
蝎毒多肽优化体CT-K3K7抗食管癌的机制研究*
2023-11-09 01:12:26张家铭水颍彬袁亚萍高社干李钟杰
华中科技大学学报(医学版) 2023年5期
张家铭, 水颍彬, 杨 青 , 袁亚萍, 高社干, 李钟杰△
1河南科技大学基础医学与法医学院,洛阳 471000 2河南科技大学第一附属医院,洛阳 471000
食管癌是全球第八大常见类型癌症,同时也是我国所有癌症中第五大最常见的类型[1-3]。作为我国第四大致死性肿瘤,我国的食管癌年发病数量大约25万例,5年生存率低于20%,每年约19万例患者死于食管癌[4-6]。手术、化疗和放疗是现代医学中治疗食管癌的常规手段,但肿瘤耐药性是食管癌防治中最为棘手的问题。高耐药性指的是肿瘤细胞对抗癌药物的敏感性降低,从而使得疗效变差,患者生存期缩短。因此,迫切需要开发新的治疗方案和研发新型治疗药物。在发掘新的有效药物方面,多肽类活性分子是研究热点之一,其具有毒副作用小、耐受性强等优点[7-8]。
蝎子在世界范围内用作传统药物已有数千年的历史,蝎毒液中的多肽是其主要的药用活性成分[9-10]。已有研究表明,从不同蝎种中获得的毒液多肽在神经、免疫、感染、心血管、肿瘤等多系统疾病中都有着良好的治疗效果[11]。蝎毒多肽Ctry2459[12]是从西藏特里豚蝎Chaerilustryznai中发现的一种抗微生物多肽,而CT-K3K7是基于Ctry2459的优化体多肽,其具有良好的抗菌活性,并被认为是治疗金黄色葡萄球菌和铜绿假单胞菌感染的潜在抗菌剂候选物[13]。本研究中,我们对CT-K3K7的抗食管癌作用进行了探究,以期为抗食管癌药物的研发提供新的思路和依据。
1 材料和方法
1.1 细胞、主要试剂及仪器
食管癌细胞系KYSE70和KYSE150购自中国科学院上海生命科学研究院细胞库。RPMI1640培养液(Gibco公司);胎牛血清(FBS)、胰蛋白酶、青-链霉素(Gibco公司);CCK-8试剂盒(Invigentech公司);碘化丙啶(propidium iodide,PI)(索莱宝科技有限公司);线粒体膜电位检测试剂盒、活性氧检测试剂盒、Annexin Ⅴ-FITC细胞凋亡检测试剂盒(碧云天生物技术有限公司);PVDF膜(Thermo Fisher Scientific公司);Anti-Bcl-2兔单克隆抗体(Abcam公司);Anti-Bax兔单克隆抗体(Abcam公司);Anti-Caspase-3小鼠单克隆抗体(Santa Cruz公司);BCA蛋白定量试剂、BSA蛋白标准品(康为世纪生物科技股份有限公司);GAPDH抗体(康为世纪生物科技股份有限公司);ECL发光显影试剂盒(Invitrogen公司)。光学显微镜(尼康);凝胶成像系统(BIO-RAD公司);流式细胞仪(BD公司);共聚焦显微镜(ZEISS公司)。
