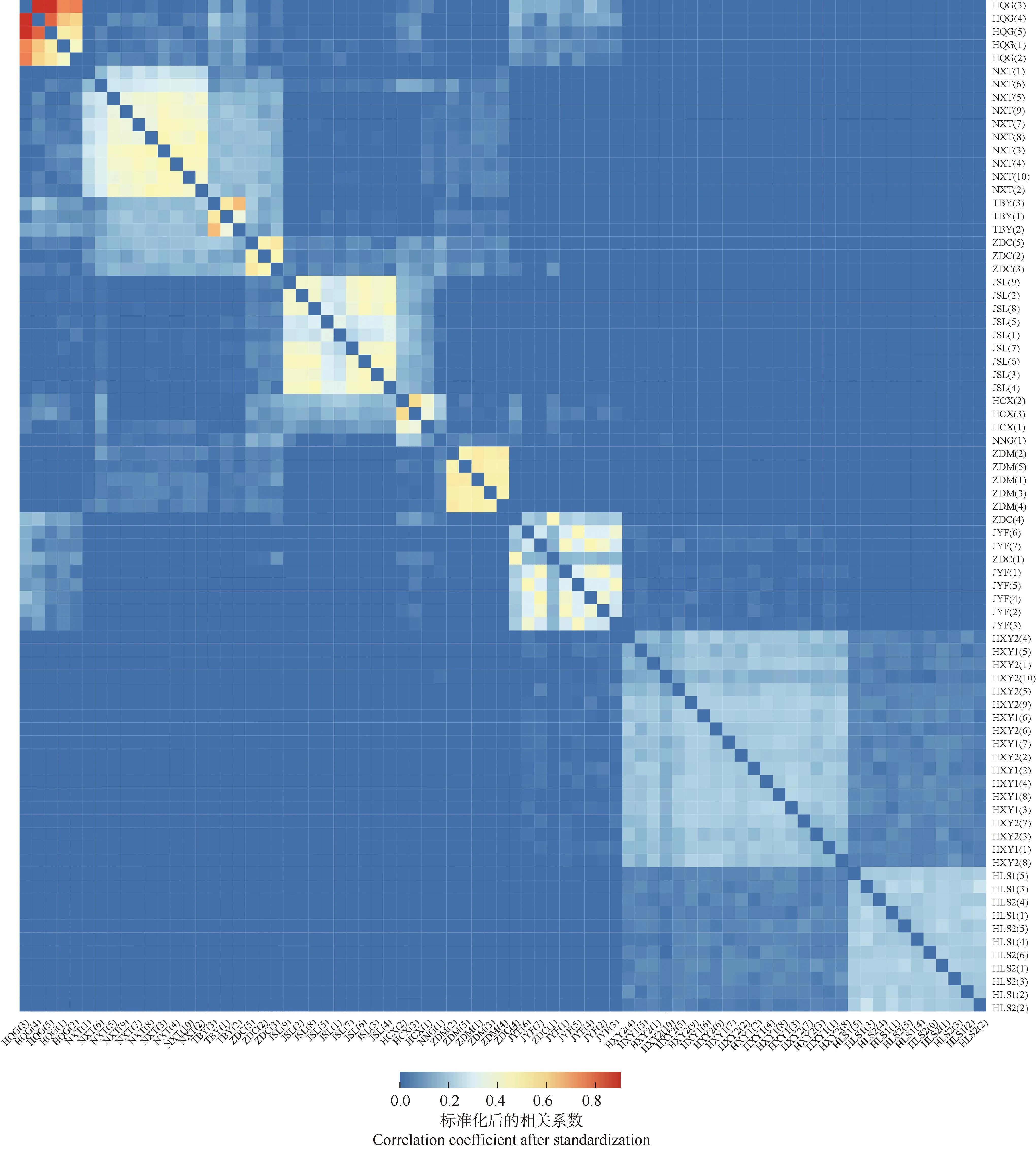
HLS: 浙江省杭州市临安区顺溪坞Shunxiwu, Lin’an District, Hangzhou City, Zhejiang Province; HXY: 浙江省杭州市西湖区云栖竹径Bamboo-lined path at Yunqi, Xihu District, Hangzhou City, Zhejiang Province; JSL: 浙江省建德市寿昌林场绿荷塘林区Lühetang Forest District, Shouchang Forest Farm, Jiande City, Zhejiang Province; HCX: 浙江省杭州市淳安县向阳村Xiangyang Village, Chun’an County, Hangzhou City, Zhejiang Province; HQG: 安徽省黄山市祁门县牯牛降Guniujiang, Qimen County, Huangshan City, Anhui Province; JYF: 浙江省金华市永康市方山口村Fangshankou Village, Yongkang City, Jinhua City, Zhejiang Province; NNG: 浙江省宁波市宁海县归云洞Guiyun Cave, Ninghai County, Ningbo City, Zhejiang Province; NXT: 浙江省宁波市象山县屠家园村Tujiayuan Village, Xiangshan County, Ningbo City, Zhejiang Province; ZDC: 浙江省舟山市定海区蔡家岙Caijia’ao, Dinghai District, Zhoushan City, Zhejiang Province; ZDM: 浙江省舟山市定海区毛陈周Maochenzhou, Dinghai District, Zhoushan City, Zhejiang Province; TBY: 台湾省台北市北投区阳明山Yangmingshan, Beitou District, Taipei City, Taiwan Province. 括号中数字为样本编号The numbers in brackets are sample numbers.图4 堇叶紫金牛13个居群77份样本间的亲缘关系热图Fig.. 4 Heat map of genetic relationship between 77 samples from 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao
