
黔中山区5个糯玉米自交系主要农艺性状的配合力分析
2023-12-13 15:25赵艳花唐谷任明刚潘中涛汪朝明
南方农业·上旬 2023年9期
赵艳花 唐谷 任明刚 潘中涛 汪朝明



摘 要 为给黔中山区新育成自交系的有效利用提供理论依据,以5个糯玉米自交系(AN-16、AN-61、AN-65、AN-2、AN-76)为材料,采用GriffingI完全双列杂交法组配25个杂交组合,分析其一般配合力(GCA)效应和特殊配合力(SCA)效应,估计遗传参数。结果表明,在不同性状间,5个自交系的一般配合力和特殊配合力存在明显差异。产量、行粒数、百粒重、穗行数GCA效应均以AN-61最高;AN-2对降低F1果穗秃尖长的GCA效应最大。用AN-61作亲本之一,对提高其F1产量,减小秃尖具有较高利用价值;用AN-2作亲本之一,对提高F1产量,减小秃尖具有正效应;利用AN-65、AN-16作亲本之一,可组配出植株矮、穗位低的杂交组合,但对提高其产量性状还需要开展重点研究。
关键词 糯玉米;自交系;农艺性状;一般配合力;特殊配合力;遗传参数
中图分类号:S513 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2023.17.011
糯玉米(Zea mays L. sinensis Kulesh)是硬质玉米突变形成的一种特殊玉米类型,因具有较高的直链淀粉含量[1],成为了蔬菜市场比较受青睐的优质玉米品种。
自交系选育是糯玉米育种的基础和难点,组配杂交组合是重点,配合力测定是自交系选育和杂交种组配的关键[2-3]。主要农艺性状配合力的强弱是判断糯玉米自交系利用价值的主要评价指标,随着各研究单位在特用玉米研究内容的不断深入,对糯玉米自交系配合力的研究也逐步深入,并培育出一批新优糯玉米品种[4-9]。近年来,贵州省大力发展蔬菜产业,作为特色蔬菜品种的鲜食糯玉米,产业规模已经达到4.67万hm2[10]。贵州山区鲜食糯玉米产业迅猛的发展势头对新优品种培育提出了更高要求,因而,加快选育和高效利用优良糯玉米自交系具有十分重要的意义。安顺市农业科学院围绕黔中山区大坝蔬菜产业发展需求,大力开展糯玉米新品种培育,经过多年选育,培育出了一批糯玉米自交系。为有效探索新育成糯玉米自交系在生产上的应用价值,本研究采用GriffingI完全双列杂交法设计,对5个新选育的糯玉米自交系的主要农艺性状及配合力进行比较分析,以期明确这些自交系配合力高低,找出遗传差异,对其利用前景进行评价,为进一步合理利用种质资源,组配出优良糯玉米杂交种提供理论参考依据。
1 材料与方法
1.1 试验材料
2019年,以AN-16(P1)、AN-61(P2)、AN-65(P3)、AN-2(P4)和AN-76(P5)等5个糯玉米自交系为材料(均为安顺市农业科学院新选育出的二环系),采用GriffingI完全双列杂交法组配25个杂交组合(含5个自交材料)。
1.2 试验方法
2020年,在安顺市农科院玉米試验地实施田间测试。试验采用随机区组设计,3次重复,3行区,行长3.5 m、行距0.8 m、株距0.26 m,密度为3 167株/667 m2。田间调查记载株高、穗位高、雄穗主轴长、雄穗一级分支数等4个性状,成熟后全部收获计产,取样室内风干后考察穗长、穗粗、秃尖长、穗行数、行粒数、百粒重和产量等7个性状,并进行配合力分析。试验数据利用EXCEL 2010进行归纳整理,利用DPS 7.05软件进行统计分析。
2 结果与分析
