
航天诱变水稻雌性半不育突变体rs(s)的变异特性分析
2023-11-13 08:36赵华帅王永芬唐秀秀付澳秋卢家毅刘永柱
华南农业大学学报 2023年6期
赵华帅,朱 迪,王永芬,唐秀秀,付澳秋,卢家毅,王 慧,刘永柱
(华南农业大学 国家植物航天育种工程技术研究中心,广东 广州 510642)
航天诱变育种是指利用空间环境下的高真空、微重力、强辐射等因素使种子内部DNA发生重组,导致生物遗传性变异,进而在地面选育出新材料、新种质的作物育种新技术[1]。与地面传统诱变相比,航天诱变具有突变范围更广、频率更大、易出现特殊突变体等优势[2-3]。雌性不育现象在植物中普遍存在,相较于雄性不育,雌性不育更直接影响水稻的结实率[4]。研究者根据胚囊的发育特点,将其划分为8个阶段,分别是孢原细胞形成期、大孢子母细胞形成期、大孢子母细胞减数分裂期、功能大孢子形成期、单核胚囊形成期、胚囊有丝分裂期、八核胚囊发育期和胚囊成熟期[5]。雌性不育现象主要分为3种:雌性器官转化为雄蕊、雌蕊结构不完整和胚囊发育异常[6]。目前已获得的雌性不育突变体主要是由胚囊败育引起,这也是导致水稻雌性不育的主要原因。
通过物理、化学诱变,转基因技术等手段鉴别出的雌配子突变体,已成为水稻雌性不育的主要研究材料。突变体osrpala是通过T-DNA插入获得的1个雌性不育材料,该突变体胚囊完全败育,减数分裂异常,不能形成功能大孢子[7]。从EMS突变体库发现1个由单隐性基因控制的雌雄不育突变体mfs1,该突变体内外稃退化,胚囊内部结构分化不完全[8]。通过穗颈注射技术将大黍基因组DNA导入水稻中,获得1个雌性不育突变体fsv1,该突变体花粉育性正常,雌性高度不育,在雌配子发生过程中功能大孢子降解或功能退化,无法形成正常胚囊[9]。
有丝分裂是水稻有性生殖的关键过程,在这一时期功能大孢子经过3次有丝分裂形成完整的七细胞八核结构,为发育成成熟胚囊做准备[10]。目前,已克隆的参与水稻雌蕊有丝分裂过程的基因仅有OsDEES1和OsAPC6,OsDEES1通过信号传导调节胚囊早期发育,RNA干扰导致OsDEES1沉默,在有丝分裂期出现异常,功能大孢子逐渐退化,干扰雌配子体的形成[11-12]。OsAPC6编码一种与细胞周期相关的泛素连接酶,调节功能大孢子的有丝分裂,影响雌配子体的发育[13-14],但不影响雄配子发育。转录组测序分析发现,OsAPC6的转录水平在esd1幼穗分化的第6阶段显著降低,ESD1可能通过调控OsAPC6降解功能大孢子有丝分裂相关酶,促进卵细胞的程序性死亡[15]。
水稻雌性不育突变主要以插入突变和亚种间杂种为主,突变方式单一,没有其他性状突变体来源丰富,极大限制了学者们对水稻雌性不育的研究[16]。在前期研究中,华南农业大学国家植物航天育种工程技术研究中心利用航天搭载诱变,在后代中筛选到1个稳定的雌性半不育突变体;对该雌性不育突变体的研究将有利于揭示雌性不育变异特性,为水稻育性研究提供理论基础。
1 材料与方法
1.1 材料
本研究所使用的野生型材料原种‘秋B’(WT)是广西农业科学院选育的常规籼型水稻;雌性半不育突变体rs(s),是原种‘秋B’经航天诱变后选育的结实率降低,且在后代中能够稳定遗传的突变体。
1.2 小穗育性调查
2021—2022年在华南农业大学校内农场,按6×6小区种植野生型、突变体和正反交杂种F1,株、行间距20 cm;F1单株种植构建F2群体并双垄种植,所有材料的田间管理方法与一般生产大田相同。
