
γ-氨基丁酸和光照对香稻稻米品质和干物质积累的影响
2023-11-21 03:44张楠麦一鸣王在满谢文君段延廖德榕梁殷悦莫钊文
作物研究 2023年5期
张楠,麦一鸣,王在满,谢文君,段延,廖德榕,梁殷悦,莫钊文*
(1 华南农业大学农学院/华南农业大学亚热带农业生物资源保护与利用国家重点实验室/农业部华南地区作物栽培科学观测试验站/广州市丝苗香米科学与技术重点实验室,广东 广州 510642; 2 华南农业大学工程学院/华南农业大学南方农业机械与装备关键技术教育部重点实验室,广东 广州 510642)
中国是世界上主要的水稻生产国之一,其水稻种植面积和产量居世界前列[1]。 香稻具有独特的芳香气味和较高的营养价值[2],其价格是普通稻米的数倍。 随着人们生活水平的不断提高,香米的消费量也在逐年增加,其生产和研究也受到了越来越广泛的关注[3]。 γ-氨基丁酸是一种重要的非蛋白质氨基酸,参与了植物的逆境胁迫响应、信号传导和碳氮平衡等多种生理生化反应[4-7]。 光照是影响水稻生长的重要环境因子之一,对水稻的产量和品质形成也具有影响[8],弱光或遮阴条件会导致水稻的产量降低[9-12]。 任万军等[9]指出,遮光条件下水稻产量降低与弱光影响了水稻的干物质积累和运转有关。 莫钊文等[13]研究表明,在灌浆成熟期进行遮阴处理对水稻的产量和品质具有显著影响。 谢文君[14]研究发现,喷施γ-氨基丁酸可调节弱光或者遮阴对香稻产量的影响。 但γ-氨基丁酸是否可以调控弱光下的稻米品质以及香稻的稻米品质与干物质积累、分配之间是否存在联系均尚不清楚。 因此,本试验以3 个香稻品种为试验材料,分析弱光处理下外源喷施γ-氨基丁酸对香稻稻米品质和干物质积累的影响,以期为提高弱光下香稻的稻米品质提供一定的参考和依据。
1 材料与方法
1.1 试验地概况
试验于2017 年晚稻季在华南农业大学教学实验农场进行。 试验田土壤pH 为6.34,含有机质21.74 g/kg、全氮1.70 g/kg、全磷1.30 g/kg、全钾0.10 g/kg。
1.2 供试材料
供试的香稻品种为Basmati、玉香油占和云粳优14,均由华南农业大学农学院水稻研究室提供。
1.3 试验设计
大田试验采用随机区组设计,以常规光照+不喷施γ-氨基丁酸的处理为对照(CK),设置了弱光(光照强度为常规光照的67%)+不喷施γ-氨基丁酸处理(T1)和弱光+喷施γ-氨基丁酸处理(T2)。T2 处理于水稻抽穗初期喷施质量浓度为250 mg/L的γ-氨基丁酸,喷施量100 mL/m2,CK 和T1 处理喷施等量的清水。 喷施结束后,T1 和T2 处理小区立即使用黑色塑料遮阳网进行遮光,遮光后的光照强度为常规光照强度的67%,遮光处理15 d 后去除遮阳网[14-15]。 供试水稻品种种子经浸种后播种于毯状育秧软盘,待秧苗长至3 叶1 心期,采用东方井关牌搭乘坐式高速插秧机插秧,插植规格为30 cm×14 cm。 每个处理3 次重复,小区面积为15 m2。 移栽前施用尿素、过磷酸钙和氯化钾作基肥,施肥量分别为纯氮180 kg/hm2、五氧化二磷90 kg/hm2、氧化钾150 kg/hm2。 水分和病虫草害的控制按照常规管理方法进行。
