
磷硒配施对冬小麦硒吸收、转运及籽粒有机硒的影响
2023-11-22 11:07胡彩霞刘红恩李畅秦世玉赵玉焕张玉鹏刘亥扬许嘉阳赵鹏聂兆君王秋红
中国农业科技导报 2023年11期
胡彩霞, 刘红恩, 李畅, 秦世玉, 赵玉焕, 张玉鹏, 刘亥扬,许嘉阳, 赵鹏, 聂兆君*, 王秋红
(1.河南农业大学资源与环境学院, 土壤污染防控与修复重点实验室, 郑州 450002; 2.河南省台前县农业农村局, 河南 濮阳 457600; 3.郑州市农业生态保护服务中心, 郑州 450006)
硒(Se)是人类和动物所必需的微量元素之一,在提高人体免疫力、抗氧化、防癌抗癌及防治心脑血管疾病等方面具有重要作用[1]。当膳食中硒摄入量低于40 µg·d-1时会造成人体硒的缺乏,加速机体衰老,导致癌症、克山病及大骨病等疾病的发生[2-3]。有研究[4]表明,我国72%的地区土壤中严重缺硒,导致农产品中硒含量过低。土壤作为作物吸收硒的主要来源,通过土施硒肥提高农产品硒含量是人体安全补硒的重要途径[5]。土壤中的有效硒被植物吸收利用后,可以转化为对人体安全的有机硒形态,包括硒蛋白、硒代氨基酸、硒多糖及硒核酸等[6-9]。
磷(P)是植物生长必需的第二大营养元素,在植物生长、代谢、提高产量和品质中发挥着不可替代的作用[10]。磷酸盐与亚硒酸盐化学性质相似,均以阴离子形态被作物吸收,因此二者在土壤-植物系统中可能存在相互作用[11]。赵文龙等[12]发现,磷硒配施水平直接影响硒在植株中的转运及吸收累积。施用磷肥抑制硒从根系向地上部的转运,显著降低植株硒含量,磷与硒存在拮抗作用[13-14]。Wang 等[15]在磷缺乏条件下,添加1 mmol·L-1KH2PO4降低了水稻根系硒含量,但提高了水稻地上部硒含量,且影响水稻幼苗体内有机硒形态转化。也有报道称磷硒在土壤溶液中存在吸附竞争关系,磷肥的施用能提高土壤中有效硒含量,进而促进冬小麦对硒的吸收和累积[16-18]。因此,关于磷硒配施在植物体内的相互影响不尽一致。有研究表明,硒在水稻体内的分配表现为叶>茎>籽粒[19];小白菜对硒的吸收和富集能力表现为地上部高于根部[20]。小麦各部位对硒的吸收累积规律也存在差异,马小艳等[21]研究发现,施用硒酸钠后冬小麦各器官硒的分配表现为叶片>籽粒>茎>颖壳。