2.1 不同组合的11个农艺性状方差分析
根据单因素随机区组设计方差分析方法,逐一对25个组合的11个农艺性状表现进行方差分析,结果见表1。结果显示,25个组合的11个性状在重复间差异不显著,组合间差异达极显著水平,由此说明,25个组合间遗传差异真实存在,可以进一步分析一般配合力和特殊配合力,估计遗传方差组成成分。
2.2 不同组合的最主要农艺性状配合力方差分析
采用固定模型对11个性状的GCA和SCA进行方差分析,结果(见表2)显示,11个观测性状的GCA均达极显著水平,说明亲本的GCA对F1株高、穗位高等11性状的效应存在极显著差异。正交SCA除秃尖长差异显著外,其余10个性状均达到极显著水平,说明亲本正交SCA对F1除秃尖长外的10性状的效应存在极显著差异,对F1秃尖长的效应存在显著差异。穗行数、秃尖长反交效应差异不显著,百粒重反交效应差异显著,其余8个性状反交效应差异极显著。5个亲本的11个性状GCA、SCA效应分析结果显示,不同亲本在不同性状间的GCA、SCA效应各异。
2.3 一般配合力效应(GCA)分析
2.3.1 植株性状
由表3可知,5个亲本中,AN-61、AN-2、AN-76具有增加株高的正效应,以AN-61效应值最大;AN-65和AN-16具有降低株高的正效应,以AN-65的效应值最大。AN-61和AN-2具有增加穗位高的正效应,以AN-2效应值最大;AN-76、AN-65和AN-16具有降低穗位高的正效应,以AN-16的效应值最大。除AN-61的雄穗主轴长与分支数均为正效应外,其余4个亲本的雄穗主轴长与分支数都显示为一正一负,其中,AN-76和AN-65的雄穗主轴长一般配合力效应值显示为正效应,AN-16和AN-2的雄穗分支数一般配合力效应值显示为正效应。
2.3.2 穗部性状
从5个穗部性状的GCA效应值表(见表3)可以看出,在5个自交系中,AN-61穗长、穗粗、穗行数、行粒数的GCA为正效应,对秃尖长减小的GCA显示正效应;AN-2穗粗、穗行数、行粒数和减小秃尖长的GCA为正效应,穗长的GCA为负效应;AN-76穗长、行粒数的GCA为正效应,穗粗、穗行数和增加秃尖长的GCA为负效应;AN-65穗长、穗粗、穗行数、行粒数的GCA为负效应,对秃尖长增加的GCA为正效应;AN-16穗粗、穗行数和秃尖长减小的GCA为正效应,穗长、行粒数的GCA为负效应。
2.3.3 产量性状
从表3可以看出,5个自交系中,除AN-16的百粒重一般配合力为负效应外,其余4个自交系百粒重的一般配合力均为正效应,以AN-61的效应值最大;只有AN-61和AN-2的产量一般配合力为正效应,其余3个自交系产量一般配合力均为负效应。
2.4 特殊配合力(SCA)效应分析
2.4.1 正交组合特殊配合力效应分析
特殊配合力是指某些特定的组合在其双亲平均表现的基础上与预期结果的偏差,是两个亲本通过互作而表现出的非加性效应,是不能遗传的部分,受环境因素影响大。正交组合SCA效应分析结果(见表4)显示,同一测定性状不同正交组合SCA效应值存在不同差异。
植株性状:10个正交组合株高SCA效应值变幅为 -5.46~21.31,正效应的组合9个,效应值以AN-16×AN-2最大,AN-16×AN-61其次;AN-16×AN-65为负效应。穗位高SCA效应值变幅为-5.00~15.77,6个正效应组合,效应值以AN-16×AN-76最高,AN-16×AN-61其次;4个负效应组合,负效应最大的组合是AN-2×AN-76。雄穗主轴长SCA效应值变幅为-1.58~2.79,正效应值组合5个,效应值以AN-61×AN-76最大;负效应值组合5个,负效应最大的组合是AN-61×AN-2。雄穗分支数SCA效应值变幅为-0.73~1.95,正效应组合8个,效应值最大的是AN-16×AN-76;负效应组合2个,负效应最大的是AN-61×AN-65。
