
中国小麦抗穗发芽种质资源的挖掘与创制
2023-11-22 08:29王旭阳余赵玉张海萍司红起马传喜
安徽农业大学学报 2023年5期
常 成,王旭阳,余赵玉,张海萍,卢 杰,司红起,陈 璨,马传喜
中国小麦抗穗发芽种质资源的挖掘与创制
常 成,王旭阳,余赵玉,张海萍,卢 杰,司红起,陈 璨,马传喜*
(安徽农业大学农学院/农业农村部黄海南部小麦生物学与遗传育种重点实验室,合肥 230036)
根据4年的表型数据,并结合实验室开发和鉴定的13个分子标记,对我国833份小麦种质资源(主要包括278份小麦微核心种质、124份地方品种和431份现代推广品种及高代品系)穗发芽抗性进行鉴定。结果表明,13个分子标记鉴定的抗/感穗发芽等位类型间相对发芽指数(RGI)差异均达显著或极显著水平,其中TaMFT-222 和TaMFT-194 标记鉴定的差异最大,U 值分别为14.98**和11.30**,均达极显著水平,其优异等位类型可以降低相对发芽指数0.21 ~ 0.32。其次是Sdr2A、CNGC2AL、Vp1-b2、TaMKK3-A、PM19、CAPS-2AL、A17-19和EX06323标记,其等位类型间穗发芽抗性差异也均达极显著水平;Qsd1和Barc321标记也能显著区分穗发芽抗性。共计鉴定出63份穗发芽抗性较好的种质资源,其中达到抗的有41份,多为红皮品种和地方品种;达到中抗的有22份,白皮半冬性居多。利用分子标记辅助选择,并结合杂交聚合,创制出12份穗发芽抗性水平达到中抗和抗的种质资源,至少携带3个抗穗发芽基因/位点。该结果为抗穗发芽新品种选育提供重要遗传资源。
小麦;抗穗发芽;分子标记;种质资源
小麦收获前遇阴雨天气导致籽粒在穗部萌发的现象称之为穗发芽,常导致减产、品质劣化以及种用丧失等,是影响小麦安全生产的主要非生物逆境之一。我国长江中下游、西南冬麦区及北部春麦区常受到穗发芽的危害;但随着气候变化,小麦成熟期雨带北移,黄淮麦区南部也时常发生穗发芽[1]。作为我国小麦主产麦区,黄淮麦区种植的是白皮半冬性小麦品种,穗发芽抗性普遍较弱,无法有效应对穗发芽问题,已成为制约该区域小麦安全生产的主要因素之一。
小麦生产实践表明,挖掘和应用抗穗发芽种质资源、培育抗穗发芽小麦新品种是解决这一问题的主要途径和有效手段。种子休眠特性是控制小麦穗发芽抗性的关键因子,休眠性强的品种,其穗发芽抗性也较强,反之则较弱[1]。前人对种子休眠及穗发芽抗性的遗传机制开展了大量研究,结果表明种子休眠属于复杂的数量性状,由基因型和环境共同决定,控制小麦种子休眠/穗发芽抗性的QTL位点遍及小麦21条染色体[1]。目前利用同源克隆或图位克隆方法已分离出6个基因,如[2]、[3]、[4]、[5-6]、[7]和[8]。