1.2 多肽及细胞培养
多肽CT-K3K7(FLKFLKKLF)由上海吉尔生化有限公司合成,其C端酰胺化,纯度>95%。CT-K3K7以冻干粉形式分装,存放于-20℃以避免反复冻融。在使用时,将其溶解于生理盐水中。KYSE70与KYSE150细胞在含有10% FBS、1%青-链霉素的RPMI1640培养液中,放置于5% CO2、37℃的恒温细胞培养箱中培养。
1.3 细胞增殖实验
利用CCK-8试剂检测CT-K3K7干预后KYSE70与KYSE150细胞增殖的变化[14]。分别取对数生长期的KYSE70及KYSE150细胞,每组设置5个复孔,每孔接种2×103个细胞,待细胞贴壁后进行实验。CT-K3K7的工作浓度分别为5、10、20、40、80 μg/mL,并设置不含有CT-K3K7的对照组。将CT-K3K7按设定的浓度分组加入细胞已贴壁的96孔板中,十字法摇匀,于细胞培养箱中静置培养24 h。每孔加入10 μL的CCK-8溶液,放入培养箱中继续孵育,2 h后用酶标仪测量450 nm处的吸光度(A)值。细胞活力(%)=(A药物组-A空白组)/(A对照组-A空白组)×100%。通过GraphPad软件计算其半抑制浓度(IC50)值,为后续实验确定最终浓度。
1.4 平板克隆实验
利用平板克隆实验进一步检测CT-K3K7对KYSE70与KYSE150增殖能力的影响[15]。分别取对数生长期的KYSE70和KYSE150细胞,按细胞浓度1×106个/mL接种于6孔培养板中。将细胞放置于37°C、5%CO2细胞培养箱中,培养至细胞融合度达80%时进行后续实验。按照前期实验所测定的IC50值,在KYSE70细胞系中选取CT-K3K7的工作浓度为5、10、20 μg/mL;KYSE150细胞系中选取CT-K3K7的工作浓度为10、20、40 μg/mL开展后续实验,且两种细胞系均设置只加PBS的对照组。按照选取的工作浓度,CT-K3K7在6孔培养板中作用细胞6 h。用胰蛋白酶将CT-K3K7干预后的细胞消化,再以每孔2×103个接种于6孔培养板,放置于细胞培养箱中培养,直到6孔板底部肉眼可见大量细胞克隆时终止培养,弃去上清后以PBS洗涤2遍,每孔加入1 mL 40%多聚甲醛,于室温下固定20~40 min,4%结晶紫染色,低倍镜下拍照并计算细胞克隆形成率,进一步评估CT-K3K7对食管癌细胞增殖能力的影响。
1.5 划痕实验