1.4 测定指标与方法
1.4.1 稻米品质测定
于成熟期收获水稻稻谷,脱粒后自然晾晒,在常温下储存3 个月后进行稻米加工品质、外观品质和营养食用品质等指标的测定[13]。
1.4.1.1 加工品质测定
加工品质指标主要包括糙米率、精米率和整精米率。 使用FC2K 型砻谷机对稻谷进行脱壳处理,再筛选出残留的谷壳和未脱壳的稻谷,称取糙米和未脱壳稻谷的质量,计算糙米率;使用JNMJ6 型检验碾米机碾磨糙米60 s,去除糠层,称取精米的质量,计算精米率;使用两层圆孔网筛将精米过筛(孔径分别为1.8 mm 与2.0 mm),采用人工将碎米挑出后再称取整精米质量,计算整精米率。
1.4.1.2 外观品质测定
使用万深SC-E 大米外观品质检测仪对整精米进行扫描分析,测定稻米的长宽比、垩白粒率和垩白度。
1.4.1.3 营养食用品质测定
使用FOSS 近红外谷物品质分析仪测定稻米的蛋白质含量、直链淀粉含量和碱消值。
1.4.2 光合特性和干物质测定
在成熟期,于晴朗的上午9:00—11:00,各小区选取代表性水稻植株4 穴,使用美国LI-COR 公司生产的便携式光合测定仪LI-6400XT 测定剑叶的蒸腾速率、气孔导度、胞间CO2浓度和净光合速率[16]。
分别在水稻抽穗后15 d 和成熟期,各小区选取代表性的水稻植株6 穴,带回实验室。 将植株地上部分别按照茎鞘、叶片和穗3 个部位剪开分装,放入烘箱中,在105 ℃条件下杀青30 min,然后在80 ℃条件下烘干至恒重,待样品冷却至室温后,用电子天平称量并记录干质量。
1.5 数据处理与分析
采用Microsoft Office 2013 进行数据整理;采用Statistix 8.0 软件进行数据统计分析,用最小极差显著法(LSD)进行多重比较。
2 结果与分析
2.1 对香稻稻米加工品质的影响
由表1 可知,与CK 相比,香稻品种Basmati T1、T2 处理的糙米率分别显著降低了0.88%和0.48%,T1 处理的整精米率显著降低了4.44%;玉香油占T1、T2 处理的糙米率分别显著提高了1.15%和1.76%,整精米率分别显著下降7.47%和7.83%;云粳优14 T1 处理的糙米率、精米率和整精米率分别显著降低了9.37%、9.58%和9.85%,T2 处理则分别显著降低了2.08%、2.81%和3.43%。 且云粳优14 T2 处理的糙米率、精米率和整精米率分别较T1处理显著提高了8.04%、7.48%和7.12%。 可见,喷施γ-氨基丁酸可以在一定程度上缓解弱光处理对整精米率的不利影响。

表1 不同处理下不同品种香稻稻米的加工品质Table 1 Processing quality of different fragrant rice varieties under different treatments %
2.2 对香稻稻米外观品质的影响