本文通过表型鉴定,并利用实验室开发和鉴定的种子休眠以及抗穗发芽分子标记,对我国小麦种质资源进行鉴定,并创制抗穗发芽新种质资源,为小麦抗穗发芽品种培育提供重要基因资源。
1 材料与方法
1.1 材料
实验室收集的833份国内小麦种质资源,主要包括小麦微核心种质(278份)、地方品种(124份)、现代推广品种及高代品系(431份)等。
1.2 田间试验与设计
于2018—2021年度秋播于安徽农业大学合肥大杨店试验站,每份试验材料种植2行,行长2 m,行距0.25 m,每行40粒,随机区组排列,常规田间管理。适期播种,常规栽培管理与病虫害防治。在开花当天选择有代表性的10 ~ 15个植株主茎穗,挂牌并注明开花日期。于开花后第35天或小麦生理成熟期(即黄熟期)收获10个挂牌的麦穗,脱粒后测定相对发芽指数(RGI)。

表1 小麦抗穗发芽性评价标准

表2 小麦抗PHS基因/位点分子标记信息
1.3 相对发芽指数测定(RGI, relative germination rate)
测定方法参考农业农村部穗发芽鉴定行业标准(NY/T 1739—2009)[9]和高威[10]等的方法,加以优化与改进。以中优9507为对照品种。取100粒胚部完整发育正常的小麦籽粒,腹股沟朝下均匀的摆放在已灭菌铺有发芽纸的培养皿(Φ90 mm)中,加入10 mL无菌水,2个重复。将培养皿放入人工气候室进行发芽实验(22 ℃、100% RH),光照时间为为14 h/10 h(白天/黑夜),连续培养48 h。
分别测定每份材料和对照品种中优9507的平均发芽率GI,然后根据以下公式,计算相对发芽指数RGI,作为评估穗发芽抗性的指标,具体如表1。
=(穗发芽籽粒数/总籽粒数)×100。
= 待测样品平均发芽率/对照品种发芽率
1.4 DNA提取及引物设计
DNA提取参照Chang等[11]方法完成。本实验室所用分子标记及其引物如表2所示,特异引物合成由上海生物工程有限公司完成。功能标记检测所需的限制性核酸内切酶在NEB和TaKaRa公司订购。PCR扩增、酶切和产物琼脂糖凝胶电泳检测均参考前人报道。
2 结果与分析
2.1 年份间相对发芽指数分析