利用细胞划痕实验检测CT-K3K7干预下KYSE70与KYSE150迁移能力的变化[15]。分别取对数生长期的KYSE70和KYSE150细胞,按细胞浓度1×106个/mL接种于6孔培养板。将细胞放置于37°C、5%CO2细胞培养箱中,培养至细胞融合率达80%进行后续实验。按照1.4中CT-K3K7的工作浓度对KYSE70和KYSE150细胞进行分组。CT-K3K7在6孔培养板中作用细胞6 h。用200 μL无菌枪头对各组细胞进行划痕,以PBS洗涤以清除漂浮的细胞,随后将细胞放入37°C、5%CO2培养箱中培养,并于0 h和24 h在低倍镜下拍照,观察细胞迁移面积的变化,评估CT-K3K7对食管癌细胞迁移能力的影响。
1.6 PI吸收测定
利用PI吸收实验探究CT-K3K7是否能够破坏KYSE70与KYSE150细胞的细胞膜完整性[16]。按照选取的实验浓度将CT-K3K7加入处于对数生长期的细胞6孔板中,细胞培养箱内孵育6 h。终止培养后用5 μg/mL PI避光处理10 min,PBS洗涤3次。最后使用共聚焦扫描显微镜检测细胞荧光强度的变化,以评估CT-K3K7对食管癌细胞细胞膜完整性的影响。
1.7 线粒体膜电位(MMP)检测
利用线粒体膜电位检测试剂盒测定CT-K3K7干预后KYSE70与KYSE150的线粒体膜电位的变化[16]。分别取对数生长期KYSE70和KYSE150细胞,按照选取的实验浓度加入CT-K3K7,于细胞培养箱孵育6 h。胰蛋白酶消化细胞,4℃条件下1000×g离心3 min收集细胞。按照试剂盒说明进行检测,计算FL1与FL2比值用以评估CT-K3K7对食管癌细胞线粒体膜电位的影响。
1.8 活性氧(ROS)检测
利用ROS检测试剂盒测定CT-K3K7干预后KYSE70与KYSE150的活性氧水平[17]。分别取对数生长期KYSE70和KYSE150细胞,按照选取的实验浓度加入CT-K3K7,于细胞培养箱孵育6 h。胰蛋白酶消化细胞,4℃条件下1000×g离心3 min收集细胞。按照ROS检测试剂盒说明书进行后续处理,流式细胞仪检测。
1.9 细胞凋亡检测
采用Annexin Ⅴ-FITC细胞凋亡检测试剂盒检测CT-K3K7干预后KYSE70与KYSE150的凋亡水平[18]。分别取对数生长期KYSE70和KYSE150细胞,按照选取的实验浓度加入CT-K3K7,于细胞培养箱孵育6 h。孵育结束后收集上清和细胞于室温下1000×g离心5 min,加入195 μL Annexin Ⅴ-FITC结合液轻轻重悬细胞,再加入5 μL Annexin Ⅴ-FITC以及10 μL碘化丙啶染色液室温避光20 min,立即上机检测细胞凋亡率,分析CT-K3K7对食管癌细胞凋亡水平的影响。
