
山桃叶片光合生理参数对土壤水分的响应特征
2023-11-09 10:36刘传林赵红霞孙效静彭广伟赵万里夏江宝
山东林业科技 2023年5期
郑 岩,刘传林,赵红霞,孙效静,彭广伟,张 军,赵万里,夏江宝*
(1.山东高速鸿林工程技术有限公司,山东 济南250001; 2.济南南郊宾馆园林部,山东 济南250002;3.山东省国土空间数据和遥感技术研究院,山东 济南250002; 4.滨州学院,山东省黄河三角洲生态环境重点实验室,山东 滨州256603)
陆地生态系统面临着降水的高度时空异质性,加之全球变暖的影响,极端干旱或洪水事件的强度、范围和频率不断增加[1]。降水在不同的尺度上影响着各种植物生理和生态过程[2]。大多数植物物种的分布和丰度在很大程度上依赖于供水条件,在干旱期间,缺水会通过影响植物光合作用和养分吸收等一系列生化和生理过程,从而对植物生长产生不利影响[3]。为了逃避或减少水分胁迫的影响,一些植物通过改变生理生态性状来适应不断变化的环境,如降低冠层气孔导度响应蒸腾驱动因子的敏感性,降低叶片光合能力或叶面积和相对生长率下降等[2,4]。光合作用是植物代谢发育的敏感生理参数,能快速反映植物对环境因子变化的响应[5],是决定植物生产力和作物产量的决定性因素[6]。在这一过程中,叶片净光合速率与光合有效辐射之间的响应关系是揭示植物光合生理过程对环境响应的基础[7-9]。光合光响应曲线的测量和模拟是了解植物光合生理生态过程的重要工具,是揭示光合生理过程对环境响应的基础[9]。通过光合光响应曲线可计算植物光合作用的最大净光合速率、光饱和点、光补偿点、暗呼吸速率以及表观量子效率等光合生理参数[10-12],从而探讨植物光合机构的运行状态、光合能力和光合效率及其影响因素[10,13]。直角双曲线模型[14]、非直角双曲线模型[15]、直角双曲线修正模型[16]和指数模型[17]是目前常用的光合光响应模型。由于不同模型对同一植物光合光响应参数拟合值存在差异[18],因此通过筛选适宜的光合光响应模型,优化光合效率参数,探讨植物光合生理参数随土壤水分的变化规律,可以更深入的了解植物的光合生理特征,可为植物的合理栽植提供理论依据。
山桃(Prunus davidiana),蔷薇科(Rosaceae)李属(Prunus),乔木。常生于山坡、山谷沟底或荒野疏林及灌丛内。山桃抗旱耐寒且耐盐碱,根系发达,分支能力强,兼具观赏、药用及生态防护价值,是优良的矿区水土保持植被恢复和荒山造林绿化树种。目前关于山桃的研究主要集中在栽培技术管理[19],生态防护功能[20-21],盐碱、干旱胁迫下及不同林龄的山桃光合生理特征[20-23]、生长特征[24]、生长稳定性[25]和适宜区分布[26]等方面。已有研究表明,水分胁迫下,山桃既可以通过增加根冠比等生长特征应对干旱条件[24],也可通过较大幅度降低蒸腾速率来提高其水分利用效率从而增强其适应干旱环境的能力[22]。