
遮荫对川芎光合荧光特性的影响*
2023-11-15 08:26陈云飞马菁华欧明烛任启飞
贵州科学 2023年5期
陈云飞,刘 芳,马菁华,欧明烛,任启飞
(贵州省植物园,贵州 贵阳 550001)
川芎(LigusticumchuanxiongHort.)是伞形科多年生草本植物[1],主产四川,为川产道地药材,以地下根茎入药,药理作用广谱,为传统大宗药材,需求量大,经济价值极高。而光是植物生长过程中重要的环境因素之一,光强不足或过剩都会对植物的光合作用产生不利的影响[2],国内外普遍认为,植物的生长对遮荫响应十分敏感,由于遮荫弱光处理改变了植物生长的微环境[3],从而对植物的光合作用和能量传递等生理过程产生重要影响。前人对川芎进行了大量研究,但多集中在其药用成分上,有研究表明川芎中的有效成分川芎嗪具有一定的抗肿瘤作用,可减少肿瘤细胞同内皮细胞粘附抗凝、抗血小板聚集,抑制肿瘤的转移[4]。基于当前对川芎的研究大多集中在其有效成分及药理疗效上,遮荫对其光合荧光特性的影响未见报道,本研究旨在探明弱光环境对川芎光合作用及荧光特性的影响,摸清川芎对遮荫环境的动态响应情况,以期能更好地栽培利用,使有限的资源获得更大的收益。
1 材料与方法
1.1 试验材料
本试验中所用川芎苓种购置于四川省仁寿创新农业开发有限公司,经人工挑选,选择大小一致无病虫害的粗壮苓种茎节作为试验材料用于播种。
1.2 试验设计
于2021年8月17开始试验,采用黄泥土和营养基质2∶1比例配置栽种基质,整地起陇,分为4块试验样地,利用3针遮阳网进行遮荫处理,每块地遮阳棚长宽高为2 m×1.5 m×1.5 m,四周及顶部遮荫,距地面留出20 cm用于通风。一共设计4个光强梯度,CK为全光对照,C1遮荫度25%,C2遮荫度50%,C3遮荫度率75%。各个遮荫处理下播种30株,播种间距为30 cm,覆土3 cm左右。对其进行水肥除草管理及病虫害防治,保证生长发育正常进行,用于测定其苗期及第一次茎叶发生期的光合参数、荧光参数、叶绿素含量。
1.3 试验方法
1.3.1 光合参数的测定
选择在晴天上午的9∶00—11∶00点,用Li-6800便携式光合仪测定川芎植株的净光合速率(A)、气孔导度(gsw)、胞间CO2浓度(Ci)、蒸腾速率(E)。选择2 cm2叶室适配器,叶室温度为环境温度,约28 ℃,样品室CO2浓度400 μmol·mol-1,进入叶室的气体流速为500 μmol·s-1,叶室相对湿度在50%。各遮荫处理下选择3株长势相同的植株,高度一致的无病虫害的健康叶片为测量叶片,每株重复测量3次。
1.3.2 荧光参数的测定
选择在晴朗天气的上午9∶00—11∶00,使用OPTI-sciences公司OS5p+便携式脉冲调制叶绿素荧光仪测定叶绿素荧光参数,包括初始荧光Fo、最大荧光Fm、PSII最大光化学效率Fv/Fm、实际光化学效率Y(II)、电子传递速率ETR。用叶夹夹住植株健康叶片,以铺满叶夹为准,进行暗适应15 min后测定Fo、Fm、Fv/Fm、Y(II)、ETR。每个处理重复3次。
1.3.3 叶绿素的测定
