
海拔对祁连山东段青海云杉林林下灌草多样性的影响
2023-12-26 01:53:10李转桃徐先英罗永忠
植物资源与环境学报 2023年6期
李转桃, 徐先英, 赵 鹏, 罗永忠,①
(1. 甘肃农业大学林学院, 甘肃 兰州 730070; 2. 甘肃省治沙研究所, 甘肃 兰州 730071)
林下植被为森林生态系统的重要组成部分,可反映生态系统稳定性及群落演替规律,对于维持物种多样性、涵养水源、保持森林生态系统功能稳定等方面具有十分重要的作用[1-4]。一般而言,林下植被越丰富,森林生态系统稳定性越强,生态系统的结构和功能调节能力也越强,因此,丰富的林下植被有利于实现森林生态效益的提高及维持森林可持续经营。目前对于林下植被多样性的研究主要集中在海拔、坡向、土壤等环境因子[5-7]及林分密度、林龄、平均胸径等林分因子[8-10]上。不同研究区的森林物种组成和物种多样性因环境因子及林分因子变化而产生较大差异,因此,林下物种多样性指数也会有不同的变化趋势。
海拔作为综合性生态变量,集结了多种环境因子,包括土壤养分、光照条件和水热条件等;在森林生态系统中,海拔是影响林下物种组成和林下植被多样性的重要因子,随着海拔的变化,对植物群落影响最大的环境因子是温度,同时引起多种环境因子变化,致使局部微气候发生改变,进一步影响植物的生长,从而形成垂直海拔梯度上植物物种多样性的分布格局[11-12]。多项研究证实海拔对植物物种组成及物种多样性有显著影响,董雪等[13]以荒漠戈壁灌木群落为研究对象,分析其在海拔梯度上的分布规律,结果表明物种多样性随海拔升高呈先升后降的单峰分布格局;许涵等[14]研究表明除了人为森林采伐干扰外,海拔梯度是影响天然林物种多样性的最重要因子。可见,沿着海拔梯度的植物群落变化规律一直是生态学研究的重要内容。
不同海拔地区不同地段的植物群落多样性研究因所选样地的区域及区域内坡向、坡度、土壤等条件不同,所得研究结果不尽相同,植物多样性随海拔上升,有的逐渐增加,有的逐渐减少,有的先增加后减少,还有的无明显规律[15-17]。张旭等[18]对不同海拔森林样地的调查结果显示:随海拔升高,Marglef指数、Simpson优势度指数和Shannon-Wiener多样性指数在乔、灌层呈下降趋势,在草本层呈“U”型变化趋势;Pielou均匀度指数在乔木层呈下降趋势,在灌、草层变化不明显。王金兰等[19]对祁连山东段不同海拔梯度杜鹃(RhododendronsimsiiPlanch.)灌丛的调查发现,灌丛灌木高度、密度和生物量均随海拔升高呈先增加后降低的单峰变化趋势。