检测了2018—2021年度833份小麦种质资源的相对发芽指数,结果如表3所示。年度间变异系数差异较大,其中2018年度RGI变异系数最大,为51%;其次是2019年度,为48%;最低的为2020年度,变异系数为38%。
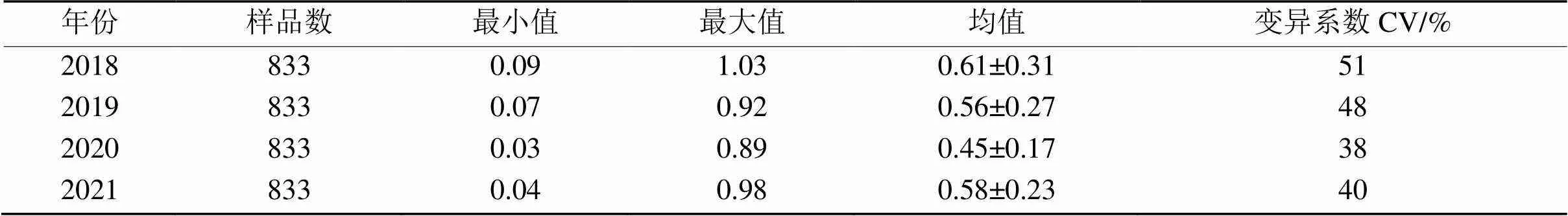
表3 相对发芽指数RGI分析
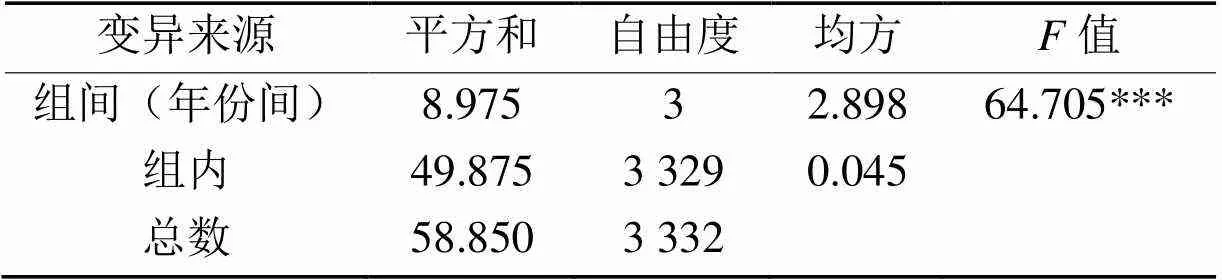
表4 年份间RGI方差分析
注:***指差异显著性达0.001水平。下同。

表 5 年份间RGI相关性分析
不同年份间的RGI差异达极显著水平(表4),说明该性状除了受基因型控制之外,也受环境影响。由表5可知,不同年份间,相关性均达极显著水平,其中2018和2020年度RGI相关性最好,相关系数为0.702***。说明本文测定的RGI值在不同环境下具有较好的一致性。
2.2 抗穗发芽基因型鉴定及分析
本文根据实验室挖掘以及前人报道的12个抗穗发芽基因/位点,开发和鉴定了13个分子标记,对833 份国内小麦种质资源进行基因型检测。根据4年的RGI均值,分析这些基因/位点的抗性等位类型(R)和感性等位类型(S)间的差异显著性,以确定基因/位点对穗发芽抗性的作用。从表6可看出,通过U-test,这13个标记鉴定的抗/感穗发芽类型间相对发芽指数差异均达显著或极显著水平,其中TaMFT-222 和TaMFT-194 标记鉴定的抗/感类型间差异最大,U 值分别为14.98**和11.30**,均达极显著水平,平均可以降低相对发芽指数0.21 ~ 0.32,在分子标记辅助选择中具有较高的应用价值。其次是Sdr2A、CNGC2AL 、Vp1-b2、TaMKK3-A、PM19、CAPS-2AL、A17-19和EX06323标记,其等位类型间穗发芽抗性差异也均达极显著水平;Qsd1和Barc321标记也能显著区分穗发芽抗性,具有一定的应用价值。在鉴定的12个基因/位点中,抗穗发芽等位类型的相对发芽指数均显著低于感性等位类型,但对穗发芽抗性表型变异的作用不同,与前期研究结果较为一致[14]。
2.3 抗穗发芽种质资源挖掘与鉴定
对833份小麦种质资源进行抗穗发芽测定,并结合分子标记辅助鉴定,共挖掘63份抗性达中抗及以上的品种/系。其中,安农1118、安农1124、安农1121及扬麦18528等41个品种/系为抗穗发芽,安农196、安农0711、百农64及皖垦麦081等品种/系为中抗穗发芽。由表7可知:本文所用的试验材料中,抗穗发芽品种/系多为红皮品种或地方农家种;中抗穗发芽品种多为白皮半冬性品种;未鉴定出高抗穗发芽品种所示。