1.10 蛋白质印迹(Western blot)实验
利用Western blot实验检测CT-K3K7干预后KYSE70与KYSE150的相关凋亡蛋白表达量的变化[19]。分别取对数生长期KYSE70和KYSE150细胞,按照选取的实验浓度加入CT-K3K7,于细胞培养箱孵育6 h。孵育结束后利用RIPA蛋白裂解液提取蛋白并经BCA定量试剂盒测定蛋白浓度,将各组蛋白加至电泳槽进行电泳(90 V,100 min);待电泳结束后将蛋白转移至经甲醇激活的PVDF膜(0.2 μm)(80 V,150 min);随后利用脱脂牛奶封闭1 h,TBST洗涤,孵育凋亡相关蛋白抗体Caspase-3(1∶500)、Bcl-2(1∶1000)及Bax(1∶1000),洗涤后孵育相应二抗,最后以ECL显色,于凝胶成像系统分析各蛋白的表达水平。
1.11 统计学方法

2 结果
2.1 蝎毒多肽优化体CT-K3K7抑制食管癌细胞增殖
通过CCK-8法检测CT-K3K7对食管癌细胞KYSE70与KYSE150细胞增殖能力的影响。如图1A和1B所示,KYSE70在CT-K3K7干预下,其生长状况与对照组相比均受到不同程度的抑制,其IC50为17.91 μg/mL。并且随着CT-K3K7作用浓度的升高,CT-K3K7对KYSE70的抑制作用逐渐增强(均P<0.05)。图1C和1D显示,随着CT-K3K7作用浓度的升高,其对KYSE150细胞活力的抑制作用逐渐增强,其IC50为39.52 μg/mL。与对KYSE70细胞的作用类似,CT-K3K7对KYSE150细胞的增殖抑制作用也呈现出明显的剂量依赖性,表明CT-K3K7具有抑制食管癌细胞增殖的作用。
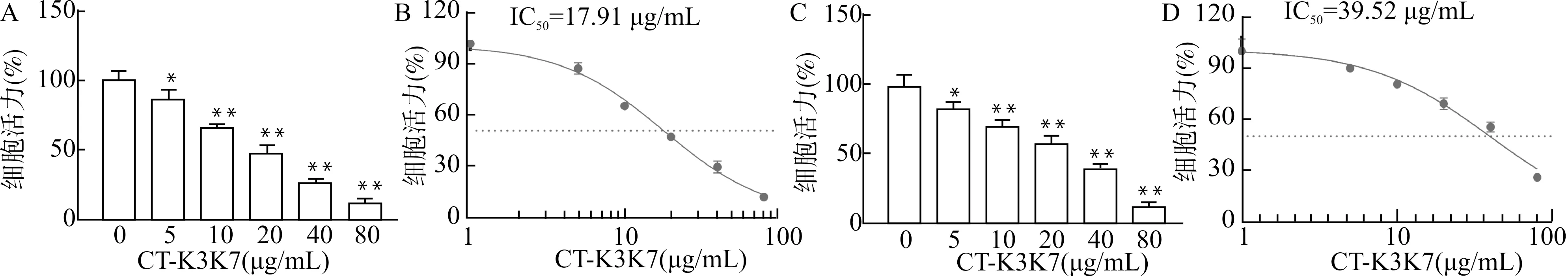
A:KYSE70细胞经不同浓度CT-K3K7处理24 h后的细胞活力;B:CT-K3K7对KYSE70细胞的浓度效应曲线;C:KYSE150细胞经不同浓度CT-K3K7处理24 h后的细胞活力;B:CT-K3K7对KYSE150的浓度效应曲线;与对照组(0 μg/mL组)比较, *P<0.05 ** P<0.01 图1 CT-K3K7对食管癌细胞增殖的影响Fig.1 Effect of CT-K3K7 on proliferation of esophageal cancer cells
2.2 蝎毒多肽优化体CT-K3K7抑制食管癌细胞克隆形成