祁连山位于青藏高原、蒙新高原和黄土高原的交会带,其地理位置特殊、海拔高,自然环境独特、水系分布广、植被丰富多样[20]。青海云杉(PiceacrassifoliaKom.)是祁连山林区的主要建群树种之一,青海云杉林是祁连山自然保护区的主要森林类型,占祁连山乔木林总面积的66.4%。现有研究多集中在植物群落的物种组成、物种多样性以及生物量与海拔梯度之间的相关关系,但关于祁连山青海云杉林林下植被多样性随海拔梯度变化的研究较少。鉴于此,本研究以祁连山东段的青海云杉林为研究对象,通过调查不同海拔梯度青海云杉林林下植被组成,分析海拔对青海云杉林林下植被多样性的影响,以期掌握青海云杉林的群落特征,并为进一步精细化林下植被多样性保护、提高森林生态效益及森林可持续经营提供理论支持。
1 研究区概况和研究方法
1.1 研究区概况
祁连山区处于西北干旱半干旱区腹地,自然环境复杂,具有面积大、地形高度差较大等特点,且该区域气候差异较大。本研究区位于祁连山东段的哈溪-祁连-大黄山地区,地理坐标为东经102°15′42.9″~103°14′48.5″、北纬37°22′13.4″~38°25′52.9″,海拔范围为2 595~3 095 m。该研究区属高寒半干旱气候区,地势由西南向东北倾斜,年均温1.5 ℃,年均降水量约400 mm。研究区的主要植被种类有青海云杉、圆柏(JuniperuschinensisLinn.)、红桦(BetulaalbosinensisBurkill)等。
1.2 研究方法
1.2.1 样地设置及植被调查 从祁连山东段的甘沙河(海拔约2 595 m)开始设置样地,均设置在研究区内立地条件基本相似的山地上。海拔每升高100 m设置1个样地(林地),共5个林地,调查并记录每个林地的海拔、坡向、坡度和土壤类型及乔木的平均胸径和平均树高,具体情况见表1。每个林地设置3个面积20 m×20 m的乔木样方、5个面积5 m×5 m的灌木样方和5个面积1 m×1 m的草本样方,灌木样方和草本样方分别位于乔木样方的四角和中心。在2022年8月完成植被调查,调查指标包括鉴定乔木样方中的乔木种类、灌木样方中的灌木种类和草本样方中的草本种类,使用皮尺(精度0.1 cm)测量乔木胸径,使用测高仪(精度0.1和0.5 m)测量乔木和灌木的高度,使用卷尺(精度0.1 cm)测量灌木的冠幅(分别测量东西向冠幅和南北向冠幅,取平均值)及草本的高度和冠幅(分别测量东西向冠幅和南北向冠幅,取平均值),按照公式“灌木盖度=(东西向冠幅×南北向冠幅)/样地面积”计算灌木层盖度,采用目测法估测草本层盖度。
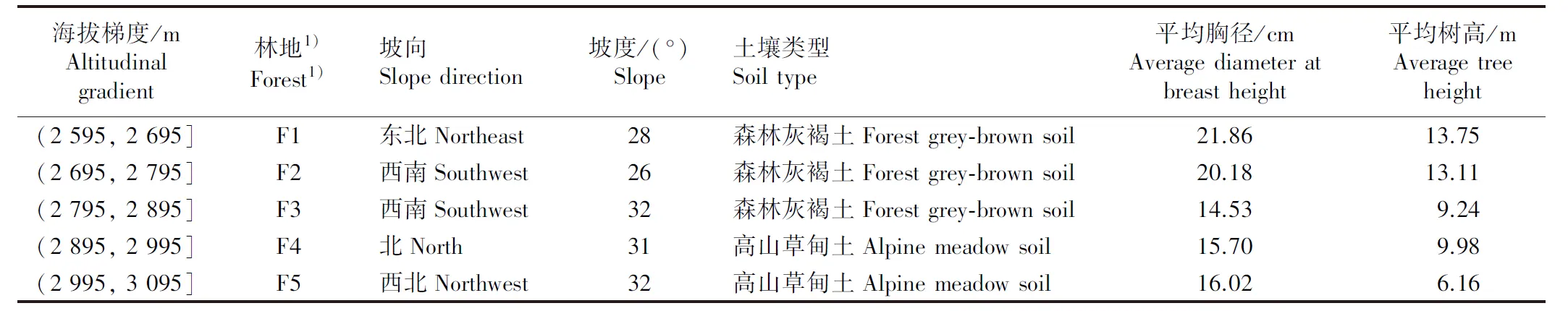
表1 祁连山东段青海云杉林各林地基本情况Table 1 Basic information of each forest of Picea crassifolia Kom. forests in the eastern Qilian Mountains