小麦是我国重要的粮食作物,其籽粒中的硒代氨基酸安全性高,易被人体吸收利用。因此,提高籽粒总硒及有机硒含量对于人体合理补硒具有重要作用。目前,相关研究主要集中在磷硒配施对植株生长、硒含量和土壤中磷、硒有效性的影响等方面[22-25],但关于冬小麦各部位硒含量分布及籽粒有机硒形态转化的研究鲜有报道。因此,本研究通过分析磷硒配施对冬小麦硒吸收、转运及籽粒有机硒的影响,以期为生产富硒小麦及合理施用磷肥与硒肥提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2019 年10 月至2020 年6 月在河南农业大学科教园区试验场进行。供试土壤类型为壤质潮土,基本理化性质为:pH 7.30,有机质含量11.04 mg·kg-1,碱解氮含量70.42 mg·kg-1,有效磷含量10.31 mg·kg-1,速效钾含量140.65 mg·kg-1,有效硒含量0.004 mg·kg-1,全硒含量0.174 mg·kg-1。
1.2 试验材料
供试小麦品种为‘郑麦379’,为河南省广泛种植的冬小麦品种,种子购于河南秋乐种业科技股份有限公司。供试硒试剂为亚硒酸钠(Na2SeO3·5H2O),磷试剂为磷酸二氢钠(NaH2PO4·2H2O);底肥分别为尿素(CH4N2O)、氯化钾(KCl)。所有供试试剂均为分析纯。
1.3 试验设计
试验以NaH2PO4·2H2O 为磷源,设置0(P0)、80(P80)和160 mg·kg-1(P160)3 个磷水平;以Na2SeO3·5H2O 为硒源,设置0(Se0)和1 mg·kg-1(Se1)2 个硒水平;共6 个处理,分别为P0Se0、P80Se0、P160Se0、P0Se1、P80Se1、P160Se1,每个处理重复4 次,共计24盆。采用盆栽试验,聚乙烯塑料盆型号为直径310 mm、深度200 mm,把过10目筛的风干土样8.0 kg装入盆中。每盆播种12粒小麦种子,定株8 棵。试验场设有塑料膜防雨棚以防止雨水淋洗。底肥用量按照每千克土壤施N 0.2 g、K2O 0.2 g,分别以尿素(CH4N2O)、氯化钾(KCl)作为肥源,并于拔节期每盆追施尿素1.0 g。生长期间保持土壤含水量为田间持水量的70%,每3 d 浇1次水。
1.4 测定项目和方法