在成熟期排除边际,野生型、突变体及F1植株各随机选取9株,每株选3个结实最好的穗子,调查结实率;利用野生型的花粉对rs(s)进行连续7 d的辅助饱和授粉,调查结实率;随机选取F2群体中206株植株,每株取3个结实最好的穗子调查结实率。
1.3 花粉育性和花粉离体萌发率考察
选取野生型与突变体rs(s)正在开花的主穗,在载玻片上滴入10 g/L I2-KI溶液,用镊子轻轻夹碎花药,释放花粉粒,在光学显微镜下观察花粉育性,判断标准:可育花粉呈圆形黑褐色,不可育花粉颜色较浅或无色[17]。花粉活力的测定采用花粉粒离体萌发的方法[18],取适量花粉放入滴有培养液的载玻片上,将载玻片放在垫有湿润滤纸的培养皿中,置于33 ℃培养箱中培养2 h,在显微镜下观察花粉离体萌发情况,花粉管长度超过花粉粒直径1/2的为萌发花粉。
1.4 整体染色与透明技术观察胚囊结构
取野生型与rs(s)即将开花的小穗,在FAA固定液中固定24 h,之后使用体积分数为50%、60%和70%的乙醇溶液逐级清洗;使用解剖针挑出子房;使用体积分数为50%、30%和10%的乙醇溶液和蒸馏水逐级对子房复水;在20 g/L的硫酸铝钾溶液中媒染;之后将子房放入10 mg/L的曙红染液中染色12 h;以体积分数为30%、50%、70%、85%和95%的乙醇溶液对子房进行梯度脱水,在无水乙醇与水杨酸甲酯体积比为1∶1的混合液体中预处理12 h,水杨酸甲酯单独处理12 h以上,在LSM880激光共聚焦显微镜下观察[19]。
1.5 内源激素含量测定
分别取野生型与突变体rs(s)在幼穗分化第5、6阶段的幼穗,作为激素测定样品,每个时期3次生物学重复,测定幼穗中生长素(IAA)、细胞分裂素(TZR)、脱落酸(ABA)和赤霉素(GA3)的含量,并计算平均值。
1.6 数据分析
本研究所有数据使用SPSS软件和Excel软件进行处理;用卡方检验来验证雌性半不育突变体的遗传特性。
2 结果与分析
2.1 育性分布
由图1可知,野生型结实率80.9%,突变体rs(s)结实率54.6%,相比野生型下降了26.3个百分点,差异极显著(P< 0.01);以原种‘秋B’为父本对突变体rs(s)进行连续7 d的辅助饱和授粉,结实率为58.3%,与rs(s)自然授粉结实率差异不显著;野生型和rs(s)正反交杂种F1的结实率,分别是77.36%和75.28%,差异不显著;野生型与F1的结实率差异不显著,但突变体rs(s)与野生型和F1差异极显著;推测半不育表型受隐性基因调控,导致rs(s)半不育的原因与雌蕊发育异常有关。

图1 野生型(WT)与突变体rs(s)及其正反交F1和辅助授粉的结实率Fig.1 The setting rates of wild type (WT),mutant rs(s) as well as their reciprocal cross F1 and auxiliary pollination
以野生型为母本、突变体rs(s)为父本构建F2群体,调查群体中206株植株的结实率。如图2所示,小穗育性表现出较大的变异幅度,从15%~94%呈现连续的双峰分布,在55%和80%处有较明显的2个峰,最低谷出现在70%左右,以70%为界线进行分组,可以分成结实率高于70%的可育株(151株)和结实率低于70%的半不育株(55株) 2个集团,对可育株集团和半不育株集团进行卡方测验,χ2=0.233<χ0.052=3.38,可育植株与半不育植株的数量比例符合1对基因的分离比3∶1,说明突变体rs(s)的半不育表型受1对隐性基因控制。