但对不同水分条件下山桃幼苗适宜的光合光响应模型及其光合参数的优化筛选探讨较少,基于山桃幼苗叶片光合生理参数的适宜水分条件尚不清晰,在一定程度上影响了山桃幼苗栽植的水分管理。鉴于此,为明确矿区废弃地山桃幼苗生长适宜的水分条件,本研究以3 a 生山桃幼苗为研究对象,模拟设置石灰岩矿区潮壤土的5 个水分梯度,测定不同土壤水分条件下山桃叶片的光合光响应过程,探讨不同土壤水分条件下山桃叶片的光合特性;利用非直角双曲线模型、直角双曲线模型和直角双曲线修正模型对不同土壤水分条件下山桃叶片光合光响应过程进行拟合分析,筛选出山桃叶片适宜的光合光响应模型,探讨其光合作用光响应过程及其主要特征参数对土壤水分的响应规律,以期为石灰岩山地矿区废弃地的植被恢复与生态修复提供理论依据和技术参考。
1 材料与方法
1.1 试验材料
本研究以3 a 生山桃实生苗为试验材料,幼苗根径平均为1.8 cm,株高统一截干为0.50 m。试验栽植盆钵为直径30 cm,高40 cm,每盆栽植1 株,共栽植10 株。模拟控制实验在山东省黄河三角洲生态环境重点实验室的智能科研温室进行,空气相对湿度平均为36%,采用自然光照,玻璃透光率超过85%,温度平均为28℃。试验用栽植土壤取自山东省泰安市东平县金山矿区,土壤类型为潮壤土,土壤容重平均为1.35 g·cm-3,田间持水量为20.51%,土壤颗粒组成为2.0~1.0 mm 的石砾含量为20%,1.0~0.25 mm 的粗砂粒含量为55%,0.25~0.05 mm 的细砂粒含量为22%,<0.05 mm 的粉粘粒含量为3%。
1.2 试验设计及指标测定
以3 a 生山桃实生苗木为试验材料,2022 年3 月份进行苗木盆栽,正常生长2 月后,通过饱和灌水后自然蒸发和植物蒸腾获取不同的土壤水分梯度,选取生长良好的山桃苗木6 株(6 盆),1 d 后获得初期水分梯度,烘干法测定0~30 cm 土壤质量含水量(SWC;%),并测算相对含水量(RWC;%),SWC 为18.56%(RWC 为90.49%),以后每隔2~3 d 获取一个水分梯度。在SWC 为15.23%、12.24%、9.56%和6.21%(RWC 分别为74.26%、59.68%、46.61%和30.28%)时分别进行测定。
从土壤水分处理的6 株苗木中,随机选取3 株苗木采用LI-6400(美国LI-COR 公司)便携式光合仪进行光合光响应过程的测定,每株苗木测定2~3 片成熟叶片,每个叶片重复计数3 次。测定时间为9:00~11:00左右,使用大气CO2浓度并外接缓冲瓶保证气路稳定,将光合有效辐射(PAR)控制在2000、1800、1600、1400、1200、1000、800、600、400、250、150、100、50、20 μmol·m-2·s-1。每个PAR 下测定时长为2 min。仪器自动记录净光合速率(Pn;μmol·m-2·s-1)、光合有效辐射(PAR;μmol·m-2·s-1)、蒸腾速率(Tr;mmol·m-2·s-1) 等参数,测算水分利用效率(WUE;μmol·mmol)为净光合速率/蒸腾速率。