参照常规分析方法[26]测定土壤pH、有机质、碱解氮、有效磷及速效钾含量。土壤有效硒的测定参照陈娥等[27]的方法。土壤总硒含量的测定参照高雯雯等[28]的方法。植株磷含量的测定参照鲍士旦[26]的方法。
于成熟期采集冬小麦植株样品。植株硒含量的测定参照 GB 5009.93—2017《食品中硒的测定》[29]。植株硒形态的测定参照王铁良等[30]方法:称取过0.150 mm 筛的植株试样0.050 0 g(精确至0.000 1 g)于15 mL 离心管中,加入5 mL Tris-HCl摇匀后超声30 min;再加入50 mg 纤维素酶和20 mg 蛋白酶K摇匀,在(50±2) ℃以250 r·min-1恒温振荡18 h。加入20 mg蛋白酶XIV,在(37±2) ℃以250 r·min-1恒温振荡18h,然后在4 ℃以10 000 r·min-1离心30 min。提取滤液过0.22 µm水系滤膜后用SA-50高效液相色谱-氢化物发生-原子荧光光谱联用仪(SA-50-HPLC-HG-AFS)测定硒形态。
1.5 数据计算、统计与分析方法
本研究用到的计算公式如下。
试验数据均为4 个重复的平均值,采用DPS 7.05版进行数据处理,采用LSD 法(P<0.05)进行多重比较,F(P)表示施磷的主效应、F(Se)表示施硒的主效应、F(P×Se)表示磷硒的交互效应。采用Excel 2003进行处理和统计分析。
2 结果与分析
2.1 磷硒配施对冬小麦干物质量的影响
由表1 可知,无论施硒与否,施磷显著增加冬小麦各部位的干物质量。与Se0处理相比,Se1处理显著降低P160水平下茎叶、颖壳、籽粒干物质量以及总生物量,其降幅分别为11.10%、15.18%、7.21%和9.44%,但显著提高P80水平下根系和籽粒干物质量,其增幅分别为17.44%和45.72%。磷硒配施情况下,在P80Se1处理下冬小麦总生物量达到最大,说明适宜的磷硒配施可促进冬小麦生长。
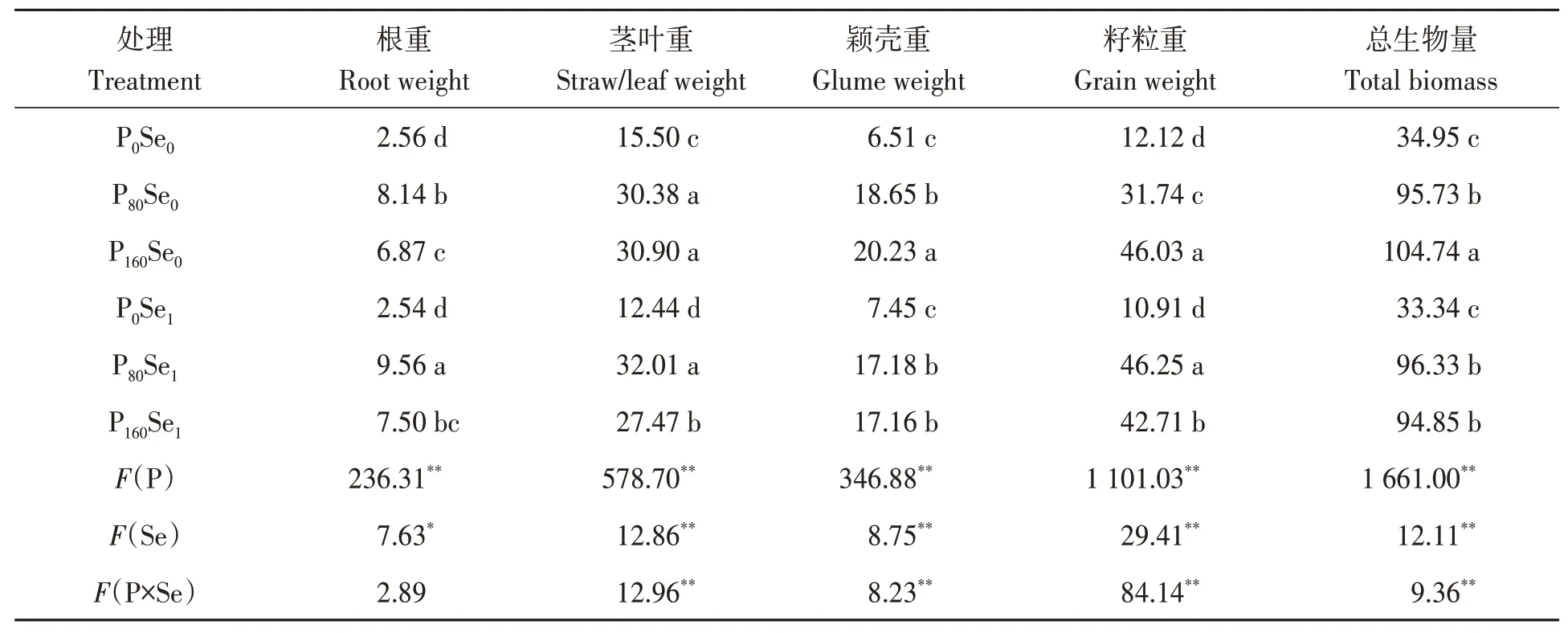
表1 磷硒配施下冬小麦各部位干物质的量Table 1 Dry weight in winter wheat under phosphorus and selenium combined application (g·盆-1/g·pot-1)
2.2 磷硒配施对冬小麦籽粒产量构成要素的影响