由表2 可知,对于Basmati,T1 处理稻米的长宽比较CK 显著降低了10.28%;T2 处理的垩白粒率和垩白度分别较CK 显著提高了429.41% 和694.34%;T2 处理的垩白粒率、垩白度和长宽比分别较T1 处理显著提高了291.3%、557.81% 和10.13%。 对于玉香油占,T1 处理的垩白粒率和垩白度分别较CK 显著提高了282.86%和437.50%;T2 处理的长宽比较CK 显著增加了10.17%,而垩白粒率和垩白度没有显著差异;但T2 处理的垩白粒率和垩白度分别较T1 处理显著降低了77.61%和81.40%,长宽比则显著提高了14.54%。 对于云粳优14,TI 处理的垩白粒率和垩白度分别较CK 显著增加了156.34%和290.19%,长宽比则显著下降2.52%;T2 处理的垩白粒率和垩白度分别较CK 显著增加了62.79%和147.72%,长宽比显著降低了2.52%;T2 处理的垩白粒率和垩白度分别较T1 显著降低了36.21%和49.46%。 可见,在弱光处理下外源喷施γ-氨基丁酸可以明显改善玉香油占和云粳优14 的稻米外观品质。

表2 不同处理下不同品种香稻稻米的外观品质Table 2 Appearance quality of different fragrant rice varieties under different treatments
2.3 对香稻稻米营养食用品质的影响
由表3 可知,不同处理对香稻稻米的直链淀粉含量无显著影响;除云粳优14 外,各处理香米的碱消值也无显著差异。 对于Basmati,蛋白质含量为T1、T2 处理分别较CK 显著提高了10.40% 和22.88%,T2 处理较T1 处理显著提高了11.31%。 对于玉香油占,T1 和T2 处理的蛋白质含量均显著高于CK,分别提高了21.53%和8.61%;而T2 处理蛋白质含量显著低于T1。 对于云粳优14,蛋白质含量为T1、T2 处理均显著高于CK,且T1 显著高于T2;碱消值为T1 处理显著低于T2 和CK,T2 与CK 则无显著差异。 综上所述,弱光处理和外源喷施γ-氨基丁酸均显著影响了香稻稻米的蛋白质含量。

表3 不同处理下不同品种香稻稻米的营养食用品质Table 3 Nutritional and eating quality of different fragrant rice varieties under different treatments
2.4 对香稻成熟期叶片光合特性的影响
由表4 可知,对于Basmati,与CK 相比,T1 处理显著降低了蒸腾速率、气孔导度、胞间CO2浓度和净光合速率,分别降低了18.50%、36.67%、13.76%和25.60%;T2 处理显著降低了气孔导度和胞间CO2浓度,分别显著降低了23.33%和11.70%;T2处理的蒸腾速率、气孔导度、胞间CO2浓度和净光合速率较T1 处理略有提高,但无显著差异。 对于玉香油占,与CK 相比,T1 处理显著降低了气孔导度和胞间CO2浓度,分别降低了32.00%和17.22%;T2 处理显著降低了胞间CO2浓度;T2 处理的蒸腾速率、气孔导度、胞间CO2浓度和净光合速率略高于T1 处理,但差异不显著。 对于云粳优14,与CK相比,T1 和T2 处理均显著降低了气孔导度和胞间CO2浓度;T2 处理的蒸腾速率、气孔导度、胞间CO2浓度和净光合速率均略高于T1 处理,但差异不显著。 由此可见,弱光处理下外源喷施γ-氨基丁酸可以改善香稻成熟期叶片的光合特性指标。