1.3 数据处理
1.3.1 非直角双曲线模型
非直角双曲线模型表达式[15,27]
式中,α 为初始量子效率;I 为光合有效辐射;Pmax为光饱和点处最大光合效率;θ 为非直角双曲线模型的曲角(0<θ≤1);Rd为暗呼吸速率。
1.3.2 直角双曲线模型
直角双曲线模型表达式[14,28-29]
式中,α 为初始量子效率;I 为光合有效辐射;Pmax为光饱和点处最大光合效率;Rd为暗呼吸速率。
1.3.3 直角双曲线修正模型
直角双曲线修正模型表达式[30]
式中,α 为初始量效率;β 为修正系数;γ 为初始量子效率与最大光合速率之比;Rd为暗呼吸速率。
2 结果与分析
2.1 山桃叶片净光合速率对土壤水分的响应
由图1 可知,土壤水分对山桃叶片净光合速率影响显著(P<0.05),在相同光合有效辐射下,随土壤水分的降低,山桃叶片Pn 呈现先升高后降低的趋势,在RWC 为74.26%条件下达最高值,整体表现为RWC74.26%>RWC90.49% >RWC59.68% >RWC46.61% >RWC30.28% 。RWC90.49% 、RWC59.68% 、RWC46.61%、RWC30.28%条件下的平均Pn分别比RWC74.26% 降低17.43% 、29.22%% 、45.45%、63.22%。在不同土壤水分条件下,当PAR≤600 μmol·m-2·s-1时,叶片Pn 随PAR升高呈现快速上升趋势,对PAR 响应敏感,随后缓慢增大至光饱和点,对应出现最大Pn,但光合光响应过程因土壤水分条件差异而不同。当PAR>600 μmol·m-2·s-1时,在RWC90.49%、RWC74.26%、RWC59.68%条件下,叶片Pn仍有不同程度的明显增加,分别在PAR 为1600 μmol·m-2·s-1、1800 μmol·m-2·s-1、1600 μmol·m-2·s-1时达到光饱和点;在RWC46.61%、RWC30.28%条件下趋于稳定,分别在PAR为1200 μmol·m-2·s-1、800 μmol·m-2·s-1时达到光饱和点。
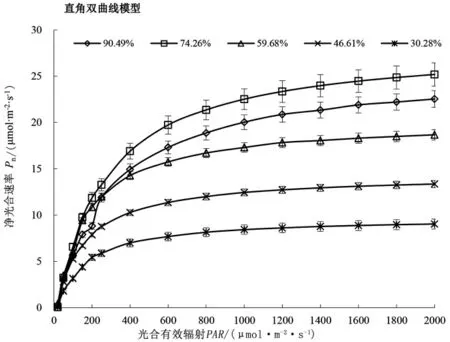
图1 不同土壤水分下山桃叶片净光合速率的光响应Figure 1 Light response of net photosynthetic rate of P.davidiana leaves under different soil moisture
2.2 不同土壤水分条件下山桃叶片净光合速率的光响应模拟