由表2可知,施磷显著提高了‘郑麦379’的穗数、穗粒数及千粒重,其增幅为5.35%~231.43%。与Se0处理相比,施硒显著增加了P80水平下‘郑麦379’的穗数、穗粒数,其增幅为15.03%~18.92%,但显著降低了P160水平下穗粒数,降幅为12.16%。冬小麦穗数和穗粒数在P80Se1处理下达到最高,说明适宜的磷硒配施可提高冬小麦产量。
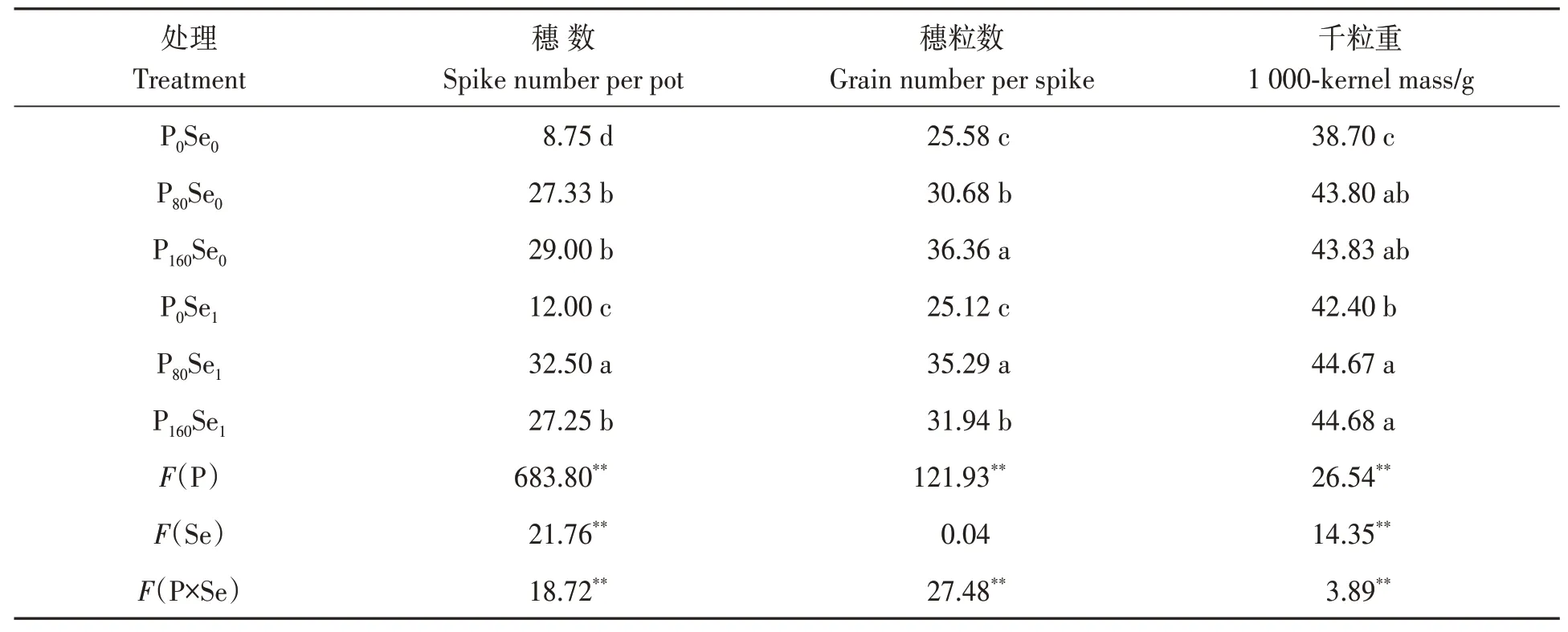
表2 磷硒配施下冬小麦籽粒产量及产量构成要素Table 2 Grain yield its components of winter wheat under phosphorus and selenium combined application
2.3 磷硒配施对冬小麦植株各部位磷含量及磷迁移系数的影响
由表3可知,施磷均显著增加‘郑麦379’各器官磷含量。与Se0处理相比,施硒显著降低了P80和P160水平下‘郑麦379’各器官磷含量,其降幅为11.54%~41.38%。根系、茎叶、颖壳和籽粒磷含量的最大值均出现在P160Se0处理,说明施硒抑制冬小麦对磷的吸收。在Se0水平下,施磷显著增加了P80和P160水平下茎叶向颖壳的磷迁移系数,其增幅分别为38.78%和28.57%,但显著降低了颖壳向籽粒的磷迁移系数,其降幅分别为29.25%和25.69%。在Se1水平下,施磷显著降低了根系向茎叶的磷迁移系数,P80和P160水平下降幅为68.87%和70.82%,但显著增加了颖壳向籽粒的磷迁移系数,其增幅分别为36.04%和15.23%,说明施硒改变磷在冬小麦中的转运,施硒可能抑制冬小麦根系对磷的吸收从而降低磷从根系向茎叶的迁移。
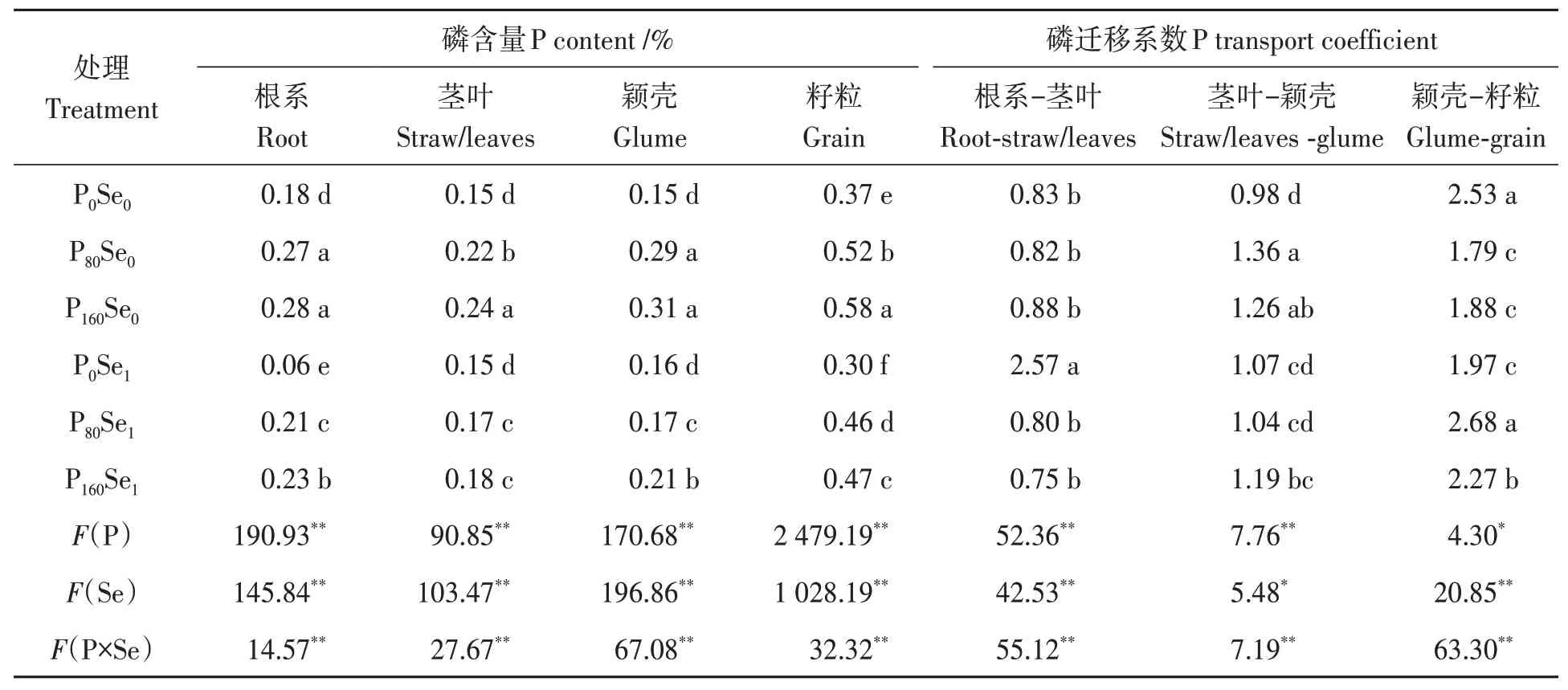
表3 磷硒配施下冬小麦各部位磷含量及迁移系数Table 3 Phosphorus content and migration coefficient of winter wheat under phosphorus and selenium combined application
2.4 磷硒配施对冬小麦植株各部位磷累积量和分配比例的影响