表4 不同处理下不同品种香稻成熟期叶片光合特性Table 4 Leaf photosynthetic characteristics of different fragrant rice varieties during maturity period under different treatments
2.5 对香稻干物质积累的影响
由表5 可知,香稻抽穗后15 d,与CK 相比,Basmati T1 和T2 处理均显著降低了茎鞘、叶片和穗的干质量;玉香油占T1 处理显著降低了穗干质量,T2 处理的茎鞘和穗干质量则分别显著降低了30.50%和39.23%;云粳优14 T1 处理显著降低了茎鞘和穗干质量,T2 处理显著降低了茎鞘和穗干质量,对叶片干质量无显著影响,且T2 处理的穗干质量显著高于T1 处理。 由此可见,外源喷施γ-氨基丁酸可以提高弱光下香稻成熟期穗干物质的积累量,且对玉香油占和云粳优14 的效果更好。
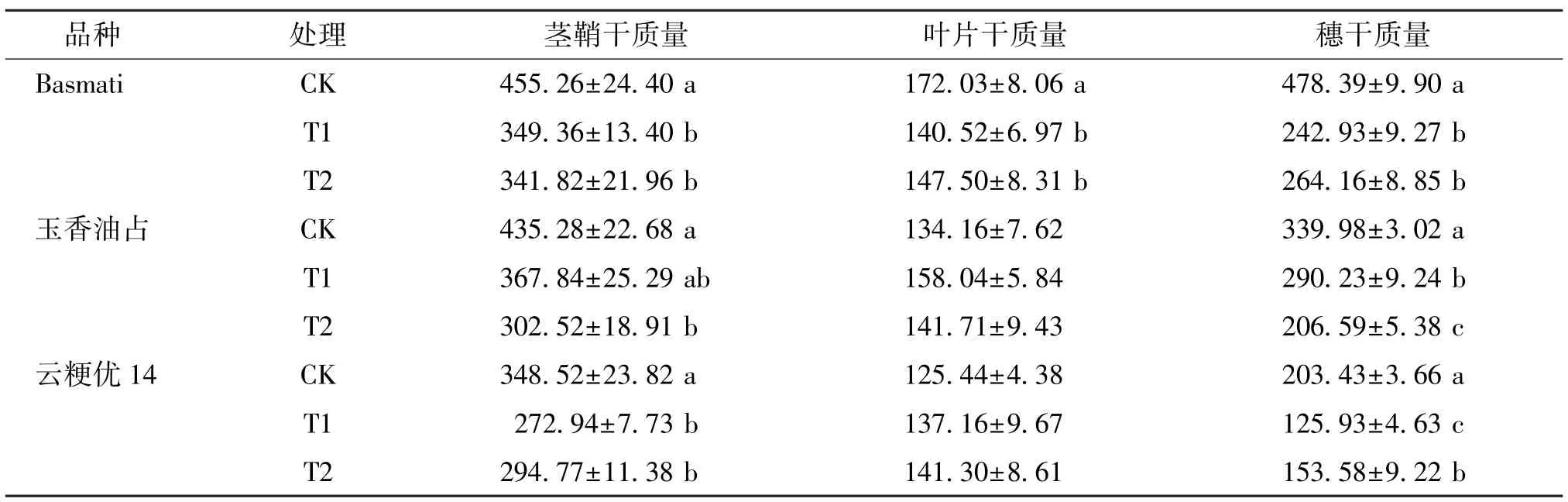
表5 不同处理下不同品种香稻抽穗后15 d 的干物质积累量Table 5 Dry matter accumulation of different fragrant rice varieties at 15 days after heading under different treatments g·m-2
由表6 可知,成熟期,与CK 相比,Basmati T1 处理的茎鞘和穗干质量分别显著降低了21.14%和33.95%,T2 处理的茎鞘和穗干质量分别显著降低了17.74%和26.79%;且T2 处理的穗干质量较T1处理显著增加了10.83%。 对于玉香油占,与CK 处理相比,T1 处理的叶片干质量显著增加了21.53%,穗干质量显著减少12.05%,各处理的茎鞘干质量则无显著差异;T2 处理的叶片干质量显著增加了28.46%,茎鞘和穗干质量则无显著差异。 对于云粳优14,与CK 相比,T1 处理的茎鞘和叶片干质量分别显著增加了9.98%和38.84%,穗干质量显著降低了30.58%;T2 处理的叶片干质量显著增加了19.40%,穗干质量显著下降了20.88%,茎鞘干质量则无显著差异。 由此可见,外源喷施γ-氨基丁酸可以增加弱光下香稻成熟期的穗干物质积累量,其中Basmati 和玉香油占的增加效果尤为显著。
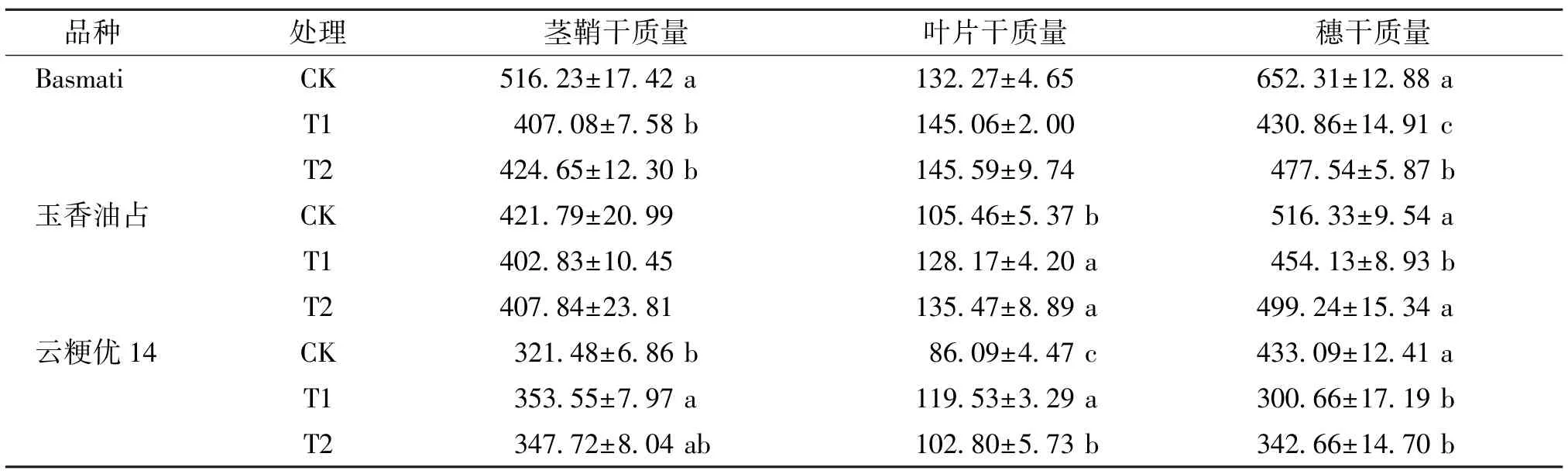