2.4 抗穗发芽育种新材料创制
本实验室长期开展小麦抗穗发芽分子机制、育种材料创新及新品种选育等研究工作,根据已挖掘和鉴定的基因/位点,开发出13个分子标记用于抗穗发芽辅助选择,并结合常规育种手段,聚合抗穗发芽基因。本研究创制抗穗发芽育种新材料12份,其中白皮半冬性5份(中抗),红皮品种7份(抗),如表8所示。通过分子标记鉴定,这些育种新材料包含3 ~ 4个抗穗发芽基因/位点,主要位于2AL()、3AS()、3BL()、4AL()、5B()和6B()染色体上。在抗穗发芽基因/位点中,(3AS)、(3AS)、(3BL)和(4AL)出现的频率较多(表8)。
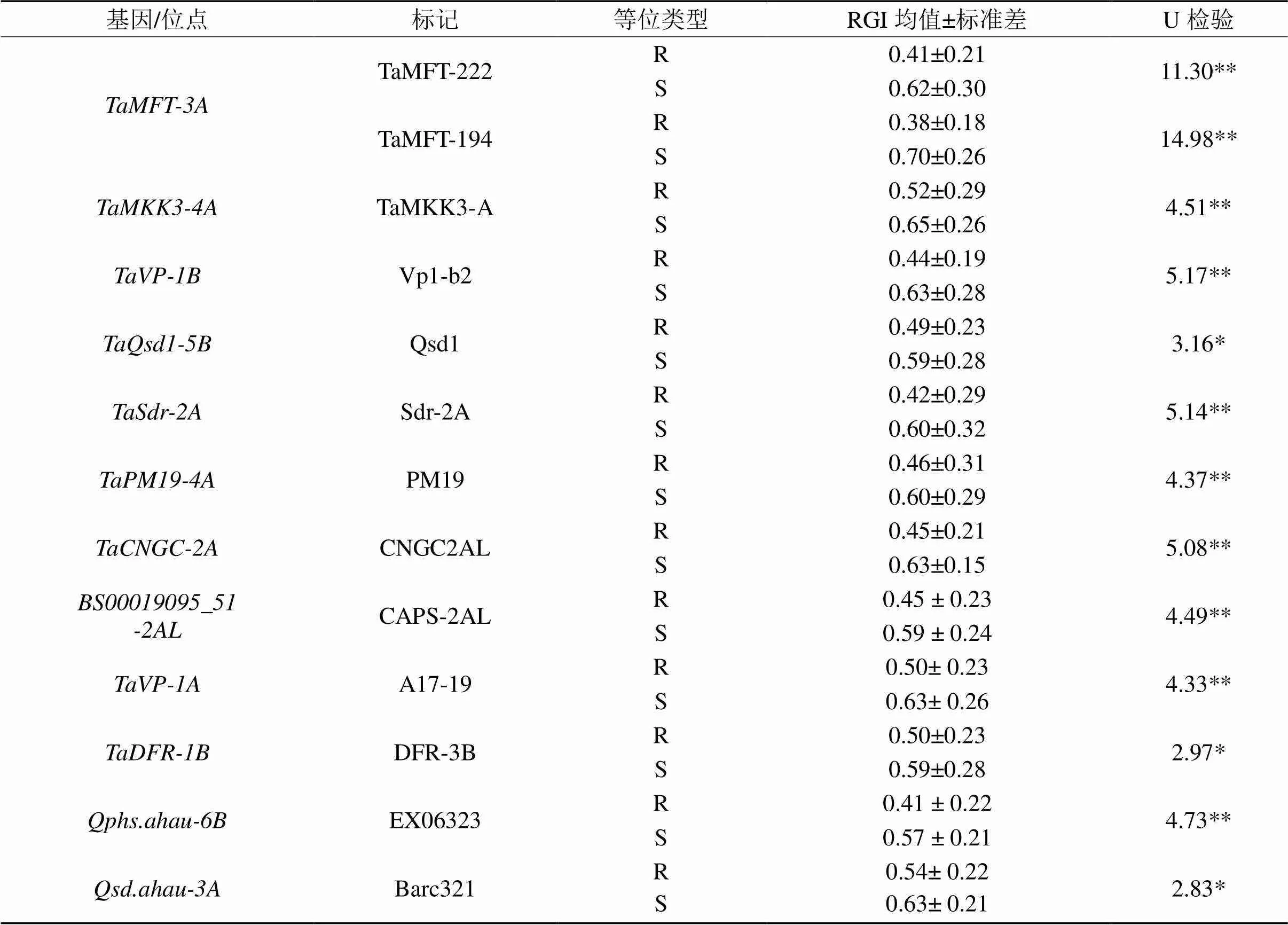
表6 抗穗发芽基因鉴定及其对表型的作用分析
注:R:抗穗发芽等位类型;S:感穗发芽类型。*和**指差异显著性分别达0.05和0.01水平。
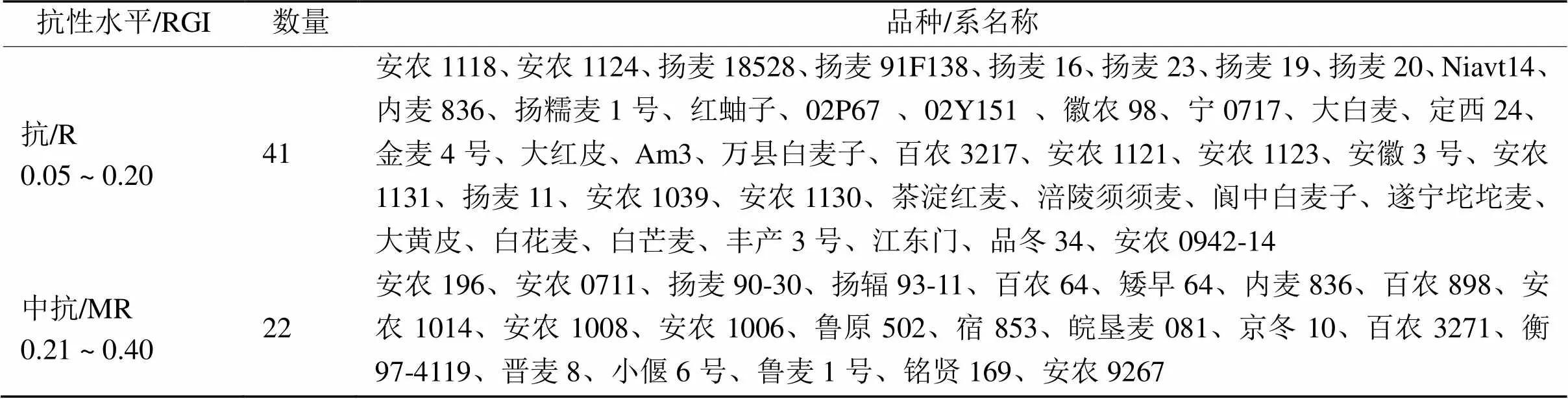
表7 中抗穗发芽及以上的小麦品种/系
3 讨论与结论
小麦穗发芽是一种严重的非生物逆境,一旦发生,难防难控,对小麦生产往往造成重大损失。选育和应用抗穗发芽小麦品种是解决这一问题的主要途径,而挖掘和应用抗穗发芽优异种质资源是新品种选育的前提[1]。小麦穗发芽既受环境因素影响,如成熟期降水等,也与品种本身的穗发芽抗性密切相关。种子休眠特性是影响穗发芽抗性的关键因子,均受多基因控制,聚合有效抗穗发芽基因/位点是提高抗性的主要手段[1, 3]。本研究结果表明,不同抗穗发芽基因/位点对于抗性的作用也不同,其中2A(、)、3AS(、)和3BL()作用较大,而和等在抗穗发芽品种/系中分布较大(结果未列出)。本实验室在前期的研究中,利用开发及鉴定的抗穗发芽分子标记进行辅助选择,聚合多个抗穗发芽基因/位点,创制出多份抗穗发芽育种新材料,选育出安农0711、安农1124等抗穗发芽小麦新品种。从种质资源创制、新品种的选育等育种实践来看,分子标记辅助选择可有效用于抗穗发芽遗传改良。
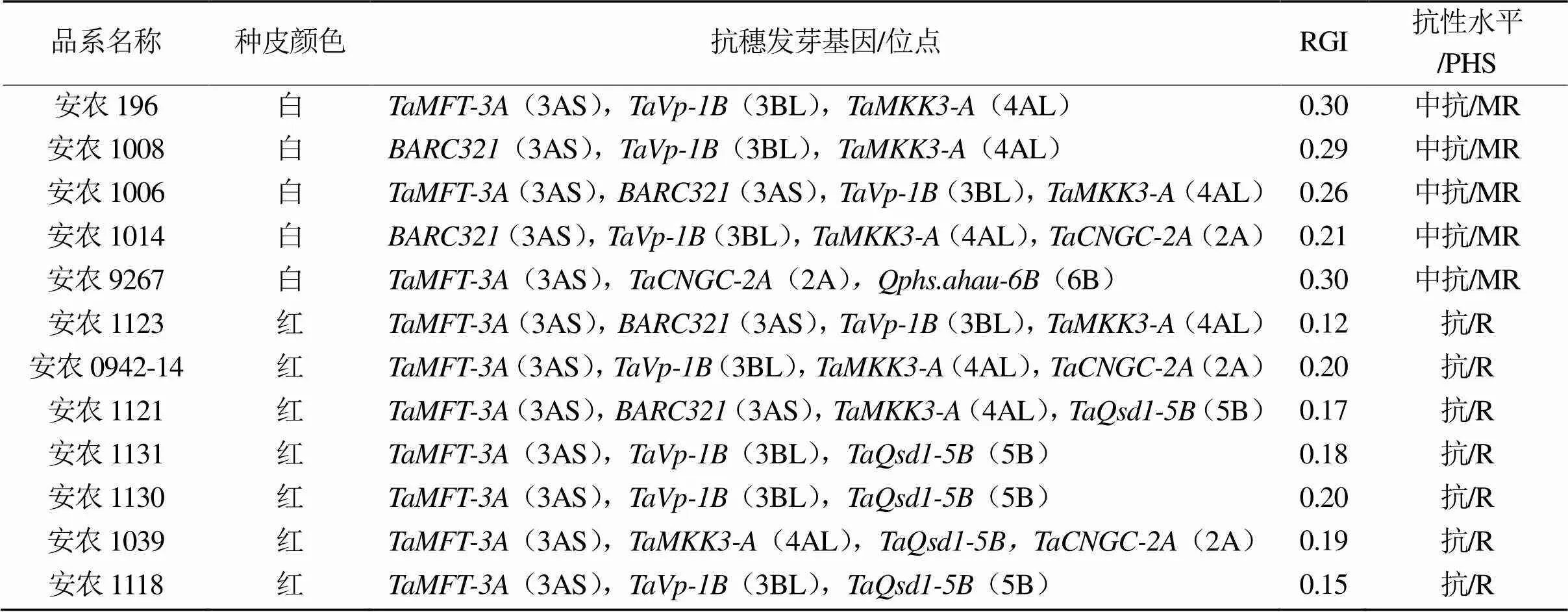
表8 小麦育种新材料穗发芽抗性及抗性基因/位点
优异的抗穗发芽种质资源是培育抗穗发芽新品种的前提,是“种子芯片”,对挖掘和创制优异种质资源尤为重要。在穗发芽抗性鉴定时,前人多采用籽粒发芽指数和穗发芽指数进行评价,由于穗发芽易受环境影响,这些方法对于测定不同环境下的穗发芽抗性会产生不可避免的误差[3-4,7,18]。因此,本文采用相对发芽指数来测定穗发芽抗性,可有效减少环境因素的影响,提高鉴定的准确性。而且,本文综合分析4年的穗发芽抗性测定数据,保证了鉴定结果的可靠性。
