
白杨无性系叶片水力及经济性状与生物量的关系*
2023-11-05 12:59侯卓梁姜在民
林业科学 2023年10期
王 薇 赵 涵 黄 欣 侯卓梁 姜在民 蔡 靖,3
(1. 西北农林科技大学林学院 杨凌 712100;2. 西北农林科技大学生命科学学院 杨凌 712100;3. 陕西秦岭森林生态系统国家野外科学观测研究站 杨凌 712100)
植物的生长离不开自身光合能力和水分传输能力的驱动。叶片既是植物形成光合产物的主要场所(金鹰等,2015),也是水分传输的终端部分,调节气孔导度开放,影响碳水交换(Sacket al.,2006;Fichotet al.,2011;Scoffoniet al.,2016)。叶片的水力性状和经济性状可体现上述两大功能特性。叶水力性状是指叶片在水分运输方面的性状(潘莹萍等,2014;金鹰等,2015),包括叶片水力导度、叶脉密度、导管直径、气孔长度、气孔密度等(潘莹萍等,2014;Liet al.,2015;Yinet al.,2018;Liuet al.,2019);叶片经济性状量化了经济型谱(LES)(Wrightet al.,2001;2004),用来表征叶片对碳的获取策略,主要包括叶片碳、氮含量、比叶面积、光合和呼吸速率等(Reich,2014;金鹰等,2015;Jinet al.,2016;Yinet al.,2018)。叶片水力性状和经济性状反映了植物对水-碳资源的获取能力和权衡策略,在一定程度上影响生物量的积累,探究二者与生物量的关系,对理解植物的存活与生长至关重要。
杨树(Populussp.)具有生长快、适应性强等特点,是我国重要的造林树种。生物量的高低是选育优良杨树无性系的关键因素(Marronet al.,2006),可通过一些简单易测的叶片性状来预测无性系生物量(Bunnet al.,2004;Dillenet al.,2009)。在叶水力性状中,大多数研究关注气孔性状(气孔长度和气孔密度)与杨树生物量的关系,但其结果因杨树种类和基因型而异(Alafaset al.,2006;Moncluset al.,2006;Dillenet al.,2008)。叶片水力导度和叶脉网络结构(叶脉密度、导管直径)通过控制叶片气体交换能力(气孔导度)对生物量进行调控(Nardiniet al.,2005;Scoffoniet al.,2016),但它们与生物量的关系在杨树种内和种间存在不确定性。因此,将水力性状指标进行综合考虑,探讨其与杨树无性系生物量的关系,对了解植物叶片水分供需对生物量的影响具有重要价值。叶经济性状中,总叶面积和单叶面积与植物生物量密切相关(Moncluset al.,2005;Gebaueret al.,2016;Baiet al.,2020)。比叶面积和叶片数量与生物量之间有无相关性,则主要取决于物种、生长条件和植株年龄等(Raeet al.,2004;Zhanget al.,2004;Marronet al.,2005)。可见,即使是同一父母本杨树无性系在不同环境或者时间内,能够预测其良好生物量的叶经济性状指标也会有所不同。
由于叶水力性状和经济性状都参与水分和二氧化碳的交换过程(Yinet al.,2018),虽对于二者关系的研究较多,但其研究结果不一。此外,二者关系的研究多是在种间水平与跨自然环境梯度的背景下开展(Sacket al.,2003;Liet al.,2015;Jinet al.,2016;Yinet al.,2018),而在遗传背景差异更小的基因型水平且基于同质园试验的研究较为欠缺,少有研究综合考虑二者与生物量关系,而这对探明植物生长是二者协同作用的结果还是各自独立驱动的结果具有重要意义。