表6 不同处理下不同品种香稻成熟期的干物质积累量Table 6 Dry matter accumulation of different fragrant rice varieties at maturity under different treatments g·m-2
2.6 对香稻不同器官干物质比例的影响
由表7 可知,对于Basmati,与CK 相比,T1 处理的茎鞘和叶片干物质比例显著提高,分别提高了15.96%和23.39%,穗干物质比例则显著降低;T2处理茎鞘和叶片干物质比例分别显著提高了10.04%和26.16%,穗干物质比例则显著降低;与T1 处理相比,T2 处理的叶片和穗干物质比例略有提高,但无显著差异。 对于玉香油占,T1、T2 处理叶片干物质比例显著高于T1;但T2 处理的穗干物质比例显著低于CK 与T1 处理。 对于云粳优14,与CK 处理相比,T1、T2 处理叶片干物质比例显著增加,穗干物质比例则显著降低。 可见,外源喷施γ-氨基丁酸影响了香稻品种抽穗后15 d 不同器官的干物质比例。

表7 不同处理下不同品种香稻抽穗后15 d 不同器官干物质比例Table 7 Dry matter ratio of different organs 15 days after heading of different fragrant rice varieties under different treatments %
由表8 可知,对于Basmati,T1 和T2 处理均显著影响成熟期叶片和穗干物质比例,相较于CK,T1处理的叶片干物质比例显著增加了45.33%,穗干物质比例显著降低了12.75%;T2 处理的叶片干物质比例显著增加了36.58%,穗干物质比例显著降低了9.09%。 对于玉香油占,与CK 处理相比,T1处理显著降低了成熟期茎鞘和穗干物质比例,显著增加了叶片干物质比例;T2 处理显著增加了叶片干物质比例,对茎鞘和穗干物质比例则无显著影响;且T2 处理的穗干物质比例较T1 处理显著提高了13.45%。 对于云粳优14,T1 和T2 处理均显著影响了成熟期叶片和穗干物质比例,与CK 处理相比,T1处理的茎鞘和叶片干物质比例分别显著提高了19.64%和51.03%,穗干物质比例则显著降低了24.76%;T2 处理的茎鞘和叶片干物质比例分别显著提高了14.73%和26.39%,穗干物质比例则显著降低了16.24%;与T1 相比,T2 处理的叶片干物质比例显著降低了16.31%,穗干物质比例则显著提高了11.33%。 因此,外源喷施γ-氨基丁酸可促进弱光条件下香稻成熟期穗干物质比例的提高。
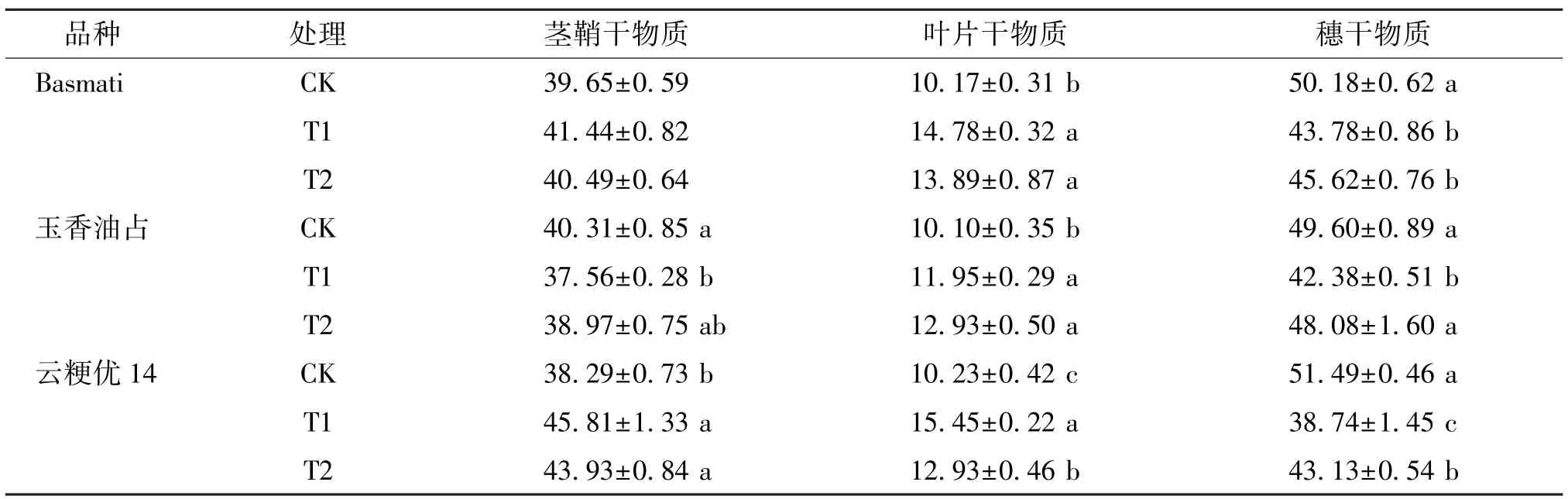
表8 不同处理下不同品种香稻成熟期不同器官干物质比例Table 8 Dry matter ratio of different organs of different fragrant rice varieties under different treatments %
3 讨论