1.2.2 相关参数计算 参考卢训令等[21]的方法分别计算乔木层、灌木层和草本层植物的重要值,乔木植物重要值的计算公式为重要值=(相对多度+相对频度+相对显著度)/3;灌木植物和草本植物重要值的计算公式为重要值=(相对高度+相对频度+相对盖度)/3;同时参考卢训令等[21]的方法计算物种丰富度指数(S)、Simpson优势度指数(D)、Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)。通过Sørensen相似性系数分析不同海拔梯度林地间的相似性[22]。
1.3 数据处理
使用EXCEL 2019软件进行数据处理;使用IBM SPSS Statistics 26.0软件进行单因素方差分析(one-way ANOVA),分析不同海拔梯度林下灌草的多样性指数差异,并采用最小显著性差异(LSD)法进行显著性检验(P<0.05);使用IBM SPSS Statistics 26.0软件进行Pearson相关性分析。
2 结果和分析
2.1 不同海拔梯度青海云杉林林下灌草物种特征分析
2.1.1 物种组成 调查结果(表2)表明:祁连山东段青海云杉林林下植物共有79种,隶属于24科50属;林下灌木植物共有19种,隶属于7科9属,其中蔷薇科(Rosaceae)种类最多,有7种,其次为杜鹃花科(Ericaceae),有4种,这2科种数之和占调查的灌木植物总种数的57.9%;林下草本植物共有60种,隶属于21科42属,其中毛茛科(Ranunculaceae)种类最多,有8种,其次为菊科(Asteraceae)、豆科(Fabaceae)和蔷薇科,均有6种,这4科种数之和占调查的草本植物总种数的43.3%。
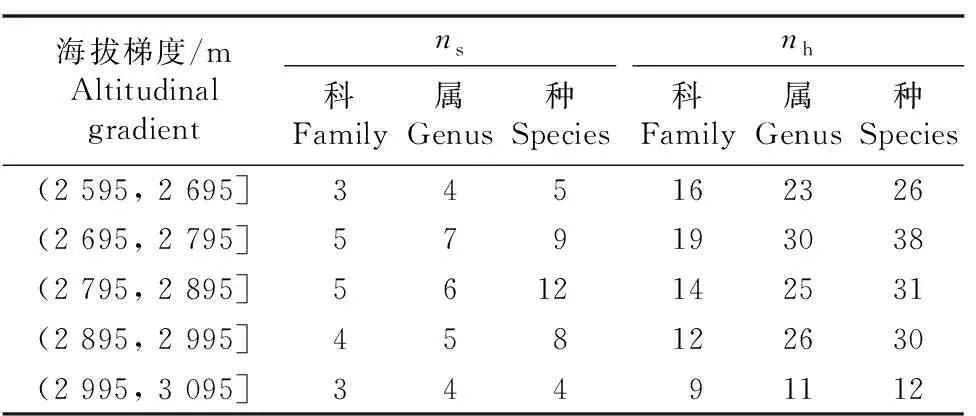
表2 祁连山东段不同海拔梯度青海云杉林林下灌草的物种组成1)Table 2 Species composition of shrubs and herbs under Picea crassifolia Kom. forests at different altitudinal gradients in the eastern Qilian Mountains1)
由表2还可见:祁连山东段不同海拔梯度林地灌木植物的科数量相差不大,海拔梯度 (2 695, 2 795] m的灌木植物属数量最多,海拔梯度 (2 795, 2 895] m的灌木植物种数最多,而海拔梯度(2 995, 3 095] m的灌木植物的科、属、种数量均最少;海拔梯度(2 695, 2 795] m的草本植物科、属、种数量均最多,而海拔梯度(2 995, 3 095] m的草本植物科、属、种数量均最少,且2个林地间数值相差较大。祁连山东段青海云杉林灌木和草本植物种数均随海拔升高呈现先增加后减少的趋势。