2.7 遗传分化和基因流分析
结果(表3)显示:堇叶紫金牛13个居群间的遗传距离为0.059 0~0.393 8,遗传分化系数为0.000 0~0.423 0,且大部分(80.8%)居群间的遗传分化系数大于0.250 0,表明大部分居群间存在较高的遗传分化。浙江省杭州市西湖区云栖竹径2个居群(HXY1和HXY2)间及浙江省杭州市临安区顺溪坞2个居群(HLS1和HLS2)间的遗传分化系数较小,遗传距离较近,可能是由于这2个居群间的地理距离较小;而HXY1和HXY2居群与地理距离较远的台湾省台北市北投区阳明山(TBY)居群和安徽省黄山市祁门县牯牛降(HQG)居群间的遗传分化系数均较大(均大于0.400 0),遗传距离也较远。说明堇叶紫金牛居群间遗传分化系数与地理距离存在一定的相关性。
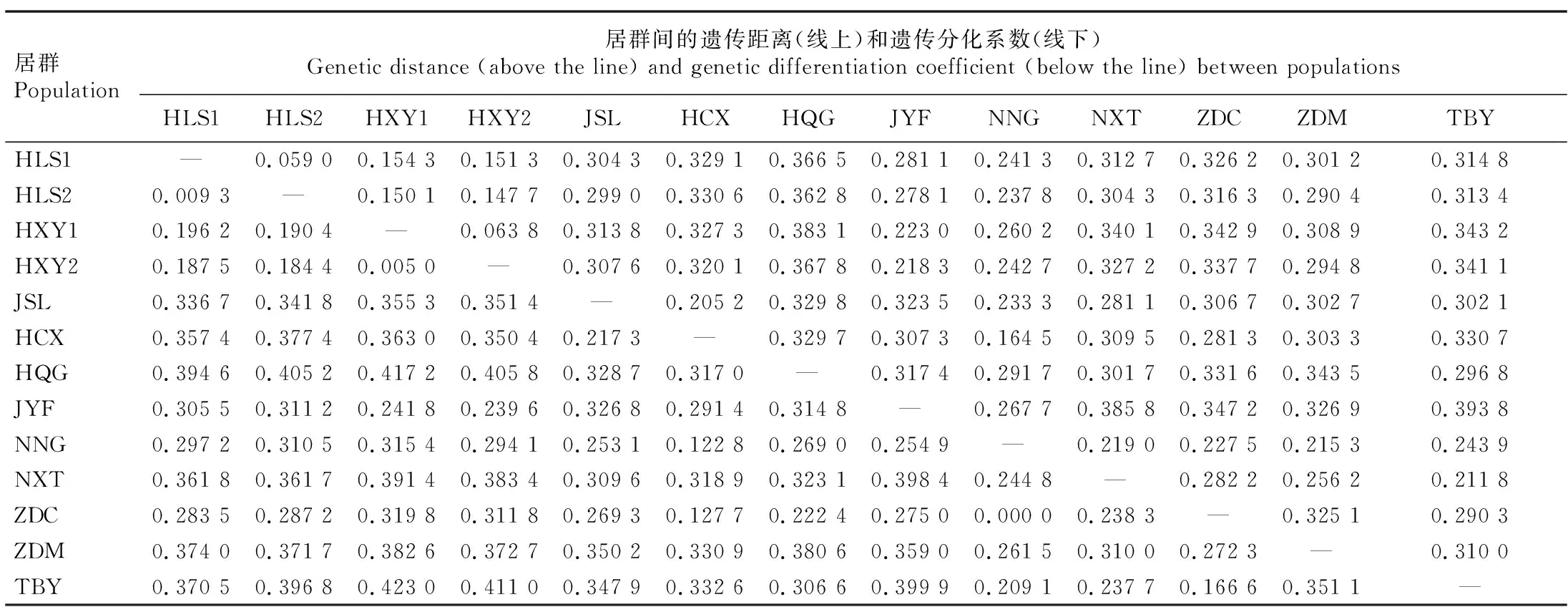
表3 堇叶紫金牛居群间的遗传距离和遗传分化系数1)Table 3 Genetic distance and genetic differentiation coefficient between populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao1)
基因流分析结果(图5)表明:浙江省宁波市象山县屠家园村(NXT)、浙江省舟山市定海区毛陈周(ZDM)和浙江省舟山市定海区蔡家岙(ZDC)居群聚为一支;HXY1、HXY2和HLS2等居群聚为另一支;TBY和HQG居群间及浙江省宁波市宁海县归云洞(NNG)和浙江省杭州市淳安县向阳村(HCX)居群间的遗传关系较近。各居群间存在一定程度的基因交流。在NNG居群分化过程中向HXY2居群产生了基因迁移事件;在NNG和HCX居群分化形成的过程中,NXT居群与二者存在基因交流,此基因迁移事件的权重最大;并且在NXT、HLS2和ZDC居群形成的过程中也均受到外界基因迁移事件的影响。
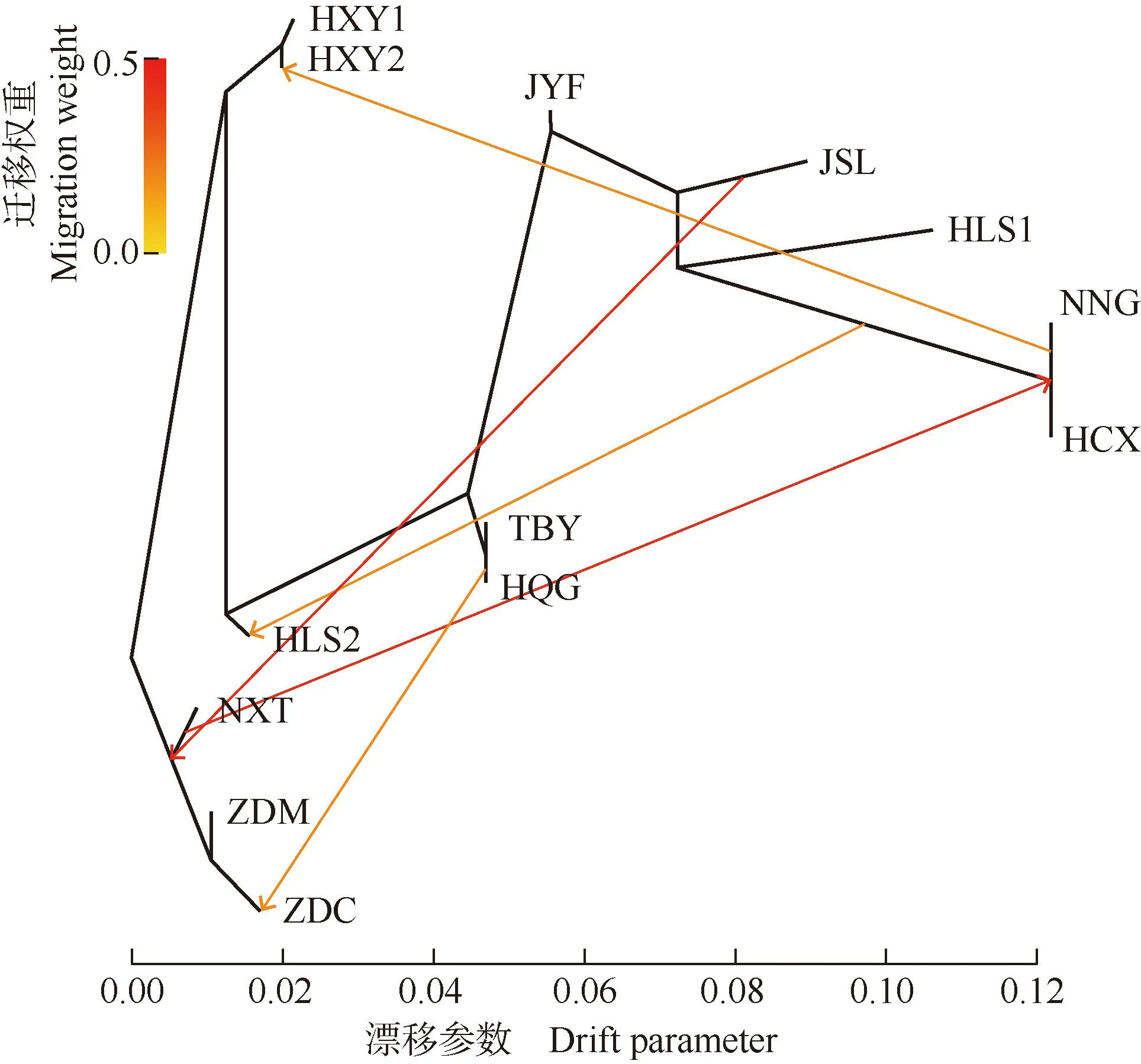
HLS: 浙江省杭州市临安区顺溪坞Shunxiwu, Lin’an District, Hangzhou City, Zhejiang Province; HXY: 浙江省杭州市西湖区云栖竹径Bamboo-lined path at Yunqi, Xihu District, Hangzhou City, Zhejiang Province; JSL: 浙江省建德市寿昌林场绿荷塘林区Lühetang Forest