由表4 可知,在Se0和Se1水平下施磷均显著增加了‘郑麦379’籽粒、颖壳、茎叶和根系的磷累积量。与Se0处理相比,施硒显著降低了P80和P160水平下‘郑麦379’籽粒、颖壳、茎叶磷累积量以及总磷累积量,其降幅为13.82%~45.97%。籽粒和总磷累积量的最小值和最大值分别出现在P0Se1和 P160Se0处理。在Se0水平下,施磷增加了籽粒和颖壳的磷累积量分配比例,其增幅为15.60%~26.37%,但降低了茎叶的磷累积量分配比例,其降幅分别为37.34%和42.12%。在Se1水平下,施磷增加了籽粒和根系的磷累积量的分配比例,其增幅为22.27%~218.58%,但降低了颖壳和茎叶的磷累积量分配比例,其降幅为32.43%~44.17%。
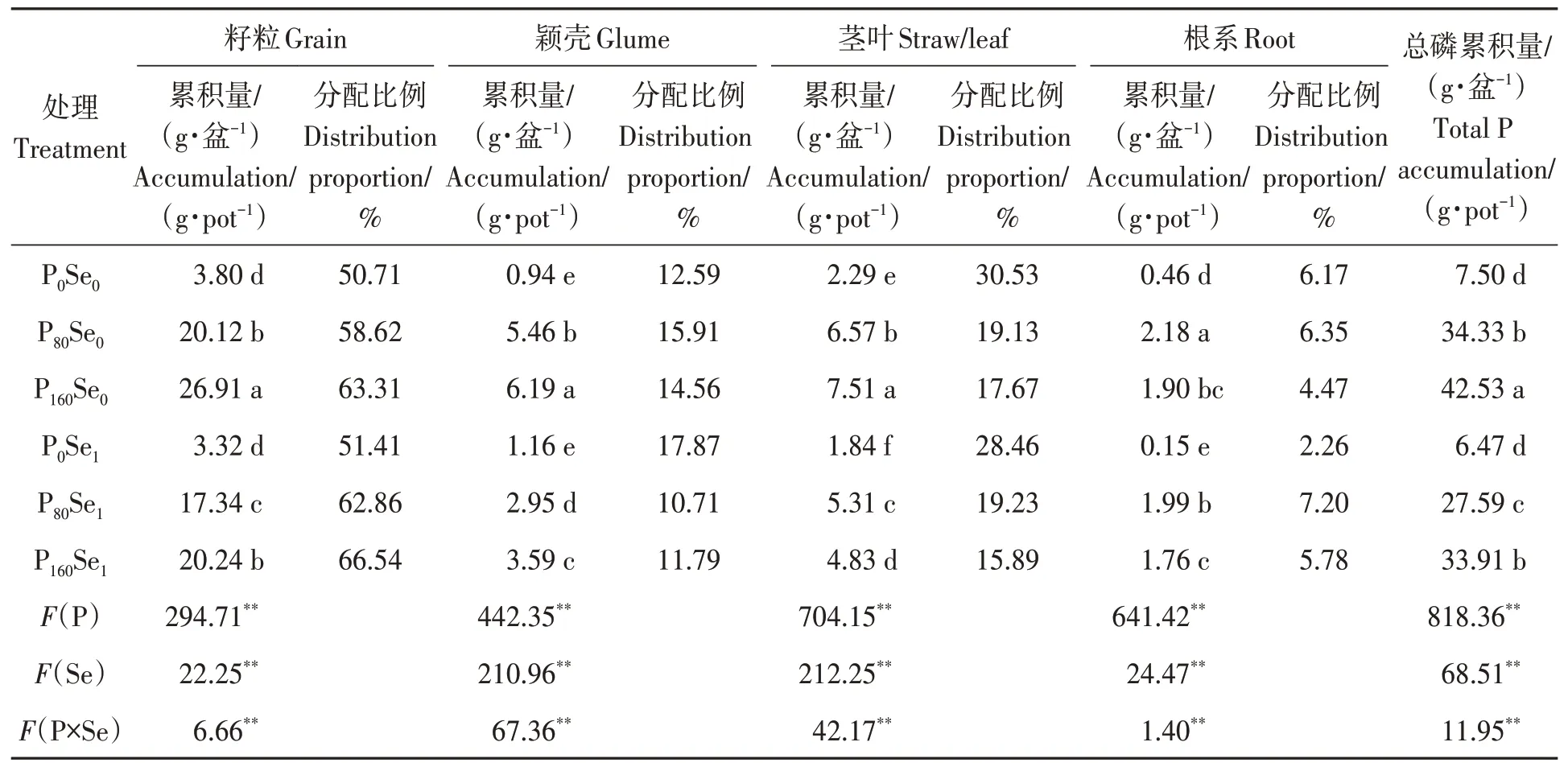
表4 磷硒配施下冬小麦磷累积量Table 4 P accumulation amount of winter wheat under phosphorus and selenium combined application
2.5 磷硒配施对冬小麦植株各部位硒含量及硒迁移系数的影响
由表5 可知,在Se1水平下‘郑麦379’各器官硒含量依次为根系>籽粒>颖壳>茎叶。在3 个磷水平下,施硒均显著提高了各器官硒含量。在Se1水平下,施磷显著降低了‘郑麦379’根系、茎叶、颖壳和籽粒硒含量,其降幅为17.63%~62.45%。根系、茎叶、颖壳和籽粒硒含量的最大值均出现在P0Se1处理。与Se0处理相比,Se1处理降低了P80和P160水平下根系向茎叶的硒迁移系数,其降幅分别为31.58%和14.29%,但显著增加了颖壳向籽粒的硒迁移系数,其增幅分别为181.93% 和118.52%,说明磷硒配施改变硒在冬小麦中的转运,促进硒从颖壳向籽粒的迁移。
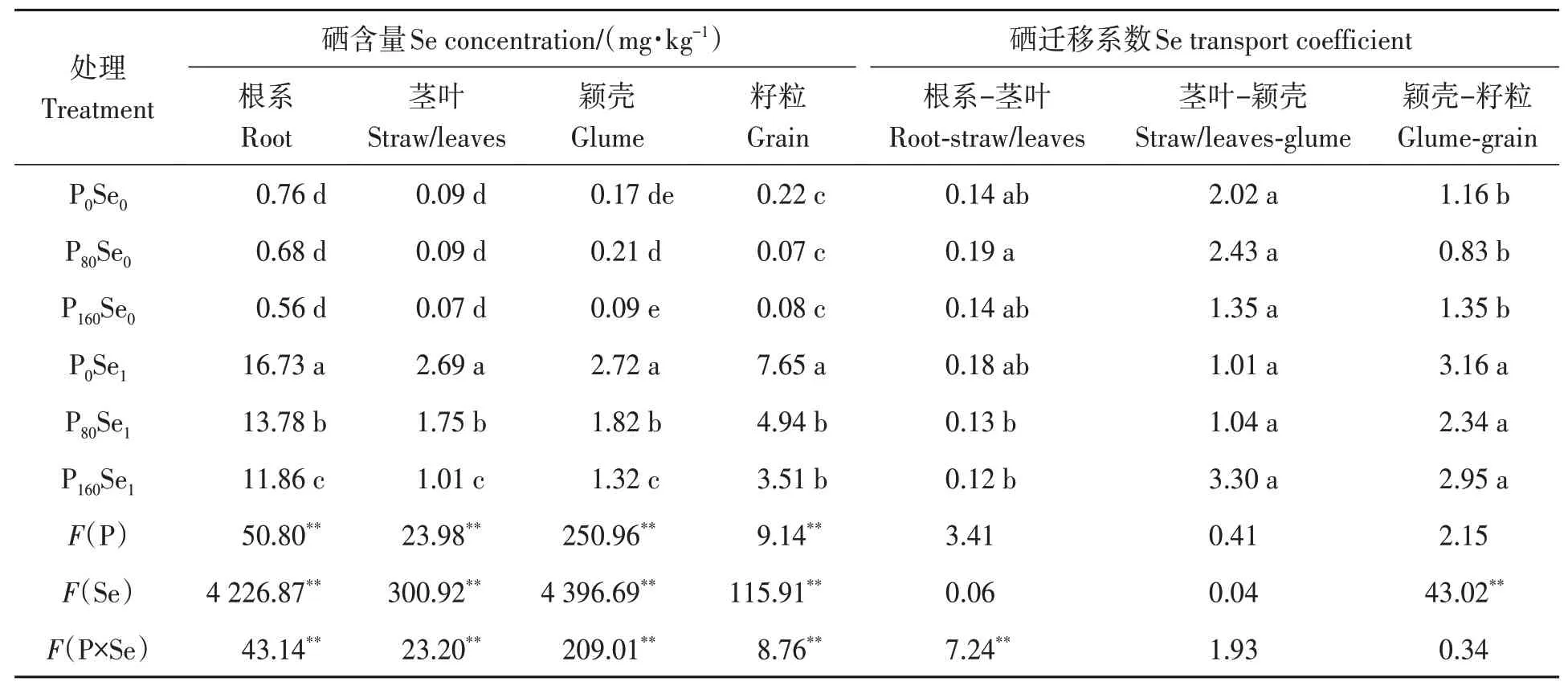
表5 磷硒配施下冬小麦各部位硒含量及迁移系数Table 5 Se content and migration coefficient of winter wheat under phosphorus and selenium combined application
2.6 磷硒配施对冬小麦植株各部位硒累积量和分配比例的影响
