
江苏淮北地区小麦种质资源的多性状评价及相关性分析
2023-11-19 06:42潘丽媛王永军李海军李丽丽孙苏阳
大麦与谷类科学 2023年5期
潘丽媛,王永军,李海军,李丽丽,侯 富,李 菁,孙苏阳
(江苏徐淮地区淮阴农业科学研究所/淮安市农业生物技术重点实验室/农业农村部淮河下游种质创制重点实验室,江苏 淮安 223001)
作物种质资源是农作物新品种选育和基础原始创新的基石,是保障国家粮食安全、推动农业绿色高效发展、维护生物多样性的战略资源[1]。小麦作为我国第三大粮食作物,对世界和我国的粮食安全具有重要的战略地位[2-3]。小麦在我国的种植范围广泛,籽粒中含有淀粉、蛋白质、脂肪、矿物质、钙、铁、硫胺素、核黄素和维生素A 等多种物质,营养价值较高,其独特的面筋特性可以用来制作多种面食,是全球35%~40%人口的主食[4]。
黄淮麦区作为中国小麦的主产区,该区域的小麦生产为我国的粮食安全提供了重要保障[5]。小麦种质资源作为新品种选育的基础,分析其农艺性状及各性状之间的相关性对小麦遗传改良具有重要作用[6-8]。农艺性状能够直观反映种质资源的优劣,是否符合育种目标,对小麦种质资源和品种农艺性状的考察和评价至今仍然是一项不可替代的内容[9]。前人关于小麦种质资源的遗传多样性研究内容较多,特别是小麦株高、分蘖数等常规农艺性状方面[10]。育种实践上,通常是对小麦种质资源进行农艺性状的变异分析、相关性及聚类分析筛选优异种质资源,提高育种水平和效率[11-12]。前人分析评价了105 份黄淮麦区小麦的农艺及品质性状,发现这些种质资源的遗传差异较小,筛选出4 份综合性状较好的材料可作为高产优质小麦育种亲本[13]。张会芳等分析了黄淮麦区不同来源地育成小麦品种性状多样性后发现,不同来源地小麦的前6 个主成分累积贡献率较小,综合得分均表现为强筋小麦大于中筋小麦,并且北片小麦的丰富度和均匀性均高于南片麦区[14]。李晶等对16 份俄罗斯小麦种质资源的21 个性状进行遗传多样性分析,结果表明其表型变异丰富,可为冬小麦品质育种提供新来源[15]。但综合来看,针对小麦种质资源在江苏淮北麦区的表现分析较少,且对于现有育成品种缺乏各性状的年代差异分析。
目前黄淮麦区小麦种质资源研究的遗传基础狭窄,本研究以地方品种、育种创制中间材料和育成品种共1 098 份种质资源为研究对象,分析其生育期、株型、籽粒及品质等17 个相关性状的变异,并进行各个性状间的相关性分析、聚类分析。对育成品种及种质材料的各性状进行不同年代的差异分析,分析小麦选育过程中的年代差异,为小麦新品种选育和遗传基础研究提供参考依据。
1 材料与方法
1.1 供试材料