任万军等[16]研究表明,始穗后遮阴处理会导致稻米垩白粒率和垩白度显著升高,且蛋白质含量也显著提高。 任万军等[9]发现,在水稻始穗期进行遮阴处理,水稻干物质积累随光强降低而减少,净光合速率的降低会引起始穗后的干物质积累量大幅度减少,干物质分配到穗的比例下降,分配到茎鞘和叶片的比例提高。 朱萍[17]研究表明,弱光条件下,水稻始穗后生产的干物质主要用于维持叶片和茎鞘生长,分配到穗部的比例大幅度减少,这可能与净光合速率有关[18]。 本研究结果表明,在弱光条件下,抽穗后15 d 和成熟期的穗部干物质积累量均呈下降趋势,而叶片干质量均呈现上升趋势。 此外,净光合速率存在品种差异,其中Basmati 品种呈现显著下降趋势。 前人[19]研究表明,弱光条件下稻米中的氨基酸含量要高于常规光照,总氨基酸含量与遮光程度成正比,遮光程度越大,则总氨基酸含量越多,因此表现为蛋白质含量增加。 此外,遮阴处理显著提高了香稻的蛋白质含量,在灌浆早期的遮阴则显著提高了垩白粒率和垩白度[20]。 γ-氨基丁酸是一种广泛参与植物胁迫反应的分子[21-23],能够提高植物的抗逆性与促进植物的生长[24-25]。 外源喷施γ-氨基丁酸可缓解弱光或遮阴对作物产生的不利影响[26]。 本研究发现,在弱光条件下外源喷施γ-氨基丁酸后,Basmati 和云粳优14 穗干物质积累量和干物质积累比例均呈现上升趋势。 此外,玉香油占在抽穗后15 d 穗部的干物质积累量呈现下降趋势,成熟期的穗部干物质积累量和干物质积累比例与抽穗后15 d 的穗部干物质积累比例均呈现增长趋势。 同时,3 个香稻品种的蛋白质含量在弱光处理下都明显提高,但直链淀粉含量则无明显变化。 弱光处理增加了3 个香稻品种稻米的垩白粒率和垩白度,而喷施γ-氨基丁酸则降低了玉香油占和云粳优14 的垩白粒率和垩白度,从而改善了香稻外观品质。 综上所述,弱光下外源喷施γ-氨基丁酸可改善香稻稻米的品质性状,这可能与干物质积累与分配有关。
4 结论
在成熟期和抽穗后15 d,弱光处理减少了穗干物质积累量,且会影响干物质积累和分配。 3 个香稻品种的蒸腾速率、气孔导度、胞间CO2浓度和净光合速率均呈下降的趋势,表明弱光对香稻成熟期光合特性产生了不利影响。 弱光处理下,3 个香稻品种稻米的加工、外观及营养食用品质均受到一定程度的不利影响,但外源喷施γ-氨基丁酸可在一定程度上改善香稻的稻米品质以及干物质积累与分配。 综合分析,γ-氨基丁酸可以改善弱光对香稻叶片光合作用特性产生的不利影响,进而改善稻米品质和优化干物质积累。
猜你喜欢
农业与技术(2024年7期)2024-04-19
干旱地区农业研究(2023年5期)2023-10-10
农业科技通讯(2023年1期)2023-02-12
上海农业学报(2022年6期)2023-01-14
福建稻麦科技(2022年3期)2022-09-14
浙江农业学报(2021年3期)2021-04-01
现代园艺(2017年23期)2018-01-18
麦类作物学报(2017年7期)2017-08-16
动物营养学报(2015年10期)2015-12-01
热带农业科学(2015年8期)2015-09-10