由表6可知,‘郑麦379’成熟期各器官硒累积量依次为籽粒>根系>茎叶>颖壳。与Se0处理相比,施硒显著增加了‘郑麦379’籽粒、颖壳、茎叶、根系及总硒累积量。在Se0水平下,施磷降低了茎叶和根系硒累积量的分配比例,其降幅为6.35%~35.18%,增加了籽粒硒累积量的分配比例,其增幅为21.67%~37.16%。在Se1水平下,施磷降低了颖壳和茎叶硒累积量的分配比例,其降幅为16.00%~48.75%,增加了根系硒累积量的分配比例,其增幅为30.06%~55.56%。颖壳、茎叶、根系和整株硒累积量的最大值均出现在P80Se1处理,籽粒硒累积量分配比例的最大值出现在P160Se1处理。

表6 磷硒配施下冬小麦硒累积量Table 6 Se accumulation amount of winter wheat under phosphorus and selenium combined application
2.7 磷硒配施对‘郑麦379’籽粒硒形态的影响
由表7 可知,在Se0水平下‘郑麦379’小麦籽粒中未检测到硒代胱氨酸、硒代蛋氨酸和无机硒。在施硒条件下,籽粒中硒形态含量依次为硒代蛋氨酸>无机硒>硒代胱氨酸。在Se1水平下,施磷降低了冬小麦籽粒中硒代蛋氨酸和无机硒含量,其降幅分别为64.34%~66.96%和16.85%~34.83%。硒代胱氨酸仅在P0Se1处理检测到,其含量为0.70 mg·kg-1。籽粒硒代蛋氨酸含量最大值出现在P0Se1处理,最小值出现在P160Se1处理,前者为后者的3.03倍。
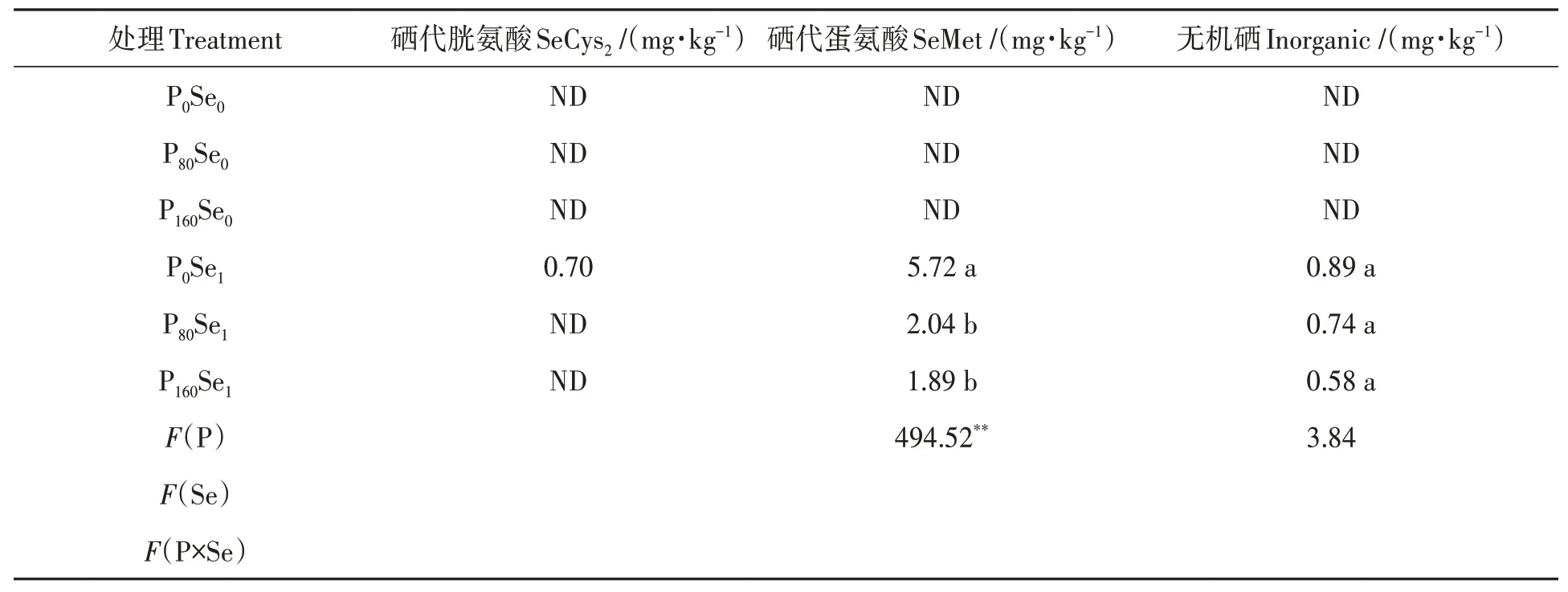
表7 磷硒配施下冬小麦籽粒硒形态Table 7 Grain Se species of winter wheat under phosphorus and selenium combined application
3 讨论
3.1 磷硒配施可显著提高冬小麦产量
磷肥在提高作物产量方面具有重要作用,硒元素能够参与植物体内的能量代谢及光合作用,调节植物生长进而促进作物高产[31-32]。本研究中,冬小麦生物量和籽粒产量均随磷肥用量的增加而增加。闫金垚等[33]发现,增施磷肥可促进不同品种水稻显著增产,这与本研究结果相似。施硒1 mg·kg-1显著增加了P80水平下冬小麦生物量及籽粒产量,降低了P160水平下冬小麦生物量及籽粒产量,表明施硒对冬小麦生物量和籽粒产量的影响受磷肥施用水平的影响。