本试验共选取1 098 份小麦种质材料,包含不同年代育成的924 份品种(来源于11 个省份和国内外地方品种与育种创制中间材料等)及174 份收藏的地理来源不明的种质材料(表1)。近年育种水平的提升和品种参试途径的丰富,使得育成品种/品系的数目伴随年代逐步增加。
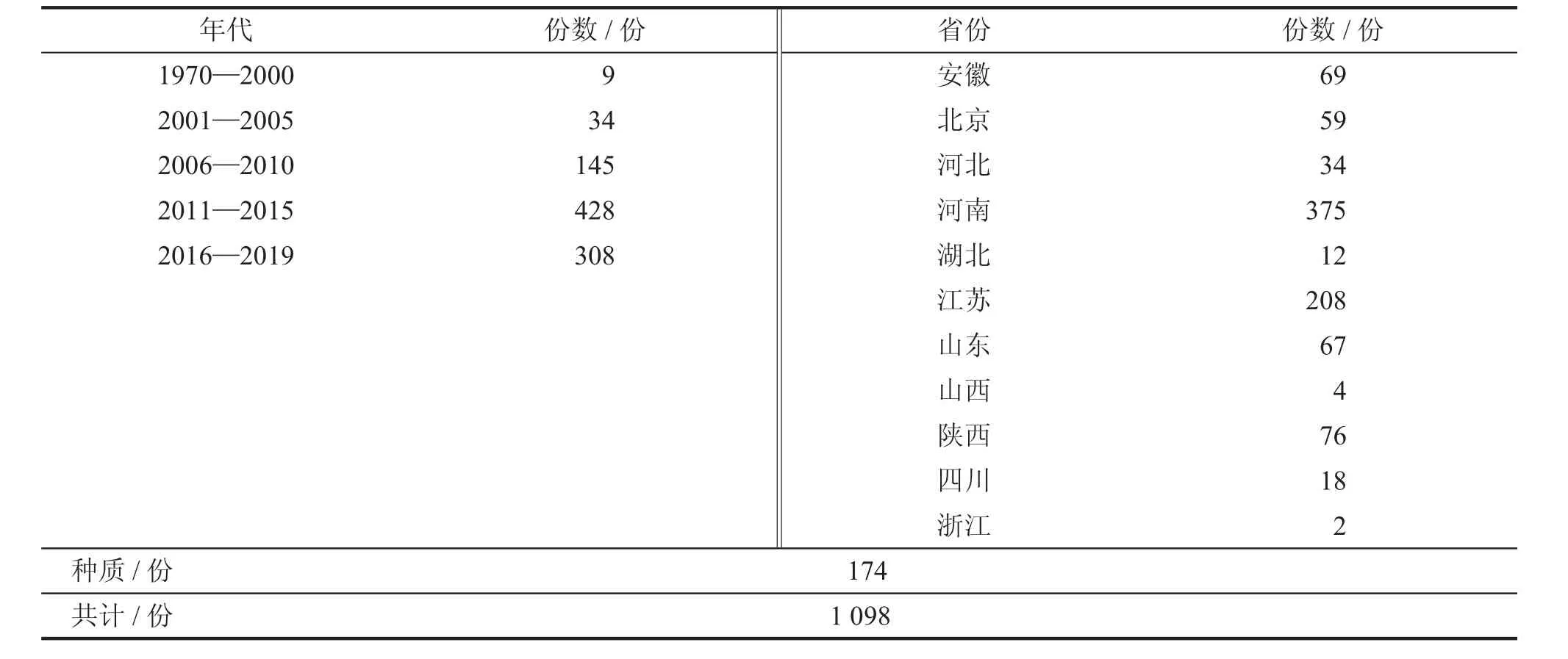
表1 小麦种质资源群体的概况
1.2 试验设计
试验材料于2018 年10 月23 日秋播种植于江苏徐淮地区淮阴农业科学研究所淮安创新基地(119°04′E、33°53′N),前茬作物为玉米。每份材料种植2 行,行播,行长1.5 m,行间距25 cm。生长季进行常规田间管理。
1.3 性状测定方法
田间调查生育期相关性状,包括抽穗期(每行50%以上麦穗顶部小穗露出叶鞘)、扬花期(每行50%以上植株开始扬花);至成熟期调查株型相关性状,包括株高、穗长和穗下节长度。收获2 个月后取适量籽粒,利用波通DA7250 近红外品质分析仪对籽粒进行品质性状测定,包括水分含量、硬度指数、容重、吸水率、粗蛋白、湿面筋、出粉率、延伸度、拉伸面积、形成时间、稳定时间、沉降值共12 个籽粒相关性状。
1.4 统计分析
采用Excel 2016、SAS 软件(v9.4)、R 语言(v4.0.2)进行数据分析,计算各个性状的最大值、最小值、极差、平均值、变异系数等参数,并进行相关性分析、单因素分析及聚类分析。
2 结果与分析
2.1 小麦种质资源群体各性状的表型概况
该研究的小麦种质资源群体共有1 098 份材料(表1),包含174 份地理来源不明的种质材料,924份育成品种。育成品种的年代跨度较大,分布在1970—2019 年,按不同年代划分表明育成品种材料呈现递增趋势,其中2010 年后育成的小麦品种占比较大,达到79.65%。按省份来看,这些材料来自我国小麦主产区的11 个省份,其中河南、江苏占比较高,分别达到34.15%和18.94%。分析不同地理来源的种质材料后发现,各个性状在不同地理来源之间的差异较小,其中仅穗长和穗下节长在不同省份之间差异较大,但细究可以看出,主要是因为冬、春小麦的差异。
由表2 可以看出,1 098 份小麦种质资源群体在淮安地区基本可以正常成熟,生育期相关性状中抽穗期变化范围为161.00~190.00 d,均值为170.08 d;扬花期的变化范围在162.00~202.00 d,均值为182.11 d。株型相关性状中株高变化范围为47.00~143.00 cm,均值为88.63 cm;穗长变化范围为4.00~22.00 cm,均值为10.90 cm;穗下节长变化范围为0~44.00 cm,均值为9.36 cm。籽粒相关性状中水分含量变化范围为11.08%~22.58%,均值为12.55%;硬度指数变化范围为41.68~81.31,均值为64.86;容重变化范围为725.40~841.58 g/L,均值为777.58 g/L;出粉率变化范围为27.58%~68.18%,均值为61.13%;吸水率变化范围为59.34%~89.62%,均值为75.63%。