2.1.2 优势种分析 不同海拔梯度青海云杉林乔、灌、草层植物的重要值分析结果(表3)显示:群落建群种主要是青海云杉、圆柏、红桦、山杨(PopulusdavidianaDode)、秦柳(SalixchingianaHao ex Fang et Skvortsov),并且随海拔梯度变化发生更替。5个海拔梯度林地灌木层的优势种由海拔从低到高分别为银露梅〔Dasiphoraglabra(G. Lodd.) Sojk〕、小叶金露梅〔D.parvifolia(Fisch. ex Lehm.) Juz.〕、山生柳(SalixoritrephaSchneid.)、银露梅、金露梅〔D.fruticosa(Linn.) Rydb.〕;5个海拔梯度林地草本层的优势种主要为薹草(Carexspp.)和珠芽蓼〔Bistortavivipara(Linn.) Gray〕。
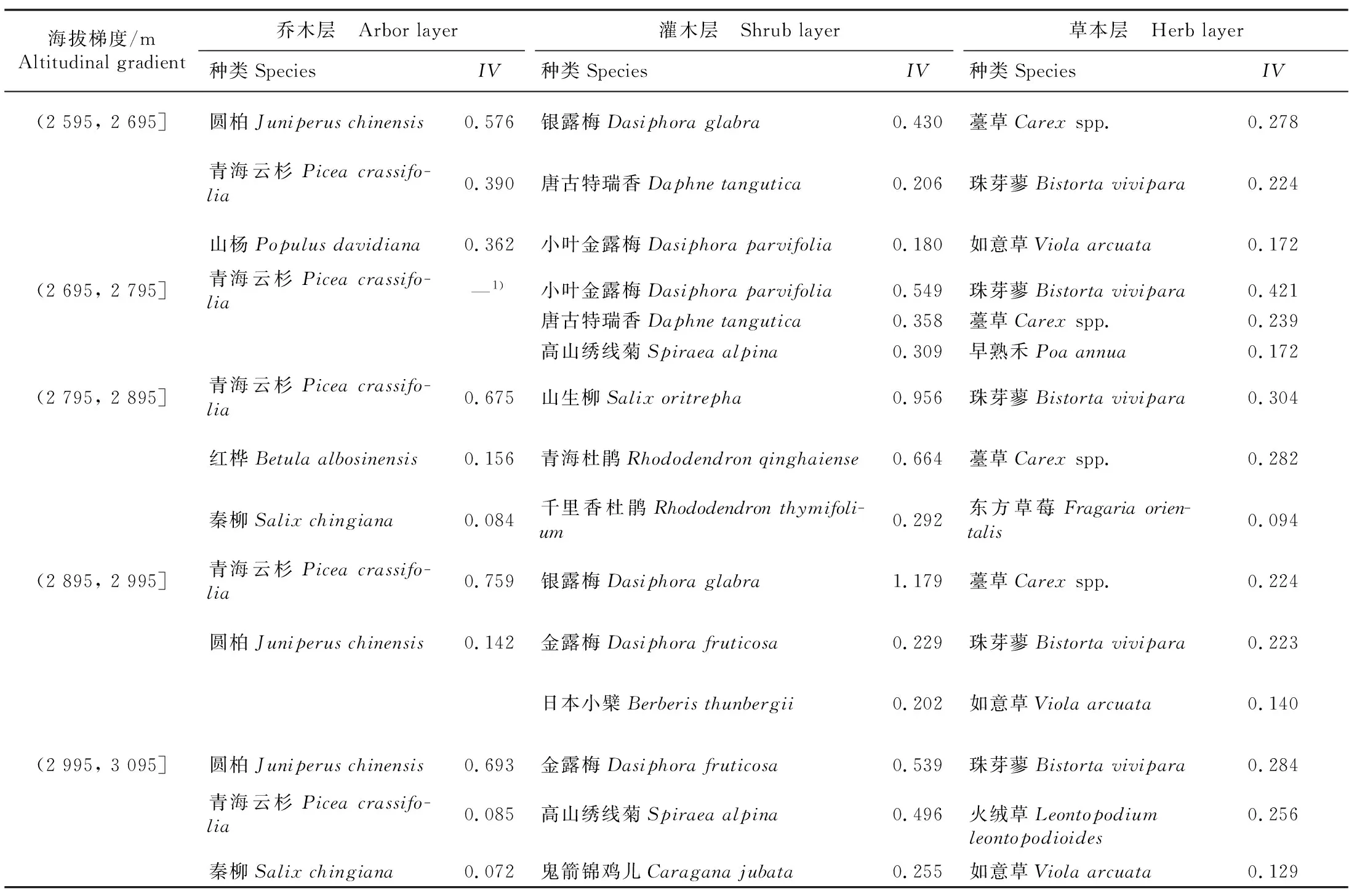
表3 祁连山东段不同海拔梯度青海云杉林乔、灌、草层植物的重要值(IV)Table 3 Importance values (IV) of species of arbor, shrub, and herb layers under Picea crassifolia Kom. forests at different altitudinal gradients in the eastern Qilian Mountains
2.2 不同海拔梯度青海云杉林林下灌草植物多样性分析
2.2.1 林下灌草植物多样性特征分析 对不同海拔梯度下青海云杉林林下灌草层的物种丰富度指数(S)、Simpson优势度指数(D)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)进行分析,结果见表4。结果显示:同一海拔梯度林地草本层的S值、D值和H′值均高于灌木层,而J值均低于灌木层。灌木层的S值、D值、H′值整体随海拔升高呈先升高后降低的趋势,且海拔梯度(2 795, 2 895] m的S值显著(P<0.05)高于海拔梯度(2 595, 2 695]、(2 695, 2 795]和(2 995, 3 095] m,海拔梯度(2 795, 2 895] m的D值极显著(P<0.01)高于海拔梯度(2 595, 2 695]、(2 695, 2 795]和(2 995, 3 095] m,海拔梯度(2 795, 2 895] m的H′值极显著高于海拔梯度(2 595, 2 695] m;J值在5个海拔梯度林地间无显著(P>0.05)差异。