本文挖掘和鉴定了63份抗穗发芽品种/系,其中抗性品种多为红皮品种,白皮品种多为中抗水平。创制的抗穗发芽育种新材料中,白皮品种穗发芽抗性也低于红皮品种,这与前人结果一致[1,18]。在我国核心种质和地方农家品种中,也有穗发芽抗性好的白皮品种。这些种质资源和育种新材料对于抗穗发芽品种培育具有重要的应用价值。
[1] 肖世和, 闫长生, 张海萍, 等. 小麦穗发芽研究[M]. 北京:中国农业科学技术出版社, 2004.
[2] YANG Y, ZHAO X L, XIA L Q, et al. Development and validation of aSTS marker for pre-harvest sprouting tolerance in Chinese wheats[J]. Theor Appl Genet, 2007, 115(7): 971-980.
[3] BARRERO J M, CAVANAGH C, VERBYLA K L, et al. Transcriptomic analysis of wheat near-isogenic lines identifies PM19-A1 and A2 as candidates for a major dormancy QTL[J]. Genome Biol, 2015, 16(1): 1-18.
[4] NAKAMURA S, ABE F, KAWAHIGASHI H, et al. A wheat homolog ofacts in the regulation of germination[J]. Plant Cell, 2011, 23(9): 3215-3229.
[5] ZHANG Y J, MIAO X L, XIA X C, et al. Cloning of seed dormancy genes () associated with tolerance to pre-harvest sprouting in common wheat and development of a functional marker[J]. Theor Appl Genet, 2014, 127(4): 855-866.