利用非直角双曲线模型、直角双曲线模型、直角双曲线修正模型拟合出了不同土壤水分条件下山桃叶片净光合速率光响应曲线(图2),均能较好的模拟出在一定光强范围内Pn随PAR 的增加而增加,且各模拟曲线的趋势与实测值所绘出的曲线趋势相同。由图2 和表1 可知,不同土壤水分条件下直角双曲线修正模型拟合的最大净光合速率(Pnmax)和光饱和点(LSP)均接近实测值;但直角双曲线模型和非直角双曲线模型拟合出的Pnmax均远大于实测值,拟合的LSP 又远小于实测值;表观量子效率(α),光补偿点(LCP)及暗呼吸速率(Rd)等参数的模拟值与实测值随水分条件的不同表现出各自的适宜性。由表1 可知,三个模型都能较好地拟合山桃叶片净光合速率光响应过程,决定系数R2均在0.975 以上,其中以直角双曲线修正模型的拟合效果最好,不同土壤水分条件下R2均大于0.99。从决定系数R2来看3 个模型拟合效果为直角双曲线修正模型>非直角双曲线模型>直角双曲线模型。
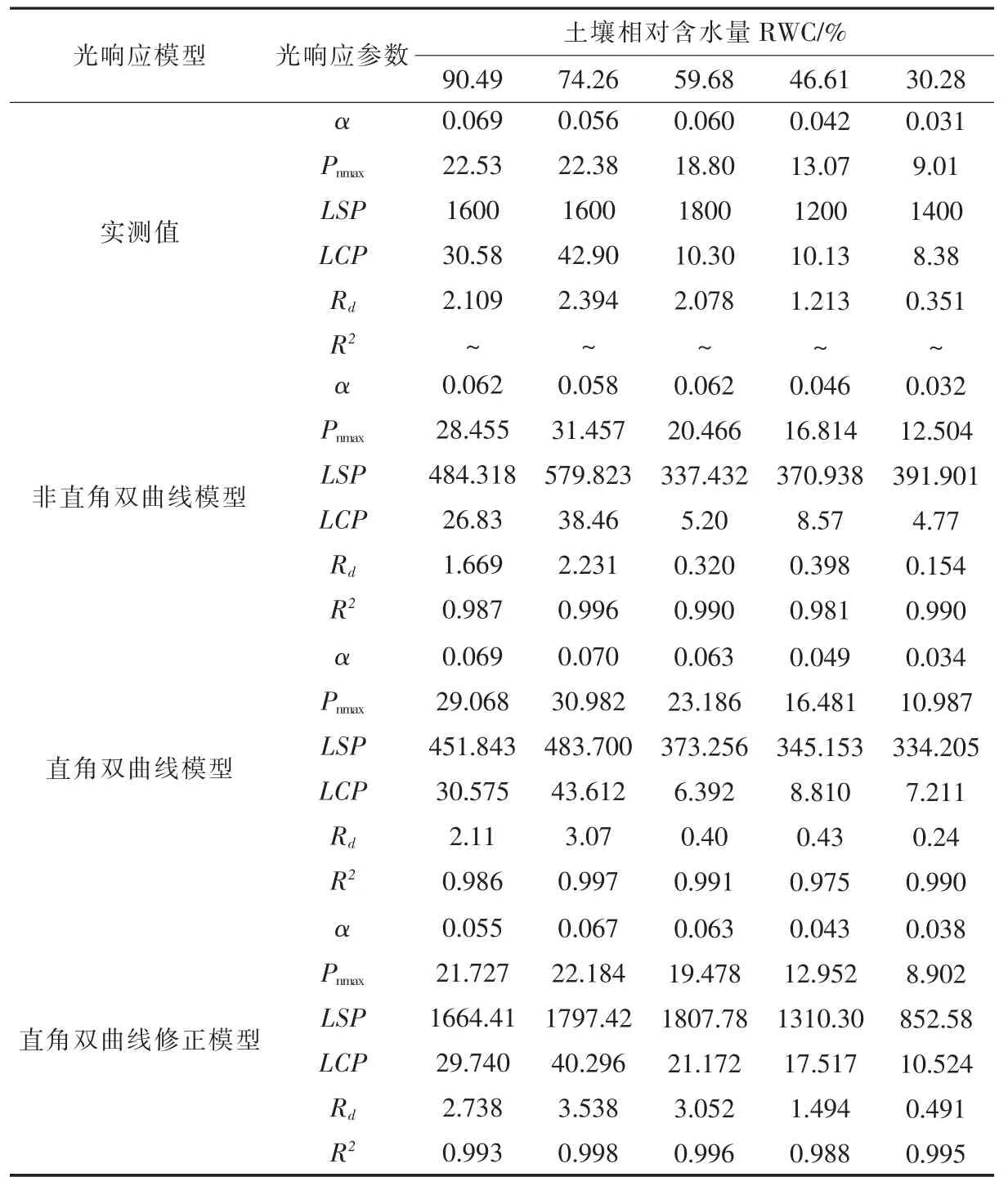
表1 不同土壤水分下山桃叶片光合光响应模型参数Table 1 Photosynthetic light response model parameters of P.davidiana leaves under different soil moisture
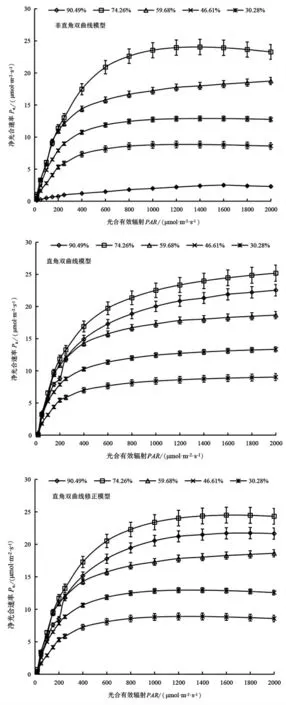
图2 不同土壤水分下山桃叶片光合光响应模型曲线Figure 2 Photosynthetic light response model curve of P.davidiana leaves under different soil moisture
2.3 山桃叶片光合光响应参数对土壤水分的响应
由图2 和表1 可知,直角双曲线修正模型能较好的拟合山桃叶片光合作用光响应过程,以直角双曲线修正模型求解的α、Pnmax、LSP、LCP 和Rd进行分析。由表1 可知,土壤水分对山桃叶片α、Pnmax、LSP、LCP 及Rd影响显著(P<0.05)。随土壤水分的降低,山桃叶片α、Pnmax、LSP、LCP 及Rd均呈现先升高后降低的趋势。α、Pnmax、LCP 及Rd均在RWC74.26%条件下达到最大值,RWC90.49%、RWC59.68%、RWC46.61%、RWC30.28%条件下α 值分别比RWC74.26%降低17.91%、5.97%、35.82%、43.28%;Pnmax分别降低2.06%、12.20%、41.62%、59.87%,LCP 分别降低26.20% 、47.46% 、56.53% 、73.88%,Rd分别降低22.61%、13.74%、57.77%、86.12%。LSP 在RWC59.68%达最高值,与其相比RWC46.61%、RWC30.28%条件下分别下降27.52%、52.84%。
2.4 山桃叶片蒸腾速率和水分利用效率对土壤水分的响应