草本层的D值随海拔升高呈先增高后降低的趋势,且海拔梯度(2 795, 2 895] m的D值分别极显著和显著高于海拔梯度(2 595, 2 695]和(2 995, 3 095] m;5个海拔梯度林地中,海拔梯度(2 695, 2 795] m的S值和H′值最高,海拔梯度(2 795, 2 895] m的J值最高。

表4 祁连山东段不同海拔梯度下青海云杉林林下灌草植物多样性分析1)Table 4 Diversity analysis of shrubs and herbs under Picea crassifolia Kom. forests at different altitudinal gradients in the eastern Qilian Mountains1)
2.2.2 林下灌草植物多样性指数及海拔间的相关关系分析林下灌木植物多样性指数及海拔间的相关性分析结果(表5)显示:灌木层的D值、H′值、J值与海拔呈负相关,S值与海拔呈正相关,且S值和J值与海拔间的相关性达显著水平;林下灌木层的各多样性指数间无显著相关性。
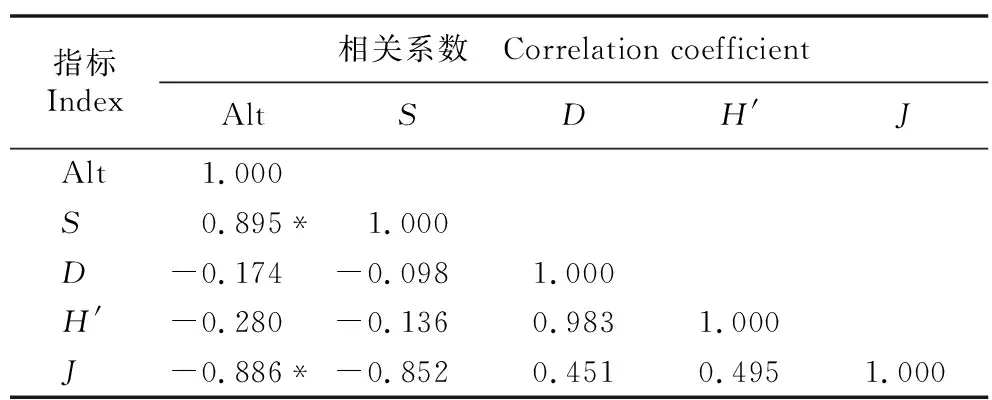
表5 祁连山东段青海云杉林林下灌木植物多样性指数及海拔间的相关性分析1)Table 5 Correlation analysis of diversity indexes of shrubs under Picea crassifolia Kom. forest and altitude in the eastern Qilian Mountains1)
林下草本植物多样性指数及海拔间的相关性分析结果(表6)显示:草本层的S值与海拔呈负相关,D值、H′值、J值与海拔呈正相关,但均未达显著水平;S值与D值、H′值呈显著负相关,H′值与J值呈显著正相关。
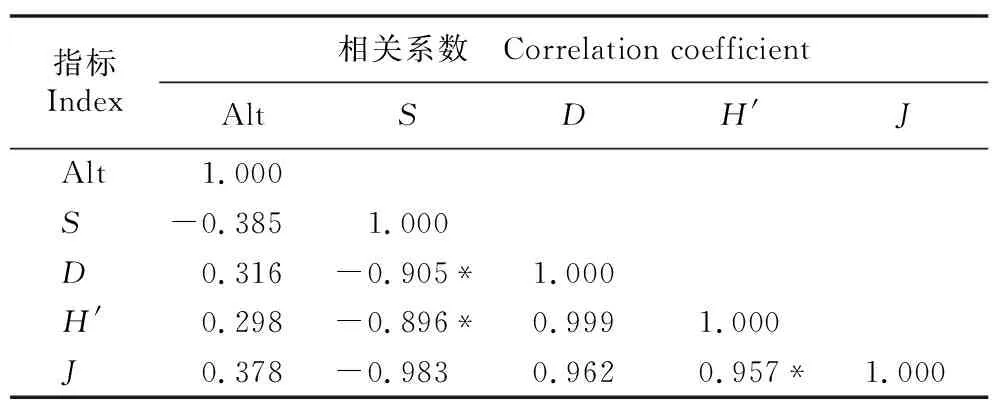
表6 祁连山东段青海云杉林林下草本植物多样性指数及海拔间的相关性分析1)Table 6 Correlation analysis of diversity indexes of herbs under Picea crassifolia Kom. forest and altitude in the eastern Qilian Mountains1)
2.3 不同海拔梯度青海云杉林间的Sørensen相似性系数分析