图2 F2群体结实率分布直方图Fig.2 Distribution histogram of setting rate of F2 population
2.2 亲本的花粉育性和胚囊育性
对亲本的表型性状进行调查发现,与野生型相比,突变体rs(s)的株高、穗长均有所增加(图3a、3b);突变体rs(s)的花器官外部结构及特征与野生型差异明显,雄蕊结构完整且花药饱满,整体发育正常,但雌蕊表现出子房干瘪瘦小、柱头伸长不明显等特征(图3c、3d)。细胞学观察显示,野生型和突变体rs(s)的花粉育性分别为93.02%和90.75%,均达到90%以上(图4);花粉粒离体萌发率分别为78.88%和80.33%,均达到80%左右(图5)。经t检验分析,野生型和rs(s)的花粉育性和花粉离体萌发率均未表现出显著差异,表明突变体rs(s)的雄蕊发育正常。

图3 野生型(WT)与突变体rs(s)表型及花器官结构观察Fig.3 Phenotype and floral organ observation of wild type (WT) and mutant rs(s)

图4 野生型(WT)与突变体rs(s)花粉碘染观察Fig.4 Observation of iodine staining for wild type (WT)and mutant rs(s) pollens

图5 野生型(WT)与突变体rs(s)花粉粒离体萌发观察Fig.5 Observation of in vitro germination for wild type(WT) and mutant rs(s) pollen grains
对野生型和突变体rs(s)的成熟子房进行观察,128个野生型胚囊中,正常率为89.06%,139个突变体胚囊中,正常率为59.71%,突变体胚囊的正常率相比野生型下降了29.35个百分点。胚囊结构见图6。图6a为正常可育的胚囊,具有完整的七细胞八核结构,胚囊内部包含1个卵细胞、2个助细胞、2个极核和3个反足细胞;胚囊退化占异常胚囊总数的14.30%,表现为空胚囊,胚囊内无任何细胞结构(图6b);胚囊中无卵器占异常胚囊总数的67.90%,主要表现为胚囊内只有反足细胞团(图6c);极核异常主要表现为极核游离(图6d),极核数目增加或减少及双腔等类型相对较少(图6e、6f)。76.80%的异常胚囊存在不同程度的七细胞八核结构不完整现象,但存在细胞结构,推测大部分胚囊在减数分裂阶段正常,能够形成功能大孢子,但在有丝分裂阶段出现异常,导致胚囊败育。

图6 突变体胚囊败育的主要类型Fig.6 Main types of embryo sac abortion in mutant
2.3 野生型和突变体rs(s)的激素含量变化
为探究水稻雌性半不育突变体的分子影响机制,我们首先测定了野生型与突变体rs(s)在幼穗分化第5、6阶段的幼穗激素含量变化。由图7可知,突变体rs(s)的IAA含量在第5阶段与野生型差异不大,但在第6阶段极显著增加,比rs(s)第5阶段增加了76.2%,比野生型第6阶段增加了61.5%,经Duncan’s多重比较分析,均在P< 0.01水平差异显著,而野生型在第5阶段和第6阶段差异不明显。rs(s)中TZR含量在第5、6阶段均低于野生型,在第6阶段差异极显著(P< 0.01),且在第5、6阶段呈现下降趋势,第6阶段相比第5阶段下降了17.5%,相比野生型第6阶段下降22.7%。野生型与rs(s)在幼穗分化第5、6阶段的ABA与GA3含量未表现明显的差异。因此,推测在幼穗分化第6阶段生长素含量的增加和细胞分裂素的下降,可能影响了突变体rs(s)雌蕊的生长发育。