通过平板克隆实验进一步检测CT-K3K7对食管癌细胞KYSE70与KYSE150细胞增殖能力的影响。如图2所示,CT-K3K7处理组的细胞克隆形成率均低于对照组,且随着作用浓度的升高CT-K3K7对KYSE70(图2A、2B)与KYSE150(图2C、2D)克隆形成的抑制作用逐渐增强,呈显著剂量依赖效应(均P<0.05)。

A:KYSE70细胞平板克隆形成实验图;B:不同浓度CT-K3K7处理后KYSE70细胞克隆形成率;C:KYSE150细胞平板克隆形成实验图;D:不同浓度CT-K3K7处理后KYSE150细胞克隆形成率;与对照组(0 μg/mL组)比较, * P<0.05 ** P<0.01图2 CT-K3K7对食管癌细胞体外克隆形成能力的影响Fig.2 Effect of CT-K3K7 on the clonogenic ability of esophageal cancer cells in vitro
2.3 蝎毒多肽优化体CT-K3K7抑制食管癌细胞迁移能力
通过细胞划痕实验检测CT-K3K7对食管癌细胞迁移能力的影响。结果如图3所示,与对照组相比,经过CT-K3K7处理后的KYSE70(图3A、3B)与KYSE150(图3C、3D)细胞的迁移能力随着CT-K3K7作用浓度的升高明显降低,呈现显著剂量依赖效应(均P<0.01)。
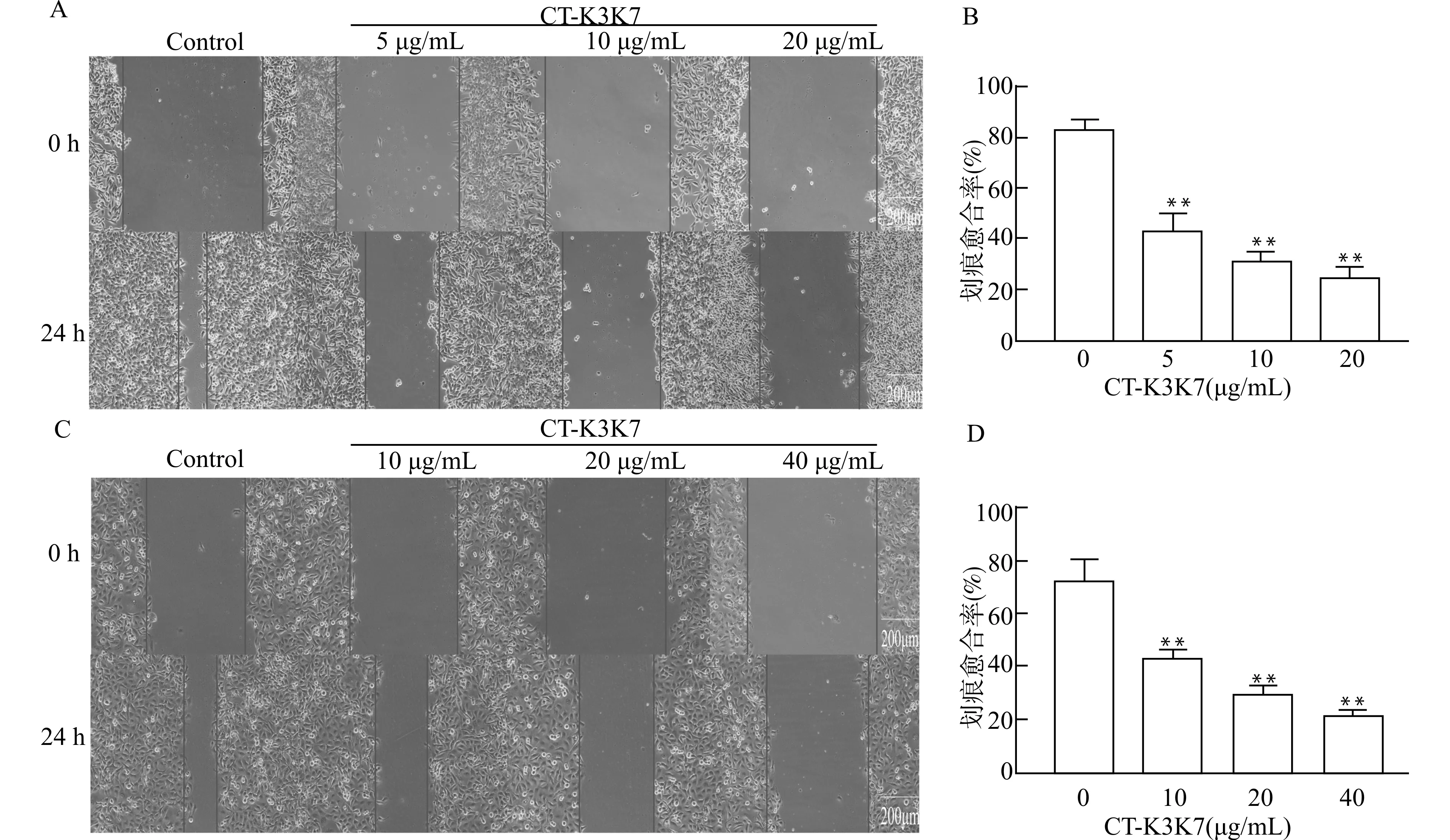
A:KYSE70细胞划痕实验;B:KYSE70细胞划痕实验统计图;C:KYSE150细胞划痕实验;D:KYSE150细胞划痕实验统计图;与对照组(0 μg/mL组)比较,**P<0.01图3 CT-K3K7对食管癌细胞体外迁移能力的影响Fig.3 Effect of CT-K3K7 on the migration ability of esophageal cancer cells in vitro
2.4 蝎毒多肽优化体CT-K3K7破坏食管癌细胞膜完整性
为探究CT-K3K7是否能够破坏细胞膜,通过对CT-K3K7处理后的KYSE70与KYSE150细胞进行PI染色以检测细胞膜的完整性。如图4所示,与对照组相比,CT-K3K7干预后的KYSE70与KYSE150细胞随着多肽作用浓度的升高,荧光染色阳性细胞数逐渐增多,与对照组差异明显。以上结果表明,CT-K3K7能够破坏食管癌细胞的细胞膜完整性,这一作用与CT-K3K7浓度呈正相关。同时也提示CT-K3K7可能对食管癌细胞具有直接杀伤作用。
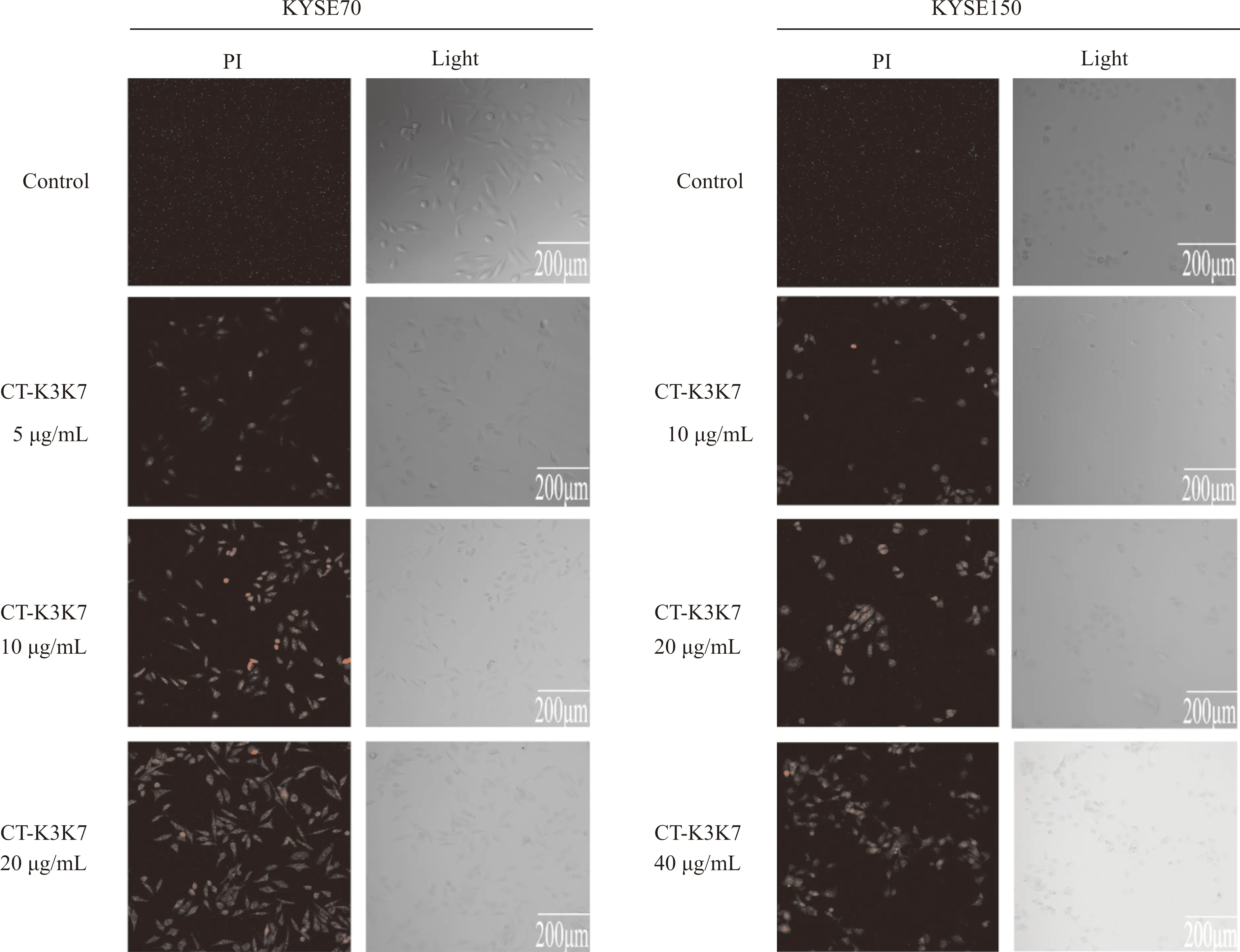