[6] ZHANG Y J, XIA X C, HE Z H. The seed dormancy allele-A1a associated with pre-harvest sprouting tolerance is mainly present in Chinese wheat landraces[J]. Theor Appl Genet, 2017, 130(1): 81-89.
[7] TORADA A, KOIKE M, OGAWA T, et al. A causal gene for seed dormancy on wheat chromosome 4A encodes a MAP kinase kinase[J]. Curr Biol, 2016, 26(6): 782-787.
[8] WEI W X, MIN X Y, SHAN S Y, et al. Isolation and characterization ofgenes for period of dormancy in common wheat (L.)[J]. Mol Breeding, 2019, 39(10): 150.
[9] 中华人民共和国农业部. 小麦抗穗发芽性检测方法(NY/T 1739—2009)[S]. 北京: 中国标准出版社, 2009.
[10] 高威, 徐康乐, 刘雪, 等. 小麦穗发芽抗性鉴定标准的改进与优化[J]. 安徽农业大学学报, 2021, 48(2): 179-184.
[11] CHANG C, ZHANG H P, FENG J M, et al. Identifying alleles ofassociated with pre-harvest sprouting in micro-core collections of Chinese wheat germplasm[J]. Mol Breeding, 2010, 25(3): 481-490.
[12] JIANG H, ZHAO L X, CHEN X J, et al. A novel 33-bp insertion in the promoter ofis associated with pre-harvest sprouting resistance in common wheat[J]. Mol Breeding, 2018, 38(5): 69.
[13] 朱玉磊. 小麦2A染色体抗穗发芽主效QTL鉴定与候选基因挖掘[D]. 合肥: 安徽农业大学, 2017.