穗部性状:10个正交组合穗长SCA效应值变幅为-0.30~3.24,正效应组合8个,以AN-65×AN-2效应值最大;负效应组合2个,以AN-65×AN-76的负效应最大。穗粗SCA效应值变幅为-0.14~0.45,正效应组合8个,以AN-61×AN-65效应值最大;负效应组合2个,以AN-16×AN-65的负效应最大。穗行数SCA效应值变幅为-0.35~0.76,正效应组合7个,效应值最大的是AN-61×AN-65;负效应组合3个,以AN-61×AN-76效应值最大。行粒数SCA效应值变幅为-1.10~7.66,正效应组合9个,效应值最大的是AN-65×AN-2;负效应组合为AN-65×AN-76。秃尖长SCA效应值变幅为-0.23~0.27,增加秃尖长的正效应组合4个,效应值最大的是AN-61×AN-76;减小秃尖长的正效应组合6个,效应值最大的是AN-61×AN-65。从5个穗部性状的SCA效应来看,AN-16×AN-61、AN-61×AN-65、AN-65×AN-2、AN-2×AN-76等5个正交组合的5个穗部性状效应均为促进增产的正效应。
产量性状:10个正交组合百粒重SCA效应值变幅为-1.10~2.82,正效应组合9个,以AN-16×AN-61效应值最大,其次为AN-16×AN-76;AN-16×AN-65的百粒重SCA为负效应。产量SCA效应值变幅为-0.49~1.39,正效應组合8个,AN-65×AN-2效应值最大,AN-16×AN-76其次;负效应组合2个,以AN-16×AN-65的负效应最大。
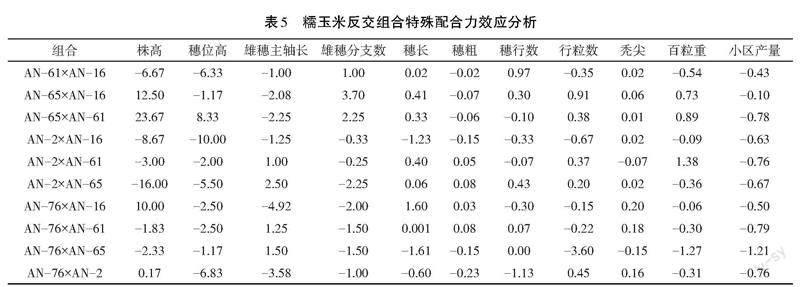
2.4.2 反交组合特殊配合力效应分析
反交组合SCA效应结果(见表5)显示,同一测定性状不同正交组合特殊配合力效应值存在不同差异。
植株性状:10个反交组合株高SCA效应值变幅为-16.00~23.67,正效应的组合4个,以AN-65×AN-61最大;负效应的组合6个,以AN-2×AN-65的负效应最大。穗位高SCA效应值变幅为-10.00~8.33,除AN-65×AN-61为正效应外,其余均为负效应,以AN-2×AN-16负效应最大。雄穗主轴长SCA效应值变幅为-4.92~2.50,正效应值组合4个,以AN-2×AN-65最高;负效应值组合6个,负效应最大的组合是AN-76×AN-2。雄穗分支数SCA效应值变幅为-2.25~3.70,正效应组合3个,效应值最大的是AN-65×AN-16;负效应组合7个,负效应最大的是AN-2×AN-65。
穗部性状:10个反交组合穗长SCA效应值变幅为-1.61~1.60,正效应组合7个,以AN-76×AN-16效应值最大;负效应组合3个,以AN-2×AN-16的负效应最高。穗粗SCA效应值变幅为-0.23~0.08,正效应组合4个,以AN-2×AN-65效应值最大;负效应组合6个,以AN-76×AN-2的负效应最大。穗行数SCA效应值变幅为-1.13~0.97,正效应组合6个,效应值最大的是AN-61×AN-16;负效应组合4个,以AN-76×AN-2效应值最大。行粒数SCA效应值变幅为-3.60~0.91,正效应组合5个,效应值最大的是AN-65×AN-16;负效应组合5个,以AN-76×AN-65的负效应最大。秃尖长SCA效应值变幅为-0.15~0.20,增加秃尖长的正效应组合8个,效应值最大的是AN-2×AN-61;增加秃尖长的负效应组合2个,效应最大的是AN-76×AN-65。