由图3 可知,土壤水分对山桃叶片蒸腾速率(Tr)影响显著(P<0.05)。随土壤水分的降低,山桃叶片Tr 及其光饱和点呈现先升高后降低的变化趋势,在RWC59.68 %条件下Tr 达最高值,与其相比RWC90.49%、RWC74.26%、RWC46.61%和RWC30.28%平 均Tr 分别下降30.45% 、14.21% 、40.89%和54.79%;山桃叶片Tr 的光饱和点 在1200 -1800 μmol ·m-2·s-1,在RWC30.28%下Tr 光饱和点达到最低值。当PAR<1400 μmol·m-2·s-1时,随PAR 增加,不同土壤水分条件下Tr 均呈现上升趋势。当PAR>1400 μmol·m-2·s-1时,随PAR 增加,在RWC30.28%、RWC46.61%条件下,Tr 呈现下降趋势;在RWC90.49%、RWC74.26%、RWC59.68% 条件下,Tr 仍有不同程度的增加,分别在PAR为1600 μmol·m-2·s-1、1800 μmol·m-2·s-1、1800 μmol·m-2·s-1时Tr 达到最大值。
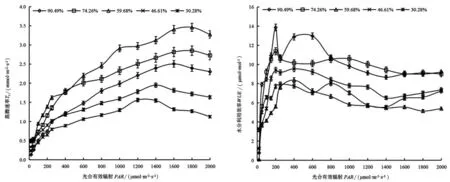
图3 不同土壤水分下山桃叶片蒸腾速率和水分利用效率的光响应Figure 3 Light response of transpiration rate and water use efficiency of P.davidiana leaves under different soil moisture
由图3 可知,土壤水分对山桃叶片水分利用效率(WUE)影响显著。随土壤水分的降低,山桃叶片WUE 整体呈现先下降后升高趋势,在RWC 59.68%处理下达最低值。与RWC59.68% 平均WUE (5.42 μmol·mmol )相比,高水分RWC90.49%、RWC74.26%平均WUE 分别升高77.23%和70.37%,低水分RWC46.61% 、RWC30.28%平均WUE 分别升高35.10%、18.57%。不同水分条件下,山桃叶片WUE随PAR 的升高表现为先迅速升高后缓慢下降的趋势,当PAR<250 μmol·m-2·s-1时,随PAR 增加,WUE 呈现迅速升高趋势;在PAR 为250~400 μmol·m-2·s-1达到WUE 的光饱和点,在PAR >4000 μmol·m-2·s-1时WUE 呈现缓慢下降趋势。
3 讨 论
植物光合光响应模型在植物光合生理参数的精准测算方面起到了重要作用,也是求解植物叶片表观量子效率、光补偿点、光饱和点、最大净光合速率等指标的主要方法[10,29]。非直角双曲线模型、直角双曲线模型和直角双曲线修正模型均能较好模拟不同土壤水分条件下山桃叶片净光合速率的光响应曲线,从接近实测值和R2来看,直角双曲线修正模型可较好模拟山桃叶片光合光响应曲线,但不同模型在具体光合光响应参数的测算上呈现各自的适宜性[8,18]。植物叶片的光补偿点、光饱和点、最大净光合速率、表观量子效率和暗呼吸速率是逆境条件下反映植物光合生理过程的主要指标[31]。受全球气候变化的影响,在未来出现极端降水或干旱事件的概率和影响程度有所增加[32],降水通过影响土壤水分的变化进而作用于植物的生长、发育和演替[33]。随土壤水分的降低,山桃叶片净光合速率及最大净光合速率均呈现先升高后降低的趋势,在土壤相对含水量为74.26%时达到最大值。