籽粒品质相关性状中粗蛋白含量的变化范围为9.54%~24.09%,均值为14.11%;湿面筋含量变化范围为20.02%~39.98%,均值为30.85%;延伸性变化范围为15.54~189.60 mm,均值为127.34 mm;拉伸面积变化范围为0 ~221.89 mm2,均值为69.15mm2;形成时间变化范围为0~5.61 min,均值为2.96 min;稳定时间变化范围为0~26.13 min,均值为6.25 min;沉降值变化范围为10.24~65.05 mL,均值为31.10 mL。该群体能够反映我国小麦主产区的育种进程和特点。

表2 小麦种质资源群体各性状的表型分析
不同类别相关性状的变异系数存在一定差异,生育期相关性状的变异系数较低,扬花期和抽穗期分别为1.98%和2.63%。株型相关性状变异系数的变化范围为14.06%~51.22%,籽粒相关性状变异系数的变化范围为1.69%~9.40%,品质相关性状变异系数的变化范围为12.14%~84.82%。
2.2 小麦种质资源群体多性状的相关性分析
对1 098 份小麦种质资源的17 个性状进行相关性分析,可以看出不同性状之间的相关性差异较大,相关系数变化范围为-0.32~0.98(图1)。生育期相关性状抽穗期和扬花期之间达到极显著的正相关,相关系数达到0.83(P<0.001)。株型相关性状之间的相关性差异较大,株高与穗下节长之间的相关性系数达到0.56(P<0.001),穗长与穗下节长之间的相关系数为0.12,未达到显著水平。籽粒相关性状中,硬度指数与吸水率相关性最高,达到0.71(P<0.001)。籽粒品质相关性状间的相关性系数均达到显著正相关(P<0.05),各性状之间的相关系数为0.34~0.98,其中:粗蛋白含量与湿面筋含量的相关性系数最高,为0.98;湿面筋含量与拉伸面积的相关性系数最低,为0.34。

图1 小麦种质资源群体各性状的相关性分析
不同类别性状间的相关性差异较大,其中株型相关性状与籽粒和籽粒品质相关性状之间的相关性较低,均未达到显著水平。但株型相关性状与生育期各性状均为负相关,相关系数为-0.28~-0.17,均达到显著水平(P<0.05)。籽粒相关性状中的硬度指数、容重和出粉率与籽粒品质相关性状之间的相关性较低,均未达到显著水平。粗蛋白含量与生育期、株型和籽粒相关性状之间相关性均未达到显著水平。以上结果表明,各类型性状内的相关性较不同类型性状间的相关性高,并且籽粒品质相关性状之间的相关性较高,而其与其他类型性状的相关性较低。
2.3 小麦种质资源群体的聚类分析