产量性状:10个反交组合百粒重SCA效应值变幅为-1.27~1.38,正效应组合3个,以AN-2×AN-61效应值最大;负效应组合7个,以AN-76×AN-65的负效应最大。产量SCA效应值变幅为-1.21~-0.1,全部为负效应。
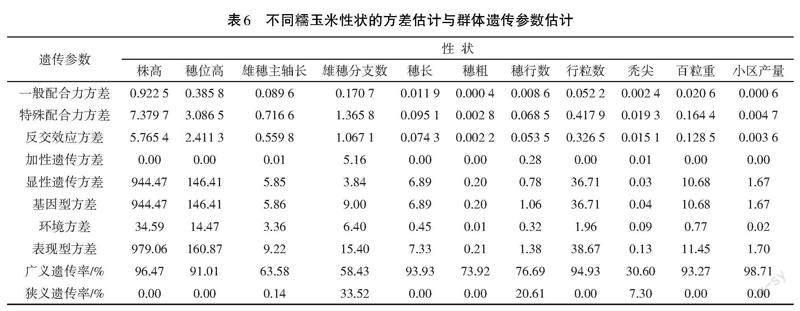
2.5 群体遗传参数估计
一般配合力方差是遗传方差中的加性变异,特殊配合力方差是遗传方差中的非加性变异。遗传力是指通过表型值预测育种值的可靠性大小的比值,即亲代与子代相似程度的指标,反映亲代变异传递到子代的能力。广义遗传率是指基因型方差占表现型方差的比率;狭义遗传率是指加性遗传方差占表现型方差的比例。从表6可以看出,11个农艺性状一般配合力方差均小于特殊配合力方差,除雄穗分支数外,其余株高、穗位高、穗长、穗粗、秃尖、穗行数、行粒数、百粒重、产量等10个性状的显性方差明显大于加性方差。说明除了雄穗分支数外,其余10个性状以显性遗传为主。从11个性状的遗传率来看,除雄穗主轴长、雄穗分支数和秃尖长等3个性状的广义遗传率较低,受环境影响较大外,其余8个性状广义遗传率均较高。狭义遗传力值均较低,说明参试杂交组合的表现型对其所携带基因的平均效应总和依赖性不高。
3 结论与讨论
配合力是评价糯玉米育种基础材料利用价值的重要指标[11]。一般配合力表现的是基因的累加效应,是能够稳定遗传的部分,一般配合力高的材料组配出现高产组合的概率较大[12]。因此,在玉米育种中,尤其是组配杂交组合之前,可以通过配合力评价来预测杂交后代的表现。特殊配合力表现的是基因的非加性效应,不能稳定地遗传给后代,但特殊配合力是针对某一对亲本材料而言,它比一般配合力更接近于杂交组合本身的表现。王向东等认为,一个优良的自交系应该具有较高的一般配合力,同时具有较大的特殊配合力理論方差[13]。
本研究测试的5个自交系中,AN-61和AN-2产量及与产量相关的主要性状的一般配合力最高,其正交组合的特殊配合力也相对较高。具体表现为果穗较粗、长,穗行数和行粒数较多,秃尖小,但是株高、穗位高一般配合力效应值偏高,其后代组合的抗逆性有待进一步鉴定。虽然AN-65、AN-16产量及与产量相关的主要性状的一般配合力较低,但是株高、穗位高一般配合力效应值也低,能有效降低杂交后代的株高、穗位高,有利于组配出植株低、穗位低的杂交组合。
随着鲜食玉米市场的不断壮大和发展,市民对鲜食糯玉米的消费倾向于果穗中等、秃尖少、穗粒稍大、糯性等口感好的商品鲜食糯玉米[14]。但是,农民对品种的选择在注重品质的同时,更注重产量,以追求经济效益最大化。结合市场鲜食糯玉米商品性要求和农民种植偏好实际,在杂交组合组配过程中,应侧重于选择产量较高、果穗形状好、口感佳的品种。在本研究5个亲本组配出的20个正反交组合中,“AN-2×AN-61”“AN-65×AN-61”产量高,穗长、穗粗、穗行数、行粒数、百粒重表现较好,秃尖小,符合农户种植偏好和市场选择喜好,但其鲜食口感和内在品质还有待进一步测定。
参考文献:
[1] 丁照华,孟昭东,张发军,等.我国糯玉米育种现状及发展对策[J].玉米科学,2006,14(3)46-48.