图4 CT-K3K7对食管癌细胞细胞膜通透性的影响Fig.4 Effect of CT-K3K7 on the cell membrane permeability of esophageal cancer cells
2.5 蝎毒多肽优化体CT-K3K7引起食管癌细胞线粒体功能障碍
为验证CT-K3K7对食管癌细胞KYSE70与KYSE150线粒体功能的影响,采用流式细胞术检测CT-K3K7干预后食管癌细胞KYSE70与KYSE150线粒体膜电位与ROS水平的变化。结果显示,与对照组相比,CT-K3K7干预后的KYSE70(图5A、5B)与KYSE150细胞(图5C、5D)的线粒体膜电位显著下降(均P<0.05);经CT-K3K7干预后,KYSE70(图5E、5F)与KYSE150(图5G、5H)细胞内ROS水平明显升高(均P<0.05),并且两者都呈显著的浓度依赖性。
2.6 蝎毒多肽优化体CT-K3K7引起食管癌细胞凋亡
采用Annexin Ⅴ-FITC/PI检测CT-K3K7对KYSE70与KYSE150细胞凋亡的影响,并通过Western blot检测凋亡相关蛋白表达水平。结果如图5I、5L所示,经CT-K3K7干预后的食管癌细胞凋亡水平显著高于对照组,且与CT-K3K7浓度呈正相关(均P<0.05)。凋亡相关蛋白Caspase-3、及Bax/Bcl-2比值是评价细胞凋亡水平的重要指标,Western blot结果(图6)显示,与对照组相比,CT-K3K7处理的KYSE70与KYSE150细胞Bcl-2蛋白表达量随着CT-K3K7浓度升高显著下调(均P<0.05),而Bax、Caspase-3蛋白表达量则随着CT-K3K7作用浓度的升高显著上调(均P<0.05),Bax/Bcl-2比值明显升高(均P<0.05)。该结果进一步表明CT-K3K7能够诱导食管癌细胞凋亡,并且这一效应具有显著的剂量依赖性。

A:KYSE70细胞蛋白印迹实验代表性图片;B:KYSE70细胞中Caspase-3蛋白表达量;C:KYSE70细胞中Bcl-2蛋白表达量;D:KYSE70细胞中Bax蛋白表达量;E:KYSE70细胞中Bax与Bcl-2的比值;F:KYSE150细胞蛋白印迹实验代表性图片;G:KYSE150细胞中Caspase-3蛋白表达量;H:KYSE150细胞中Bcl-2蛋白表达量;I:KYSE150细胞中Bax蛋白表达量;J:KYSE150细胞中Bax与Bcl-2的比值;与对照组(0 μg/mL组)比较,* P<0.05 **P<0.01图6 CT-K3K7对食管癌细胞Caspase-3、Bax、Bcl-2表达的影响Fig.6 Effect of CT-K3K7 on the expression of Caspase-3,Bax,and Bcl-2 in esophageal cancer cells