取健康的供试川芎新鲜叶片0.2 g,洗净吸干剪碎,用95%乙醇黑暗中浸提24 h,期间摇匀3次,直至叶片完全脱色。利用岛津UV-1780紫外分光光度计分别在645 nm和663 nm处测定吸光度,分别计算叶绿素a、叶绿素b、总叶绿素含量。每个处理重复3次。
1.3.4 数据处理
采用EXCEL软件统计数据及初步处理,利用SPSS.26.0 软件对数据进行统计分析,采用最小显著差法(LSD)进行差异显著性检验,采用皮尔逊法进行相关性分析。
2 结果与分析
2.1 不同遮荫处理对川芎光合作用的影响
遮荫可影响植物的一系列生理过程从而间接地影响光合作用[5],其中最大净光合速率(A)、胞间CO2含量(Ci)、蒸腾速率(E)和气孔导度(gsw)可作为植物在遮荫环境下生长状况的响应评价指标[6],其中A的强弱与环境中的光强因子联系最为直接,能够表述植物的生长势。
不同遮荫处理下川芎苗期和第一次茎叶发生期的光合情况如表1所示,川芎苗期在不同遮荫下的净光合速率A差异显著,并随着遮荫程度的增加呈现逐渐下降趋势,相较CK,C1处理的净光合速率降低了22.17%,C2处理的净光合速率降低了50.1%,C3处理的净光合速率降低了82.17%;胞间CO2含量Ci亦在不同遮荫下呈现显著差异,并随着遮荫程度的增加呈现升高的趋势,在C3处理下含量最高,相较于CK,C1处理的胞间CO2含量升高了4.57%,C2处理的胞间CO2含量升高了5.63%,C3处理的胞间CO2含量升高了15.89%;苗期川芎的气孔导度gsw与蒸腾速率E随着遮荫程度的增加呈现一个缓慢下降的趋势,对照CK为最高值,与其他3个遮荫处理差异显著,3个遮荫处理之间差异不显著。

表1 不同遮荫处理对川芎光合作用的影响
第一次茎叶生长期不同遮荫下的净光合速率A差异显著,并随着遮荫程度的增加呈现先略微升高再下降的趋势,相较CK,C1处理的净光合速率升高了4.78%,为最高值,C2处理的净光合速率降低了30.72%,C3处理的净光合速率降低了63.99%,但是相较于苗期来说整体净光合速率值有所下降;胞间CO2含量Ci亦在不同遮荫下呈现显著差异,并随着遮荫程度的增加呈现逐渐升高的趋势,在C3处理下含量最高,相较于CK,C1处理的胞间CO2含量升高了14.82%,C2处理的胞间CO2含量升高了20.15%,C3处理的胞间CO2含量升高了28.09%;气孔导度gsw随着遮荫程度的增加呈现逐渐下降的趋势,CK处理为最大值,相较于CK,C1处理下降了37.66%,C2处理下降了47.4%,C3处理下降了50%;蒸腾速率E表现为CK处理下大于遮荫处理,与3个遮荫处理差异显著,3个遮荫处理之间差异不显著。
2.2 不同遮荫处理对川芎叶绿素荧光参数的影响
荧光参数是反映遮阴对植物叶片光合作用过程影响的重要指标,不同遮荫处理下川芎苗期和第一次茎叶生长期的荧光参数如表2所示,两个生长时期的初始荧光Fo和最大荧光Fm差异不显著,苗期Fo在177.67~190之间,Fm在994~1126之间,第一次茎叶生长期Fo在162~202.33之间,Fm在867.67~1142.67之间;而苗期的PSII最大光化学效率Fv/Fm则呈现随遮荫程度的增加先升高再下降的趋势,CK处理与C2处理差异不显著,与C1和C3差异显著,但变化幅度不大,相较于CK,C1处理下的PSII最大光化学效率升高1.46%,C3处理下的PSII最大光化学效率下降0.73%;第一次茎叶生长期的PSII最大光化学效率Fv/Fm在CK处理下为峰值,但各处理间变化亦不大,相较于CK,C1处理下的PSII最大光化学效率下降0.48%,C2处理下的PSII最大光化学效率下降0.24%,C3处理下的PSII最大光化学效率下降1.45%;两个时期的实际光化学效率Y(II)差异不显著,但整体表现为全光照下低于遮荫条件下,说明川芎为适应遮荫环境,提高了光子供给PSII反应中心的效率。
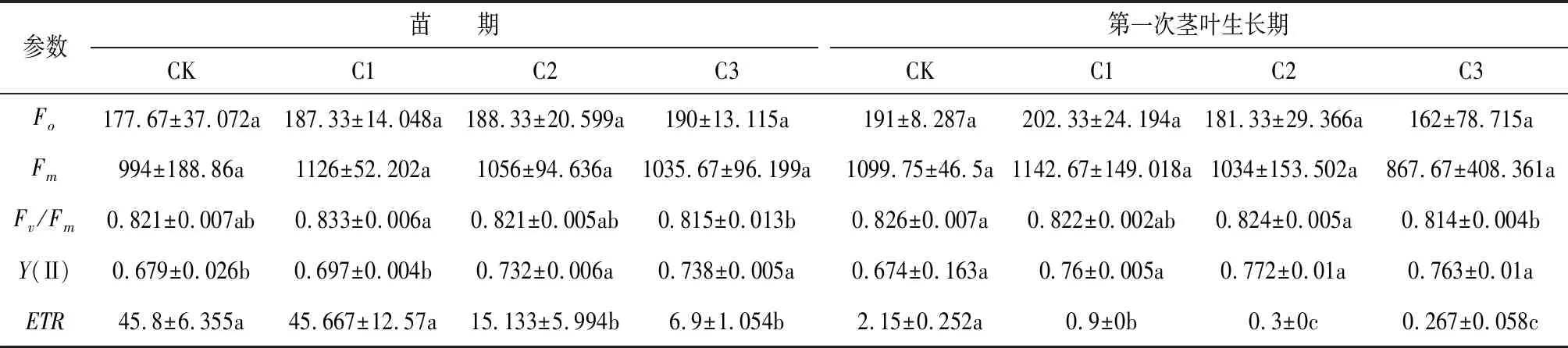
表2 不同遮荫处理下川芎的荧光参数
两个时期的电子传递速率ETR差异显著,均随遮荫程度的增加呈递减的趋势,CK处理下的苗期ETR与C1处理差异不显著,但是与C2和C3差异显著,C2处理的ETR下降66.96%,C3处理的ETR下降84.93%;第一次茎叶生长期的电子传递速率ETR亦表现出显著差异,相较CK,C1处理的ETR下降58.14%,C2处理的ETR下降86.05%,C3处理的ETR下降87.58%,可见全光处理下对激发态电子利用能力更强,电子的传递速率更快。
2.3 不同遮荫处理对川芎叶绿素含量的影响
光能直接影响叶绿素含量及比值的变化,从而影响光合作用。本研究中不同遮荫处理下川芎苗期和第一次茎叶生长期的叶绿素含量如表3所示,不同遮荫处理下苗期和第一次茎叶生长期的叶绿素总量和叶绿素b差异显著,并随着遮荫程度的增加呈现先增大再减小的趋势,在C2处理下为峰值,苗期的C1和C2处理下叶绿素总量和叶绿素b显著高于CK,叶绿素总量分别高出7.79%和9.88%,叶绿素b分别高出19.35%和27.64%;第一次茎叶生长期的C2和C3处理下叶绿素总量和叶绿素b显著高于CK,叶绿素总量分别高出25.43%和14.54%,叶绿素b分别高出76.6%和44.42%;两个生长时期的a/b值则随遮荫程度增加呈现先降低再回升的趋势,CK处理下为峰值,C2处理为谷值。整体看出遮荫条件下的叶绿素总量、叶绿素a以及叶绿素b均高于全光照,说明本试验中川芎叶片叶绿素含量受到了遮荫的显著影响,其为维持自身正常生长试图通过改变自身叶绿素含量及比例构成来增强其在弱光环境下的光捕获能力。
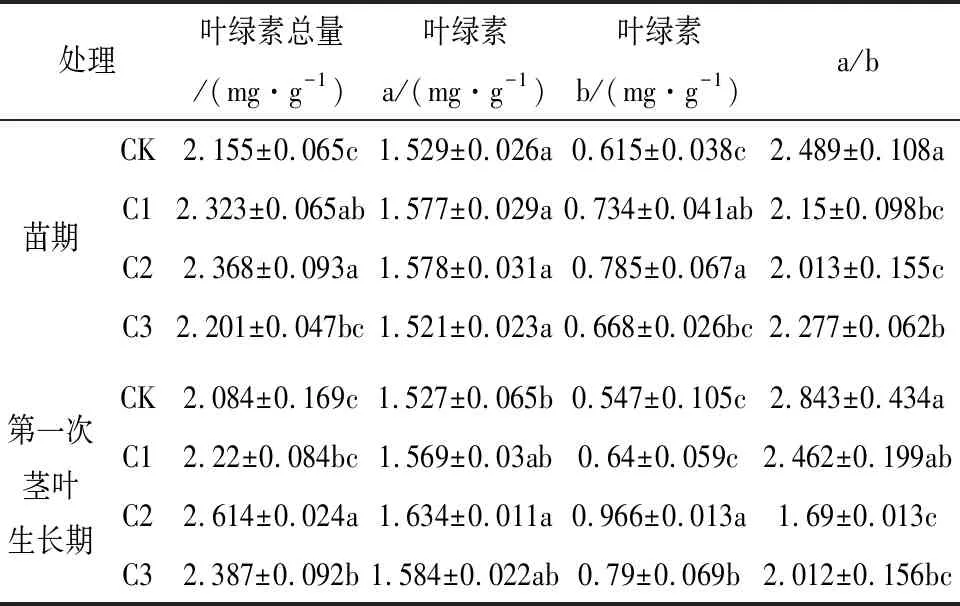
表3 不同遮荫处理下川芎的叶绿素含量
2.4 遮荫处理与川芎各项生长参数的相关性分析
遮荫处理与川芎生长各项参数相关性分析如表4和表5所示,遮荫处理也与苗期蒸腾速率、净光合速率和气孔导度极显著负相关,与胞间CO2含量显著正相关,与实际光化学效率Y(II)极显著正相关,与电子传递速率ETR极显著负相关;遮荫处理也与第一次茎叶生长期的净光合速率呈显著负相关,与胞间CO2含量极显著正相关,与电子传递速率ETR显著负相关,与总叶绿素和叶绿素b呈现显著正相关,和叶绿素a/b值呈现显著负相关。

表4 遮荫处理与川芎苗期光合荧光参数的相关性分析

表5 遮荫处理与川芎第一次茎叶生长期光合荧光参数的相关性分析
3 讨论