Nie 等[8]研究发现,在不同硒水平下,冬小麦籽粒产量随施磷水平的增加呈先增加后降低的趋势,这与本研究结果一致,可能是因为在正常磷水平(P80)条件下,配施一定量的亚硒酸盐能提高植株光合作用进而促进增产[34-35];而在高磷水平(P160)条件下,植物吸收磷酸盐与亚硒酸盐共用磷转运蛋白通道[36],施硒显著降低了P160水平下小麦各部位磷含量及累积量,抑制了冬小麦根系对磷的吸收进而导致磷对植株的生长的促进作用减弱。赵文龙等[12]指出,在硒含量≤5 mg·kg-1时,施磷均能显著增加小白菜地上部和地下部生物量,促进小白菜的生长,这与本研究结果不太一致,可能与作物种类有关。
3.2 磷硒配施降低冬小麦各器官硒含量,但提高硒累积量及硒从颖壳向籽粒的迁移
研究表明,土壤和叶面施硒均可提高植株叶片和水稻根、茎、叶及籽粒硒含量[37-38]。本研究中,与Se0处理相比,施用亚硒酸钠可显著提高冬小麦根、茎叶、颖壳及籽粒硒含量,与史丽娟等[39]对高粱的研究结果相似。在土壤-植物系统中,磷酸盐与亚硒酸盐关系复杂[12,17]。在土壤中由于磷酸盐与亚硒酸盐竞争吸附位点,增加磷的供应水平可减少土壤表面对Se 的吸附,提高土壤中有效硒含量;在植物体中,由于磷酸盐与亚硒酸盐用相同的转运通道,磷肥的施用降低了磷转运蛋白的表达量,二者存在竞争吸收作用[25]。本研究表明,在Se1水平下,施磷显著降低了冬小麦各部位硒含量。Wang 等[15]在水培条件下发现,水稻地上部硒含量与施磷水平呈显著负相关,这一方面可能是因为施磷降低了磷转运蛋白的表达量进而抑制冬小麦对硒的吸收[25,36],另一方面可能是由于施磷显著增加了植株生物量进而引起的生物稀释效应。赵文龙等[12]得出相似结论,施用磷肥促进小白菜生长产生生物稀释作用,进而降低小白菜各部位硒含量。安军妹等[16]在田间试验条件下发现,亚硒酸盐(7.2 kg·hm-2)与磷酸一铵(180 kg·hm-2)配施能促进小麦对硒的吸收,显著增加籽粒硒含量,这与本研究结果不一致,这可能与作物品种、磷肥形态和磷硒施用水平有关。本试验在Se1水平下,P80和P160处理显著增加冬小麦各部位硒累积量,且在P80Se1处理下达到最大值。