[14] ZHU Y L, WANG S X, ZHANG H P, et al. Identification of major loci for seed dormancy at different post-ripening stages after harvest and validation of a novel locus on chromosome 2AL in common wheat[J]. Mol Breed, 2016, 36(12): 174.
[15] CHANG C, ZHANG H P, ZHAO Q X, et al. Rich allelic variations ofand their associations with seed dormancy/pre-harvest sprouting of common wheat[J]. Euphytica, 2011, 179(2): 343-353.
[16] BI H H, SUN Y W, XIAO Y G, et al. Characterization of DFR allelic variations and their associations with pre-harvest sprouting resistance in a set of red-grained Chinese wheat germplasm[J]. Euphytica, 2014, 195(2): 197-207.
[17] ZHU Y L, WANG S X, WEI W X, et al. Genome-wide association study of pre-harvest sprouting tolerance using a 90K SNP array in common wheat (L.)[J]. Theor Appl Genet, 2019, 132(11): 2947-2963.
[18] 朱玉磊, 王升星, 赵良侠, 等. 以关联分析发掘小麦整穗发芽抗性基因分子标记[J]. 作物学报, 2014, 40(10): 1725-1732.
Excavation and creation of pre-harvest sprouting resistant germplasm resources in Chinese wheats
CHANG Cheng, WANG Xuyang, YU Zhaoyu, ZHANG Haiping, LU Jie, SI Hongqi, CHEN Can, MA Chuanxi
(School of Agronomy, Anhui Agricultural University/Key Laboratory of Wheat Biology and Genetic Improvement on South Yellow & Huai River Valley, Ministry of Agriculture and Rural Affairs, Hefei 230036)
Based on four years' phenotypic data, and combined with 13 molecular markers developed and identified in our laboratory, 833 wheat germplasm resources (including 278 wheat micro-core germplasms, 124 local varieties and 431 modern promoted varieties and advanced lines) were identified for pre-harvest sprouting (PHS) resistance. The results showed that the differences in relative germination index (RGI) were significant or extremely significant between the resistance and sensitive alleles (R/S) of the 13 molecular markers, respectively. Among these markers, the largest difference between R/S was identified in TaMFT-222 and TaMFT-194 markers, which U values were 14.98** and 11.30**, respectively, and reached the extremely significant level. The RGI could be reduced from 0.21 to 0.32 by the two resistant alleles, respectively. The next markers were Sdr2A, CNGC2AL, Vp1-b2, TaMKK3-A, PM19, CAPS-2AL, A17-19 and EX06323, as well as the significant differences in PHS resistance were also detected among R/S alleles. The Qsd1 and Barc321 markers could also significantly distinguish PHS resistance between R/S alleles. A total of 63 germplasm resources with high PHS resistance were identified in this study. The 41 germplasms mainly including red-grained and local varieties had higher PHS resistance than that of the 22 germplasms mostly with the white grained. Through using hybridization combined with marker-assisted selection, the PHS resistance genes/loci were pyramided, and 12 new breeding materials with high pre-harvest sprouting resistance were created, which carried at least 3 genes/loci for PHS resistance. The result can provide important genetic resources for breeding new varieties with high PHS resistance.
wheat; pre-harvest sprouting resistance; molecular marker; germplasm resources
S512.102.4
A
1672-352X (2023)05-0745-06
10.13610/j.cnki.1672-352x.20231030.014
2023-10-31 08:23:19
[URL] https://link.cnki.net/urlid/34.1162.S.20231030.1110.026
2022-11-15
国家现代小麦产业技术体系专项(CARS-03),国家自然科学基金联合基金(U20A2033),安徽高校协同创新项目(GXXT-2021-058)和安徽省科技重大专项(2021d06050003)共同资助。
常 成,教授。E-mail:changtgw@126.com
马传喜,教授,博士生导师。E-mail:machuanxi@ahau.edu.cn
猜你喜欢
今日农业(2022年13期)2022-09-15
作物学报(2022年6期)2022-04-08
国际医学放射学杂志(2021年5期)2021-10-22
空间科学学报(2021年1期)2021-05-22
麦类作物学报(2018年4期)2018-05-11
中国麻业科学(2018年6期)2018-04-09
环境保护与循环经济(2017年5期)2018-01-22
第一财经(2017年36期)2017-09-25
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