有研究表明,遮阴对光合作用的影响因植物种类、生育期和遮阴时间的不同结果有所差异,遮阴通常降低单叶的净光合速率[7],本研究结果显示,川芎苗期和第一次茎叶生长期的净光合速率A随遮荫程度的增加呈现显著降低的趋势,与前人的研究结论一致,且整体看来苗期的净光合速率A较第一次茎叶生长期的值高,说明受遮荫影响的时间越长,其光合能力越弱。Flexas J等对葡萄(VitisviniferaL.)的研究显示,其光合速率、蒸腾速率和气孔导度随着遮荫程度的提高而明显下降[8],与本研究结果的趋势一致。本研究中两个生长时期的净光合速率A与胞间CO2含量Ci的变化趋势并非一致,气孔导度gsw、蒸腾速率E与胞间CO2含量Ci的变化趋势亦不相同,说明胞间CO2含量和气孔导度不是影响其净光合速率下降的直接因素,而是遮荫造成的绝对低的光照环境,使其捕获的光能总量不足从而导致净光合速率的下降。
叶绿素荧光动力学技术在测定叶片光合作用过程中更具有反映“内在性”的特点[9],探讨叶绿素荧光参数的响应,可了解不同光照下植物光合作用的内部机理[3]。实际光化学效率Y(II)的大小反映了吸收的光子供应给PSII反应中心的效率[10],本研究中,Y(II)在两个时期中均表现为在全光照下的值低于遮荫条件下,说明川芎为适应遮荫环境,提高了光子供给PSII反应中心的效率。且电子传递速率ETR差异显著,均呈现随遮荫程度增加而逐渐降低的趋势,均在CK处理下为峰值,可见全光处理下对激发态电子利用能力更强,电子的传递速率更快。
叶绿素是植物的光合色素,具有吸收和传递光量子的功能,本试验中川芎叶绿素含量受到了遮荫的显著影响,遮荫条件下的叶绿素总量、叶绿素a以及叶绿素b均高于全光照,说明其为维持自身正常生长试图通过改变自身叶绿素含量及比例构成来增强其在弱光环境下的光捕获能力。本研究中叶绿素含量的变化趋势与熊淼[11]等对两种白及叶绿素含量的研究结论一致,均为对遮荫环境的一种内部适应。但是受光强的绝对限制,叶绿素的增加无法补偿光照不足造成的净光合作用下降,说明重度遮荫带来的影响已经超过了川芎自我保护调节的阈值,从而造成了不可逆的制约。
遮荫对植物叶绿素荧光和光合作用等的影响已有相当多的研究,所得出的结论主要是两个方面:一是遮荫可以促进植物的生长,增加植物生物量,提高光合作用效率;二是遮荫会延缓植物的生长,降低植物的光合作用,影响有机物的积累[11]。综上,遮荫对川芎的影响表现符合第二方面,川芎耐阴性弱,在弱光环境下生长情况较差。
遮荫环境不光直接改变了光强和光质,还会影响其温度和湿度等环境因子,因而,川芎在遮荫环境下的生理生态适应机制仍然是一个复杂的综合过程,为进一步摸清其对遮荫环境的响应机制,下一步可具体地量化不同环境因子,同时还要从叶片解剖结构、关键酶基因的表达等方面进行深入研究。
猜你喜欢
湖北农业科学(2021年13期)2021-08-04
中医眼耳鼻喉杂志(2021年2期)2021-07-21
新疆农业科技(2021年2期)2021-07-03
基层中医药(2021年11期)2021-06-05
中学生物学(2020年12期)2020-03-30
中成药(2018年7期)2018-08-04
特别健康(2018年2期)2018-06-29
新课程·下旬(2017年7期)2017-08-14
西北林学院学报(2015年5期)2015-01-03
生物技术通报(2014年10期)2014-03-21