对1 098 份小麦种质资源的17 个性状进行聚类分析(图2),结果表明,这些种质资源在马氏距离500 处可聚为4 类,其中:I 类含有227 份材料,II 类含有215 份材料,III 类含有258 份材料,IV 类含有398 份材料。不同类别的材料个数较为均匀,表明该群体各个性状的差异较为显著,可作为很好的研究群体。
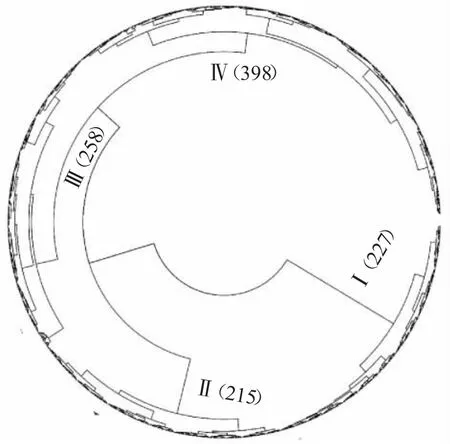
图2 小麦种质资源群体的聚类分析图
就育成品种而言,按照年代划分结果进一步分析发现,I 类中共含有178 个育成品种,其中2011—2015 年育成的为44 份,2016—2019 年育成的有107 份,占比分别达到24.72%和60.11%。II 类中含有181 个育成品种,其中占比最高的年代为2011—2015 年育成的品种,共有100 份,占比达到55.25%。III 类中含有220 个育成品种,各个年代材料的分布相对均匀,2006—2010 年占比为20.00%,2011—2015 年占比为45.91%,2015—2019 年占比为28.18%。IV 类中含有345 个育成品种,占比最高的为2011—2015 年育成的品种,达到53.33%。根据分类结果,可以看出2001—2010 年育成的品种较为均匀地分布在4 个类别中,表明这个年代育成的品种在本群体中的表型较为丰富。
2.4 不同年代小麦种质资源各性状的变化趋势
对不同类别的各性状不同年代的单因素分析表明,不同性状在各个年代间的表现存在一定的差异。生育期性状中抽穗期和扬花期在不同年代中表现一致,种质材料生育期较长,育成品种按年代由远及近有越来越长的趋势。株型性状中差异较为显著的是株高和穗下节长,由表3 可知,育成品种的株高降低,从2000 年前的99.56 cm 降至85.01 cm。穗下节长变短,从之前的13.22 cm 降至8.17 cm。籽粒性状中不同年代差异较大的是吸水率,随着年代的推移,吸水率由71.61%升至78.55%。籽粒品质各性状中年代差异较大的是沉降值,由30.93 mL 升至34.51 mL。
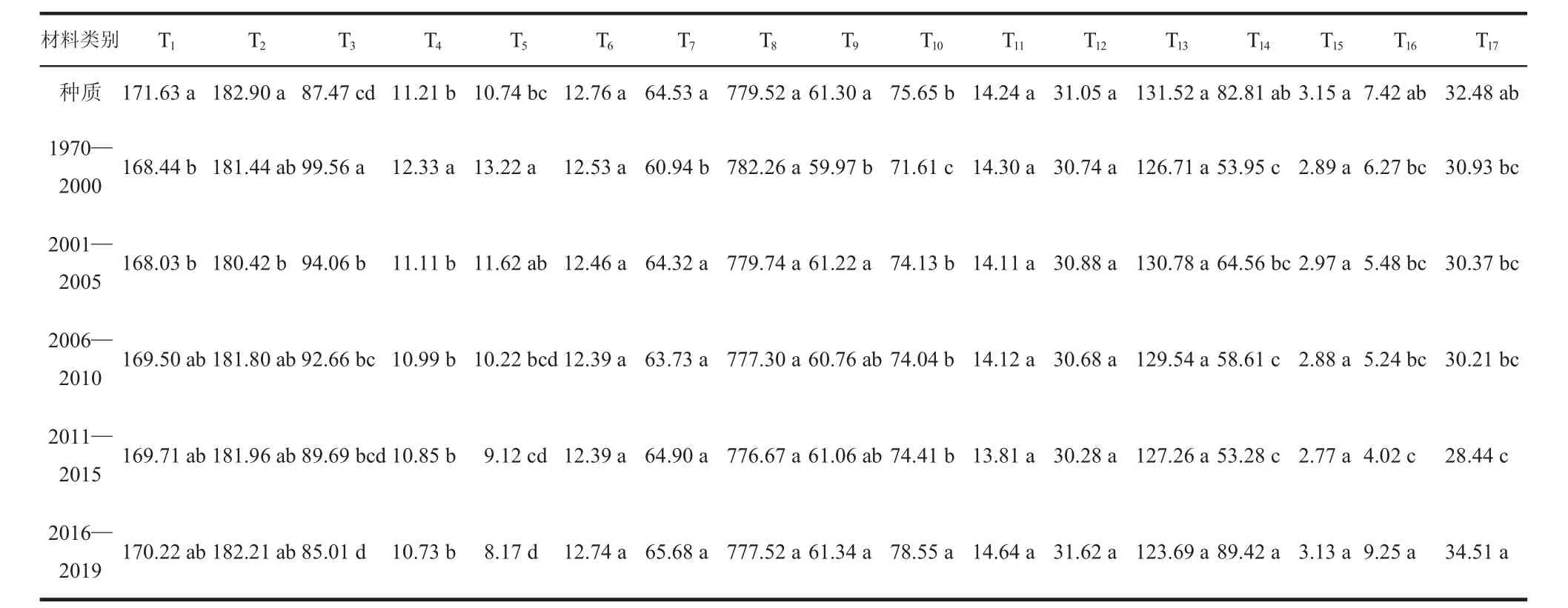
表3 不同年代小麦种质资源各性状的表现差异分析
3 讨论与结论
农业种质资源是支撑农业科技创新、现代种业发展的重要物质基础。对1 098 份小麦种质资源材料的农艺性状及品质相关性状的分析表明,该群体具有丰富的遗传多样性,其中株型相关性状和籽粒品质相关性状的遗传差异显著,遗传改良的潜力较大。特别是穗下节长和稳定时间具有较高的变异系数,表明该群体可作为这些性状的遗传基础研究群体。