3 讨论
多肽是一类具有良好开发前景的药物候选分子,对病毒、细菌、癌症、真菌均表现出良好的活性,且不易产生耐药性[20-21]。蝎子作为宝贵的传统药物资源[22-25],其毒液是发掘抗癌多肽的良好资源。目前已经从多个蝎品种中发掘出许多具有抗癌作用的蝎毒多肽,如来自东亚钳蝎的蝎毒肽BmKn-2对人口腔癌与结直肠癌细胞具有杀伤作用[26-27]、来自Vaejovismexicanus的VmCT1对乳腺癌的抗癌特性[28]和来自Scorpiomauruspalmatus的蝎毒肽Smp24与Smp43对人肺癌与肝癌的抗肿瘤作用[29-31]。蝎毒多肽优化CT-K3K7是基于西藏特里豚蝎抗微生物多肽Ctry2459的改造肽,具有较好的体内外抗菌作用[13]。
癌细胞的增殖能力在肿瘤的发展过程中发挥着关键的作用,抗癌多肽如Eucalrobusone C[32]、Carnosic acid[33]、Cordycepin[34]在抗肿瘤的过程中可通过干预细胞的增殖能力来发挥作用。本研究证实CT-K3K7对食管癌细胞增殖具有良好的抑制作用,且具有浓度依赖性(图1)。为进一步探究CT-K3K7对食管癌细胞增殖的影响,本研究探究了CT-K3K7对KYSE70与KYSE150细胞克隆形成的影响。结果表明,CT-K3K7可以显著抑制KYSE70与KYSE150细胞的克隆形成(图2),进一步说明CT-K3K7有效地抑制了KYSE70与KYSE150细胞的增殖。癌转移是肿瘤患者死亡的主要原因,而癌细胞迁移则是癌转移的重要初始步骤[35],抗癌多肽如Smp43、Cecropins、与Smp24通过抑制肿瘤细胞的迁移能力[29,31,36]来干预癌症的发展。为此,本研究进一步探究了CT-K3K7对KYSE70与KYSE150细胞的迁移能力的影响。结果表明,CT-K3K7对KYSE70与KYSE150迁移能力具有显著的抑制性,并有显著的浓度依赖效应(图3)。对于抗癌多肽来说,其通常可以直接与癌细胞细胞膜相互作用,通过与膜脂质相互作用,引起细胞膜结构的改变,破坏细胞膜的完整性从而起到直接杀伤癌细胞的作用,如抗癌多肽HPRP-A2[16]、Smp24[37]和CT-p19LC[38]。本研究通过PI吸收检测表明CT-K3K7能够对KYSE70与KYSE150细胞膜完整性造成破坏(图4),并且造成KYSE70与KYSE150活力显著下降,这表明CT-K3K7可通过破坏食管癌细胞膜直接杀伤食管癌细胞。