[2] 胡俏强,钱俊,王慧,等.“逆境严选”鲜食玉米自交系选育方法及育种应用[J].上海农业科技,2020(6):77-79.
[3] 姜龙,陈殿元,周岚,等.玉米自选系配合力的分析[J].种子,2019(1):72-75.
[4] 赵久然,卢柏山,史亚兴,等.我国糯玉米育种及产业发展动态[J].玉米科学,2016,24(4):67-71.
[5] 姚坚强,鲍坚东,朱金庆,等.中国糯玉米WX基因种质资源遗传多样性[J].作物学报,2013(1):43-49.
[6] 郝德荣,冒宇翔,陈国清,等.我国鲜食甜糯玉米育种现状与展望[J].浙江农业科学,2016(4):478-481.
[7] 汪黎明,孙琦,孟昭东,等.我国鲜食玉米育种现状及进展分析[J].玉米科学,2005,13(3):35-38.
[8] 傅同良,刘先友.贵州糯玉米育种现状、种质创新及发展方向[J].种子,2008(6):78-81.
[9] 袁雨晴,邹军,李士敏,等.贵州山区鲜食糯玉米丰产稳产性分析及评价[J].种子,2017(5):74-78.
[10] 邹军,章洁琼,龙英,等.20个鲜食玉米品种产量与主要性状的灰色关联度分析[J].种子,2019(11):138-142.
[11] 莫润秀,黄开健,黄爱花,等.20个CIMMYT耐低氮玉米自交系主要性状的配合力分析[J].西南农业学报,2019(12):2732-2739.
[12] 吴宏亮,康建宏,张立杰,等.8个玉米自交系穗部性状配合力分析[J].玉米科学,2013,45(10):20-23.
[13] 王向东,高根来,张凤琴,等.玉米产量性状配合力遗传分析[J].玉米科学,2001,9(1):31-33.
[14] 孙丽娟,赵志宏,贺娟,等.我国现实玉米相关标准问题分析及对策[J].作物杂志,2019(2):46-50.
(责任编辑:丁志祥)
收稿日期:2023-03-17
基金项目:安顺市科技计划项目“青贮玉米种质资源鉴选与新品种选育”(安市科农〔2022〕03号);贵州省科技计划项目“优质高产糯玉米新品种顺糯007应用与示范”(黔科合成果〔2023〕一般035)。
作者简介:赵艳花(1978—),女,贵州普定人,本科,高级农艺师,主要从事作物遗传育种和栽培技术研究。E-mail:nkszhyh@163.com。
猜你喜欢
北京农学院学报(2019年1期)2019-02-22
农民致富之友(2018年23期)2018-12-19
河南农业科学(2017年4期)2017-04-12
中国中药杂志(2016年20期)2016-11-19
西南农业学报(2016年5期)2016-05-17
现代食品(2016年24期)2016-04-28
西南农业学报(2016年6期)2016-04-16
上海农业学报(2016年5期)2016-02-10
食品科学(2013年17期)2013-03-11