相关性分析结果表明,不同类别性状内部的相关性差异较大,抽穗期和扬花期的相关性最高,达到0.83。株高与穗下节长和穗长达到显著正相关,但穗长和穗下节长与籽粒品质相关性状之间的相关性均不显著,表明该群体内穗长和穗下节长的遗传基础较为独立。张婷分析了黄淮麦区4 个省份的263 个小麦品种(系)种质资源遗传多样性,发现株高与穗长呈现正相关,相关系数达到0.37[16],与本文结果一致。籽粒品质相关性状的相关性较高,性状之间均达到显著水平,表明品质各个性状之间的遗传基础较为复杂,遗传改良难度较大。李楠楠等分析了我国四大冬小麦生产区品种的农艺性状,发现蛋白质含量等品质性状与产量呈现正相关的关系[17]。对不同地理来源小麦的分析发现,本研究群体各个性状在不同地理来源之间的差异较小,其中仅穗长和穗下节长在不同省份之间差异较大,原因可能在于冬、春小麦的差异。本研究群体中品质相关性状与其他性状之间的相关性较低,表明本研究种质资源群体内品质性状的遗传基础相对独立。
对这些种质资源群体的聚类分析表明,该群体可以分为4 类,种质材料在4 类中均有分布,育成品种的类群按照年代划分有所差异,其中:I 类中2016—2019 年的育成品种占比达到60.11%,表明近年育成品种的同质化现象较为严重,遗传基础狭窄,多局限在多数骨干亲本之间[18];但2001—2010 年间育成品种较为均匀地分布在4 个类别中,表明这些育成品种的表现较为丰富。
为进一步探究不同年代小麦种质材料特别是育成品种各个性状的变化趋势,通过单因素分析发现,这些性状在不同年代间变化较大的是株高、穗下节长、吸水率和沉降值,但品质重要性状粗蛋白和湿面筋含量并没有较大差异。表明在育种过程中,育种家较为重视矮秆、籽粒吸水率,但粗蛋白和湿面筋含量由于其自身遗传改良难度较大并未受到重视。
综上所述,本研究利用的1 098 份小麦种质资源在17 个性状上,遗传差异较大,多样性丰富,可作为遗传改良研究的基础群体。聚类分析表明,近年育成品种的同质化程度高,遗传基础较为狭窄,育种过程中对于株高和籽粒的吸水率等性状的选择较为重视。该群体的遗传基础差异大,变异丰富,下一步可与分子标记技术相结合,进一步挖掘该群体不同性状的遗传基础,全面解析该种质资源群体,为提高淮北麦区的育种效率和水平提供依据和材料基础。
猜你喜欢
今日农业(2022年13期)2022-09-15
现代畜牧科技(2021年4期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
河北农业科学(2018年2期)2018-07-26
中国麻业科学(2018年6期)2018-04-09
商洛学院学报(2017年2期)2017-05-17
高中生学习·高三版(2016年1期)2016-05-30
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
中学生数理化(高中版.高二数学)(2016年4期)2016-03-01