通过调查不同海拔梯度青海云杉林林下共有或各自拥有的灌草植物,得到不同青海云杉林林下灌草植物的Sørensen相似性系数,结果见表7。海拔梯度(2 695, 2 795] m与(2 595, 2 695] m林地灌草植物合计64种,共有植物21种,相似程度最高,Sørensen相似性系数达0.615;海拔梯度(2 795, 2 895] m与(2 695, 2 795]、(2 595, 2 695] m林地间的Sørensen相似性系数较高,分别为0.588和0.580;海拔梯度(2 995, 3 095] m与其他4个海拔梯度林地间的相似性均较低,尤其是与海拔梯度(2 595, 2 695] m林地间,仅有5种共有植物,Sørensen相似性系数仅为0.213。随海拔升高,相邻海拔梯度林地间的相似性逐渐降低,以海拔梯度(2 595, 2 695] m与(2 695, 2 795] m间的相似性最高,海拔梯度(2 895, 2 995] m与(2 995, 3 095] m间的相似性最低。
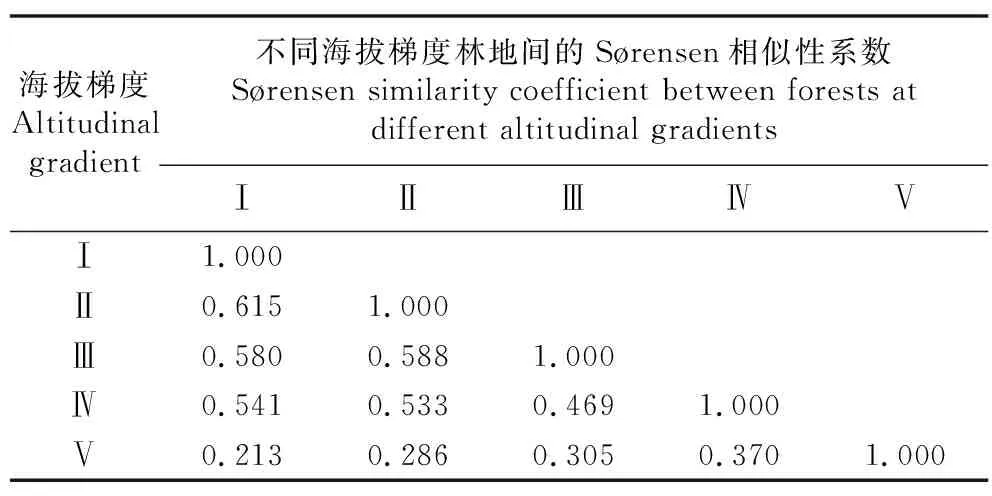
表7 祁连山东段不同海拔梯度青海云杉林间的Sørensen相似性系数1)Table 7 Sørensen similarity coefficients between Picea crassifolia Kom. forests at different altitudinal gradients in the eastern Qilian Mountains1)
3 讨论和结论
3.1 海拔对青海云杉林林下灌草植物组成的影响
物种组成结构是植物群落的重要特征,随海拔变化,林下植被的物种组成、分布结构等均会因海拔形成的各种变量因子产生明显的区别[23-25]。张入匀等[26]的研究结果表明:物种多样性的分布格局与环境条件紧密相关,不同区域沿海拔梯度的物种多样性变化趋势存在一定差异;冯哲等[27]通过研究木本植物幼苗的组成及数量随海拔的变化发现,不同海拔梯度的物种组成及群落结构明显不同;陈水飞等[28]对武夷山国家公园不同海拔梯度植物群落的物种组成研究表明:不同类型植物群落随海拔升高,样地内植物科、属、种的数量总体呈减少的趋势。本研究中,蔷薇科植物种数在5个海拔梯度林地灌木层中均最多,是每个林地的重要组成部分,说明蔷薇科植物在5个海拔梯度都具有较强的适应性;其中,除海拔梯度(2 995, 3 095] m外,蔷薇科的小叶金露梅在各林地类型中均有分布,并在海拔梯度(2 695, 2 795] m属于灌木层的优势种,说明小叶金露梅对高海拔的适应能力较强,是本调查区域的适生种和自然生长最广的灌木树种。5个海拔梯度林地的草本层植物较灌木层丰富,其中毛茛科的草本植物种类最多,菊科、豆科和蔷薇科次之,但这4科种数之和在草本植物总种数中占比不足一半。