荒漠地区白刺(Nitraria tangutorum)等3 种荒漠灌木灌水后净光合速率日均值低于灌水前[34],鲁中地区野生酸枣(Ziziphus jujuba var.spinosa)、青海云杉(Picea crassifolia),呼和浩特大青山南麓油松(Pinus tabuliformis)的光合作用也表现出随土壤水分的降低先升高后降低的趋势[35-37],药用植物马比木(Nothapodytes pittosporoides(Oliv.) Sleum.)表现出先下降后逐步升高再迅速降低的趋势[38],相关研究表明植物叶片净光合速率对土壤水分存在一定的阈值效应。在正常水分供应下,山桃叶片最大可能截获光能,促进自身的生长发育,但由于土壤水分的降低则使其光合电子传递能力和能量分配减弱,破坏植株体内水分平衡,并引起气孔关闭和光合速率下降[39],导致光合作用能力下降进而降低植物的活力,影响植物生长[40]。
光补偿点、光饱和点表征了植物对光照的需求,由此可判断植物对强光和弱光条件的适应能力。阴生植物的光饱和点、光补偿点通常分别在500 μmol·m-2·s-1、20 μmol·m-2·s-1之下;阳生植物的光饱和点、光补偿点通常分别在800 μmol·m-2·s-1、50 μmol·m-2·s-1以上[41]。在不同土壤水分条件下,山桃叶片光饱和点处于852.58-1807.78 μmol·m-2·s-1之间,光补偿点处于10.524—40.296 μmol·m-2·s-1之间,由此可见,山桃叶片的光照生态幅较宽,对强光及弱光的适应性均较强,阳生植物特性显著。但随着土壤水分的降低,山桃叶片的光照生态幅先升高后降低,在土壤相对含水量为59.68%时光照生态幅最宽,在土壤相对含水量为46.61%、30.28%时光照生态幅变低分别为1292.783 和842.056 μmol·m-2·s-1。表观量子效率是反映植物对弱光利用能力的指标[42]。在本研究中,随土壤水分的降低,山桃叶片表观量子效率呈先上升后下降趋势,说明适当的干旱胁迫会提升山桃叶片对弱光的利用能力,但重度水分亏缺会降低山桃对光能的利用。但也有研究发现马比木叶片表观量子效率随土壤水分的减少呈逐渐减低趋势[38]。综上所述,山桃叶片光合作用对土壤水分存在一定的阈值效应,水分过高或过低山桃叶片光合作用均呈现下降趋势,但高水分条件下山桃叶片的光合作用显著高于低水分条件,适宜的水分条件可显著提升山桃叶片的光合能力;干旱胁迫降低了山桃叶片对弱光以及光强的利范围,山桃叶片对光环境的适应能力减弱。