磷肥的施用一方面会提高植物蒸腾作用,增强植物对硒的吸收能力;另一方面由于磷酸根和亚硒酸根在土壤中竞争吸附位点,施磷提高硒的有效性,促进植株对硒的吸收[17-18]。Liu 等[14]也发现,适宜的磷水平能促进水稻地上部对硒的累积,但过量施磷却抑制硒的累积。潘丽萍等[40]研究表明,甘薯中硒的分配规律表现为叶片>茎叶>块根。本研究中,‘郑麦379’成熟期各器官硒累积分布依次为籽粒>根系>茎叶>颖壳,且磷硒配施处理促进了颖壳向籽粒的迁移,进而增加了硒在籽粒中的占比,这与刘红恩等[5]研究结果一致。
3.3 施硒增加籽粒有机硒含量
植物体内存在多种含硒化合物,包括蛋白硒、硒代氨基酸、硒多糖以及硒核酸等[41-43],而大多数研究集中在硒代半胱氨酸、硒代胱氨酸和硒代蛋氨酸[15,44]。研究表明,施硒显著增加小麦籽粒中总硒和有机硒含量,且含量最多的有机硒是硒代蛋氨酸[45]。卢鹏飞等[46]研究表明,叶面喷施纳米硒、有机硒和无机硒肥均可显著提高硒代氨基酸和无机硒含量,且喷施无机硒亚硒酸钠时籽粒中无机硒含量最高。本研究中,在Se0水平下未检测到硒代蛋氨酸、硒代胱氨酸和无机硒,可能是因为在不施硒时冬小麦籽粒中硒含量较低。在Se1水平下,冬小麦籽粒硒形态含量依次为硒代蛋氨酸>无机硒>硒代胱氨酸,这与前人研究结果不太一致[46-47],可能与外源硒肥形态和施硒方式有关。Wang 等[15]在水培条件下研究结果表明,水稻体内硒代蛋氨酸含量占比最高,且磷酸盐的施用显著影响亚硒酸盐在水稻体内的代谢。本研究表明,在Se1水平下施磷显著降低籽粒中硒代蛋氨酸和无机硒含量,一方面是因为施磷抑制了冬小麦对硒的吸收[25];另一方面可能是因为磷参与植物体内的蛋白质合成,进而影响植物体内硒的代谢[23]。目前,磷肥与硒肥配施影响冬小麦籽粒硒的代谢机制尚未清楚,仍需进一步研究。
猜你喜欢
麦类作物学报(2022年1期)2022-03-01
粮食加工(2020年4期)2020-02-28
农民致富之友(2019年33期)2019-12-20
福建农林大学学报(自然科学版)(2019年1期)2019-01-25
植物保护(2017年1期)2017-02-13
西南农业学报(2016年6期)2016-04-16
中学生(2015年4期)2015-08-31
新疆农垦科技(2014年5期)2014-02-28
新疆农垦科技(2014年2期)2014-02-28