因生境条件不同,蝶形花科(Papilionaceae)、杜鹃花科、虎耳草科(Saxifragaceae)、苦苣苔科(Gesneriaceae)、木贼科(Equisetaceae)、茜草科(Rubiaceae)、瑞香科(Thymelaeaceae)、荨麻科(Urticaceae)、罂粟科(Papaveraceae)、鸢尾科(Iridaceae)均只有1种,表明群落调查中的草本植物基本为伴生种或偶见种,物种分布随机,没有形成明显的竞争关系。蓼科(Polygonaceae)有掌叶大黄(RheumpalmatumLinn.)、珠芽蓼、线叶萹蓄(PolygonumparonychioidesC. A. Mey. ex Hohen.)3种,其中珠芽蓼在5个海拔梯度林地中皆有分布且重要值均较大;此外,莎草科(Cyperaceae)的薹草在4个海拔梯度林地中均出现且重要值均较大;因此珠芽蓼和薹草为5个海拔梯度林地草本植物的优势种。本研究还发现,不管是低海拔的矮化圆柏-青海云杉混交林,还是高海拔的青海云杉-圆柏混交林和圆柏-青海云杉矮林,堇菜科(Violaceae)的如意草(ViolaarcuataBlume)均占据一席之地,表明如意草可以适应不同海拔梯度的林下生态环境,成为该研究区域不同海拔梯度下的重要组成物种。
3.2 海拔对青海云杉林林下灌草植物多样性的影响
由于调查样地的总体海拔范围不够大,因此没有形成直观的植被垂直带谱,在不同海拔梯度上植被的结构和组成差异也较大,其中,个体多度和所选样地的生境条件是差异形成的重要因子[29],因此不同林地类型的灌木层和草本层植物多样性指数存在一定差异[30]。杨壹等[31]通过对贺兰山不同海拔典型植被的多样性研究表明:不同海拔区域植物多样性总体存在显著(P<0.05)差异且具有明显的垂直分布格局。另有多项研究表明:灌木层和草本层的物种丰富度指数、Pielou均匀度指数、Simpson优势度指数和Shannon-Wiener多样性指数随着海拔增加呈单峰变化趋势,符合物种多样性垂直分布的“中间高度膨胀”理论[32-34]。本研究灌木层的物种丰富度指数、Simpson优势度指数和Shannon-Wiener多样性指数及草本层的Simpson优势度指数对海拔的响应结果也与之一致,呈现先升高后降低的变化趋势。本研究中物种多样性在草本层间的差异较小,而在灌木层间的差异显著,总体上灌木层的多样性指数低于草本层,且不同林地灌草层之间的多样性指数差异较明显;灌木层物种多样性受海拔影响较大,草本层物种多样性则受影响较小,这可能是由于海拔升高会导致温度降低,不利于灌木植物的生长发育,而草本层种类多为耐寒植物,适应能力强,因此其多样性指数变化较小,这与杨志汝等[35]对秦岭太白山巴山冷杉(AbiesfargesiiFranch.)天然林群落物种多样性的研究结果一致。