District, Shouchang Forest Farm, Jiande City, Zhejiang Province; HCX: 浙江省杭州市淳安县向阳村Xiangyang Village, Chun’an County, Hangzhou City, Zhejiang Province; HQG: 安徽省黄山市祁门县牯牛降Guniujiang, Qimen County, Huangshan City, Anhui Province; JYF: 浙江省金华市永康市方山口村Fangshankou Village, Yongkang City, Jinhua City, Zhejiang Province; NNG: 浙江省宁波市宁海县归云洞Guiyun Cave, Ninghai County, Ningbo City, Zhejiang Province; NXT: 浙江省宁波市象山县屠家园村Tujiayuan Village, Xiangshan County, Ningbo City, Zhejiang Province; ZDC: 浙江省舟山市定海区蔡家岙Caijia’ao, Dinghai District, Zhoushan City, Zhejiang Province; ZDM: 浙江省舟山市定海区毛陈周Maochenzhou, Dinghai District, Zhoushan City, Zhejiang Province; TBY: 台湾省台北市北投区阳明山Yangmingshan, Beitou District, Taipei City, Taiwan Province.图5 堇叶紫金牛13个居群间的基因流分析Fig.. 5 Analysis on gene flow among 13 populations of Ardisia violacea (T. Suzuki) W. Z. Fang et K. Yao
3 讨论和结论
影响物种遗传结构和遗传多样性的因子有很多,如生长环境[20]、分布区域[21]、种群数量以及繁殖系统特性[22]等,这些因子会影响物种对环境的适应能力[23]。本研究发现堇叶紫金牛的群体遗传多样性较低。蔡超男等[24]在研究濒危植物海南风吹楠(HorsfieldiahainanensisMerr.)时发现,由于自身更新能力弱及受过度人为活动的影响,海南风吹楠群体呈现出遗传多样性低的现状;桃儿七〔Sinopodophyllumhexandrum(Royle) Ying〕[25]、长序榆(UlmuselongataL. K. Fu et C. S. Ding)[26]、领春木(EupteleapleiospermaJ. D. Hooker et Thomson)[27]等濒危植物也有类似规律。