对抗癌多肽而言,破坏细胞膜完整性并不是其唯一的抗肿瘤机制,其能够通过与线粒体作用,破坏线粒体结构和功能导致线粒体功能障碍,从而诱导细胞线粒体途径凋亡[39-40],如抗癌多肽Isoorientin[41]、CSEI[42]、Bombesin related peptides[43]等皆具有此效应。而线粒体膜电位去极化和ROS富集是线粒体功能障碍的重要表现。本研究通过进一步的流式细胞检测发现CT-K3K7引起KYSE70与KYSE150细胞内线粒体膜电位的去极化(图5A~5D)与ROS水平上升(图5E~5H)并且导致KYSE70与KYSE150的凋亡率的上升(图5I~5L)。Bcl-2与Bax是凋亡分子机制研究的主要靶分子,而Bax和Bcl-2的促凋亡及抗凋亡作用相互制衡,通常在体内可通过复杂的机制调控细胞内的稳态平衡,但当细胞处于异常应激状态时,出于细胞本能反应,往往伴随Bax表达水平提高的同时Bcl-2的表达受到抑制进而打破稳态介导细胞凋亡[44]。进一步的结果表明,经CT-K3K7干预后的食管癌细胞凋亡相关蛋白Caspase-3及促凋亡蛋白Bax表达量均显著升高,同时伴随抗凋亡蛋白Bcl-2表达量明显降低(图6)。由此可见,CT-K3K7不仅可通过破坏细胞膜完整性,直接杀伤食管癌细胞,还可以通过介导食管癌细胞线粒体功能障碍来诱导食管癌细胞凋亡,从而发挥抗癌作用。
综上所述,CT-K3K7可以有效抑制食管癌细胞KYSE70与KYSE150的增殖并具有浓度依赖性,并抑制其迁移能力。CT-K3K7可通过破坏细胞膜直接杀伤食管癌细胞,也可以导致食管癌细胞线粒体功能障碍最终引起食管癌细胞发生凋亡。因此,蝎毒多肽优化体CT-K3K7具有作为抗肿瘤分子药物的潜力并为研发抗食管癌药物提供了新的候选分子或分子模板。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
现代园艺(2017年13期)2018-01-19 02:28:09
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
西南军医(2016年1期)2016-01-23 02:22:29
中国继续医学教育(2015年4期)2016-01-07 07:38:06
川北医学院学报(2015年5期)2015-12-05 08:22:43
药学与临床研究(2015年4期)2015-06-05 11:35:54
科学中国人(2015年16期)2015-02-28 09:14:02
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