草本层植物物种多样性表现为海拔梯度(2 695, 2 795] m的物种丰富度指数和Shannon-Wiener多样性指数高于其他海拔梯度,海拔梯度(2 795, 2 895] m的Simpson优势度指数和Pielou均匀度指数高于其他海拔梯度,且Simpson优势度指数随海拔升高呈先升高后降低的趋势;不同海拔梯度灌木层的Pielou均匀度指数没有显著差异,但草本层差异较大,由于乔木树种及一些高大的灌木形成的林冠层较高,灌木层植物可获得足够的生存资源(如降水、光照等),故其Pielou均匀度指数差异不大,但林冠对林下草本植物产生遮蔽作用,林内光照强度减弱且林冠会截留部分降水,因此草本植物间存在种间竞争,草本层植物多样性会相应降低,导致草本层在不同海拔梯度的Pielou均匀度指数产生差异;另一方面,不同草本层植物种类对生存空间有不同的需求,如光照、水分、土壤等,生存空间不同导致Pielou均匀度指数差异较大。本研究中,不同海拔梯度林地间Sørensen相似性系数随海拔升高逐渐降低,原因是随海拔升高,林地生境会发生很大改变,青海云杉林林下灌草植物对生境条件更加敏感,物种的稳定性、抗干扰能力降低,从而导致各林地的植物种类较容易发生变化。
3.3 结论
祁连山东段青海云杉林林下灌草植物物种组成丰富,共有24科50属79种,灌木层物种多样性受海拔影响显著,草本层物种多样性受海拔影响不明显,灌木层的物种丰富度指数、Simpson优势度指数和Shannon-Wiener多样性指数整体随海拔升高呈单峰变化趋势,符合物种多样性垂直分布的“中间高度膨胀”理论。随着海拔升高,相邻海拔梯度林地间的相似性逐渐降低,以海拔梯度(2 595, 2 695] m与(2 695, 2 795] m林地间的相似性最高,海拔梯度(2 895, 2 995] m与(2 995, 3 095] m林地间的相似性最低。海拔梯度作为综合性生态变量,集结了很多不同的环境因子(如土壤腐殖质含量、水分条件、温度等),因此,在今后的工作中可在环境因子方面进行深入研究。
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
少儿美术(2020年8期)2020-11-06 05:21:38
中国水土保持科学(2019年6期)2019-04-26 05:13:48
乡村科技(2018年8期)2018-02-13 12:16:18
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
浙江农林大学学报(2016年3期)2016-06-30 03:08:44
甘肃林业(2016年6期)2016-02-27 11:29:12
西部林业科学(2015年6期)2016-01-27 10:39:16