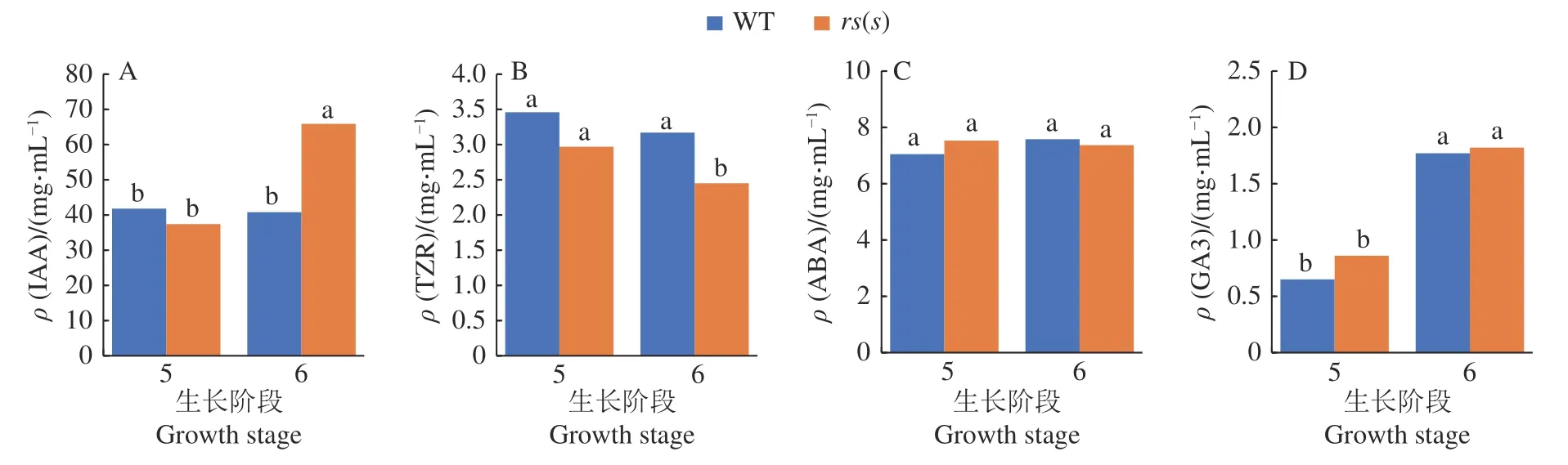
图7 野生型(WT)和突变体rs(s)的激素含量变化Fig.7 Changes of hormone content in wild type (WT) and mutant rs(s)
3 结论与讨论
3.1 rs(s)可能是一个新的雌性半不育突变体
水稻雌配子败育现象较少,通常伴随雄性不育,单纯雌不育突变体不易获得,典型的雌性不育植株通常表现出种子发芽率低、子房干瘪、无花柱和柱头或柱头伸长不明显等特征[20]。减数分裂和有丝分裂是水稻有性生殖的2个关键过程,包含细胞数目的增加以及功能分化,决定胚囊的正常发育[10]。水稻雌性不育的形成原因复杂,在雌蕊发育的过程中,任何一个时期发生异常都可能会导致雌性不育[21]。目前对雌性不育的研究主要集中在减数分裂阶段的同源染色体配对和功能大孢子,而对有丝分裂异常导致的雌性不育现象的研究较少[22]。本研究通过航天诱变获得的雌性半不育突变体rs(s),雄蕊发育正常,但雌蕊在胚囊发育的有丝分裂阶段异常,不能形成完整的七细胞八核结构;花器官表现出子房干瘪瘦小、花柱和柱头伸长不明显等典型雌性不育特征,与前人的结论[20]一致,且胚囊发育在有丝分裂阶段发生异常,败育时期特殊,材料来源新颖,可能是一个新的雌性半不育突变体,具有较高的研究价值。
3.2 生长素参与调控水稻雌蕊发育
通过对水稻、石榴和白菜等雌性不育材料进行转录组分析发现,差异表达基因在激素的信号转导通路显著富集[23-26]。生长素的内稳态和极性运输是各种雌蕊组织形成的关键因素,在白菜fsm突变体中,随着生长素含量升高,雌蕊的生长发育受到抑制,导致子房干瘪瘦小、柱头伸长不明显,胚囊败育[27-28]。研究表明植物激素转导相关基因的缺失会抑制一核、二核和四核阶段的有丝分裂,包括一些生长素运输(PIN1、AUX1、LAX1),生长素合成(YUC1、YUC2、YUC8、TAA1)和细胞分裂素合成(CKX、LOG)相关基因。AUX1/LAX是生长素转运蛋白家族主要的输入载体,PIN是主要的输出载体,PIN基因表达量下调,使生长素外向转运受到抑制,雌蕊中生长素含量升高,抑制雌蕊生长[29]。细胞分裂素主要存在于根尖、嫩叶、幼穗等分裂旺盛的部位,可以调控细胞周期的G1/S期和G2/M期的转变及心皮的发育,在拟南芥的心皮中检测到强烈的细胞分裂素响应信号[30]。研究表明,生长素含量升高会抑制蔗糖转运蛋白的活性,导致蔗糖从源到库的流通不畅,叶片中的蔗糖无法为雌蕊的生长发育提供足够的营养物质[31-32]。突变体rs(s)在幼穗分化的第6阶段,生长素含量急剧增加,导致雌蕊发育畸形,子房干瘪瘦小,胚囊发育停滞在有丝分裂阶段,但胚囊发育异常是否与生长素抑制了蔗糖转运蛋白的活性有关,仍需进一步研究。
猜你喜欢
华北农学报(2022年4期)2022-09-14
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
东北农业大学学报(2020年12期)2021-01-15
江苏农业科学(2019年10期)2019-07-08
江苏农业科学(2016年11期)2017-03-21
红蜻蜓·低年级(2016年9期)2016-05-14
中国民族民间医药·下半月(2015年4期)2015-05-22