水分利用效率是反映干旱胁迫条件下植物生长和光合生理性状有关的功能指标[42]。水分利用效率低表明植物缺乏良好的保水能力,无法适应过度干旱环境[43]。干旱胁迫下植物持续需要碳水化合物维持新陈代谢会耗尽碳水化合物储备,导致碳饥饿,并在长时间条件下导致植物死亡[44]。在干旱和营养不良地区维持较高水力调节和光合作用的植物表现出较高的水分利用效率,从而获得更大的生存和性能[45-46]。在本研究中,随土壤水分的降低,山桃叶片的净光合速率和蒸腾速率均表现为先升高后降低的趋势,但两者在不同水分条件下达到最高值,分别是土壤相对含水量为74.26%和59.68%,而水分利用效率表现为先下降后升高趋势,在土壤相对含水量为59.68%时达最高值。表明山桃叶片蒸腾速率比净光合速率对土壤水分降低的敏感程度更高、下降程度更大,通过降低蒸腾速率减少水分散失,增加气孔阻力,降低气孔导度[47],以达到提高水分利用效率从而增强其适应干旱环境的能力,这与黄土丘陵区干旱胁迫下山桃以降低蒸腾速率提高瞬时水分利用效率的结论类似[22]。但土壤水分与植物叶片水分利用效率之间的响应关系研究存在较大差异,例如随着土壤水分的降低,鲁南地区野生酸枣叶片净光合速率、蒸腾速率、水分利用效率均先升高后降低[36],贵州马比木叶片蒸腾速率整体下降,水分利用效率先下降后升高再下降趋势[38],白刺等3 种荒漠灌木灌水后水分利用效率日均值升高,蒸腾作用对土壤水分的变化响应比光合作用敏感[34],水分胁迫对澳大利亚小麦品种水分利用效率影响不大[48],表明植物叶片净光合速率、蒸腾速率对土壤水分响应的敏感程度不一样,同时受植物种类、研究区域或模拟水分条件的不同,导致瞬时水分利用效率与土壤水分之间存在不同的响应关系。
4 结论
非直角双曲线模型、直角双曲线模型、直角双曲线修正模型均可拟合不同土壤水分条件下山桃叶片光合光响应曲线,但从接近实测值和R2来看,直角双曲线修正模型最优。随干旱胁迫的加重,山桃叶片表观量子效率、最大净光合速率、光饱和点、光补偿点和暗呼吸速率均呈现先升高后降低的趋势,除光饱和点在土壤相对含水量59.68%达最高值外,其他光合光响应参数在土壤相对含水量为74.26%达最高水平。随土壤水分的降低,山桃叶片净光合速率和蒸腾速率呈现先升高后降低的变化趋势,分别在土壤相对含水量74.26%和59.68%条件下达最高值,而水分利用效率表现为先降低后升高的趋势,在土壤相对含水量90.49%条件下达最高值。高水分条件下山桃叶片光合作用、蒸腾作用和水分利用效率显著高于低水分条件;随土壤水分的降低,山桃叶片呈现蒸腾作用比光合作用更加敏感的响应表现,呈现出干旱胁迫下以降低蒸腾作用提升水分高效利用的生理特性。
猜你喜欢
动漫界·幼教365(小班)(2021年6期)2021-07-21
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
新课程·下旬(2017年7期)2017-08-14
北方文学·中旬(2017年1期)2017-03-15
小说月刊(2016年1期)2015-12-29
西北林学院学报(2015年5期)2015-01-03
新课程·中旬(2014年10期)2014-12-26
中国蜂业(2013年11期)2013-01-23
植物营养与肥料学报(2011年2期)2011-10-26