而对伯乐树(BretschneiderasinensisHemsl.)的研究发现,伯乐树具有较丰富的遗传多样性[28],这可能与其分布范围较广、种群数量相对较多有关。相关研究表明:同属的濒危物种遗传多样性通常低于广域分布种,如普陀鹅耳枥(CarpinusputoensisCheng)遗传多样性低于疏花鹅耳枥〔C.laxiflora(Siebold et Zucc.) Blume〕[29-30],五小叶槭(AcerpentaphyllumDiels)遗传多样性低于青榨槭(A.davidiiFranch.)[31-32]。本研究中,堇叶紫金牛13个居群整体遗传多样性偏低,同属广域分布种朱砂根(ArdisiacrenataSims)遗传多样性相对较高[33-34]。由此推断遗传多样性低可能与濒危植物分布范围小、种群数量少有关,这可能是濒危植物的共有特征。
遗传分化系数是用于衡量居群间遗传分化程度的重要指标[35]。一般而言,当遗传分化系数大于0.25时,群体间遗传分化程度高;遗传分化系数小于0.05时,群体间遗传分化程度低[36]。本研究中,堇叶紫金牛居群间遗传分化系数多数大于0.25,说明堇叶紫金牛居群间遗传分化程度较高。濒危植物观光木〔Micheliaodora(Chun) Nooteboom et B. L. Chen〕和金铁锁(PsammosilenetunicoidesW. C. Wu et C. Y. Wu)的遗传分化系数(分别为0.375 2和0.452 9)[37-38]与堇叶紫金牛相近。濒危植物遗传分化程度通常高于广域分布种,如红砂〔Reaumuriasoongorica(Pall.) Maxim.〕和麻花秦艽(GentianastramineaMaxim.)的遗传分化系数(分别为0.088和0.114)[39-40]明显低于上述濒危植物,相对于广域分布种,濒危植物的生境片段化现象较为严重,居群间存在地理隔离,受异质化生境的影响而产生分化。
堇叶紫金牛系统发育树和主成分分析结果表明:整体表现为各居群内的样本聚在一起,这主要是因为堇叶紫金牛果实较大,主要通过重力传播,难以传播到较远的地方[5],因而居群间基因交流相对较少[41]。同时堇叶紫金牛也存在距离较远的居群间的基因交流,浙江省杭州市临安区顺溪坞2个居群(HLS1、HLS2)和浙江省杭州市西湖区云栖竹径2个居群(HXY1、HXY2)间的亲缘关系较近,但相较于地理距离较远的浙江省金华市永康市方山口村(JYF)和浙江省舟山市定海区蔡家岙(ZDC)居群,HLS1、HLS2、HXY1、HXY2居群与浙江省杭州市淳安县向阳村(HCX)、安徽省黄山市祁门县牯牛降(HQG)、浙江省建德市寿昌林场绿荷塘林区(JSL)居群间的地理距离更近,但亲缘关系却相对更远;此外,浙江省宁波市象山县屠家园村(NXT)和ZDC居群均与台湾省台北市北投区阳明山(TBY)居群存在较远的地理距离,但这3个居群间亲缘关系较近;HCX居群与TBY、JYF、ZDC居群有较近的亲缘关系,推测这可能是由鸟类取食并传播其种子所致。吸引鸟类传播的果实(种子)通常具有成熟时颜色鲜艳、体积较小、数量较多、果期长等特点[42-43],如圆叶相思树(AcaciacyclopsA. Cunn. ex G. Don)成熟时肉质假种皮橙红色[44],红豆杉〔Taxuschinensis(Pilger) Rehd.〕肉质假种皮红色[45],而红色、橙色等颜色对鸟类具有视觉吸引的效果[46],这些颜色的果实通常是鸟类取食的偏好对象。堇叶紫金牛果实颜色红艳,挂果时间10月至12月,符合鸟类秋冬季取食的偏好特征,因此可能存在鸟类传播扩散种子的现象。
堇叶紫金牛居群面积0.01~0.80 hm2,各居群间地理距离较远,生境片段化现象较为严重,加剧了居群内个体数量的减少,导致居群内近交几率增加,从而使遗传多样性降低。虽然堇叶紫金牛居群间存在较远距离的基因迁移,对居群间遗传多样性的提高具有一定作用,但对于堇叶紫金牛居群衰退的现状难以起到较大的影响。因此,针对堇叶紫金牛遗传多样性降低,应优先保护HQG、NXT和ZDC这3个遗传多样性较高的居群,建立优先保护单元;应制定相应的原地保存与迁地保存策略,减少人为干扰,保护现有野生群体;开展堇叶紫金牛相关的繁育工作,通过人工辅助的手段来维持并扩大居群数量,从而更好地保护其遗传多样性。
猜你喜欢
中小学校长(2022年12期)2022-12-30 12:50:46
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
动漫星空(2019年6期)2019-06-05 03:24:00
科学与技术(2019年8期)2019-04-10 12:27:48
草地学报(2018年5期)2018-11-07 02:25:00
经济研究导刊(2018年18期)2018-09-14 01:12:04
现代农业科技(2016年16期)2017-07-19 18:06:21
现代农业科技(2016年21期)2017-03-06 23:58:20
中国篆刻·书画教育(2016年3期)2016-03-29 16:46:41