本研究以来源于同一父母本[Populus alba‘I-101’ ×(P. alba × P. glandulosa) ‘84K’]产生的8 个具有不同生长速率的白杨无性系为对象,通过测定叶水力性状、叶经济性状和地上部分生物量,分析无性系间生物量、叶水力性状和经济性状的差异,并探究叶水力性状与经济性状的关系,以及解析二者与生物量的关系,以期探寻能够预测高产白杨无性系地上生物量的早期生理指标,为无性系的选育提供理论与实践指导。
1 研究区概况
研究地点位于西北农林科技大学苗圃(34°15′N,108°03′E),地处关中平原中部,属半湿润半干旱地区,暖温带大陆性季风气候。土壤类型为沙壤土。年平均气温12.9 ℃,无霜期221 天,年平均降水量635 mm,年平均蒸发量1 000 mm。
2 材料与方法
2.1 试验材料
试验选择白杨杂交产生的8 个无性系为材料,即K1、K2、K3、Z1、Z2、Z3、M1、M3,无性系在2017 年的胸径由大到小排序为K1>K2>K3>Z1>Z2>Z3>M1>M3。2009 年这8 个无性系的母树种植于西北农林科技大学渭河试验站。至2017 年2 月(8 年生),每个无性系母树剪取长约25 cm 的插穗,扦插在位于陕西杨凌西北农林科技大学苗圃同质园中。所有扦插无性系苗管理措施基本一致,不施加肥料,生长季去除杂草,生长依靠自然降雨,仅在无性系幼苗阶段的伏旱天气进行灌溉。2020 年7—9 月(4 年生)在每个无性系中选取6 棵生长状况良好、代表总体平均生长水平的植株,于每棵植株冠层顶部采集1 根当年生枝条,选取该枝条的3 个较大叶片,测定其水力性状、经济性状。除叶片维管水力导度是测定无性系侧枝叶片而得(方法详见2.2.1.1),每个无性系的其他叶片指标数据均为上述所测定的18 个叶片的平均值。
2.2 试验方法
2.2.1 叶片水力性状的测定 叶片水力导度通常由叶片维管水力导度和非维管水力导度2 部分组成。叶片维管水力导度表征主要在叶脉中运输的水分;叶片非维管水力导度表征主要穿过维管束、海绵、栅栏等叶肉组织,最后从气孔扩散的水分(Sacket al., 2006;Trifiloet al., 2021)。研究叶水力导度和维管水力导度,有助于判断影响生物量积累的关键。
1)叶片维管水力导度(KleavesHPFM)的测定 采用高压液流计法(high pressure flow meter,HPFM)(Yanget al.,1994;Sacket al.,2002):每个无性系选择6 棵生长状况良好的植株,每棵植株选取1 根基部直径至少为1 cm 的向阳侧枝,将其放入黑色塑料袋并带回实验室复水1 夜。于第2 天取出枝条,在水下用刀片将枝条基部修整光滑,连接至HPFM 装置中,以7 kPa s-1的速率加压至500 kPa,之后维持压力稳定。HPFM 会每隔8 秒自动记录1 次水流阻力值。待一定时间后,观察到叶片背面有水排出,且阻力值比较稳定时,记录此时阻力值为枝条的阻力(Rshoot)。随后,剪掉枝条上的所有叶片和叶柄,再次等待阻力值稳定,其稳定值为剪掉叶片和叶柄的阻力(R-leaves),并测定所有叶片面积(AL)。KleavesHPFM是叶片的水力阻力(Rleaves)的倒数(Scoffoniet al.,2012),并用AL标准化。计算公式如下:
式中:Rleaves为水力阻力(MPa s·kg-1);Rshoot为剪掉叶片和枝条后阻力(MPa s·kg-1);R-leaves为叶柄阻力(MPa s·kg-1);KleavesHPFM为叶片维管水力导度(kg·m-2s-1MPa-1);AL为叶面积(m2)。
每个无性系选取6 根来自不同植株上的带叶片的侧枝来测定叶片维管水力导度,取平均值作为最终结果。
2)叶片水力导度(维管和非维管KleafEFM)的测定采用蒸发通量法(Evaporative flux method,EFM)(Sacket al.,2002):在上述每个无性系选取的6 棵植株中,于每棵树的冠层顶部选取1 个当年生向阳枝条,在傍晚采集后迅速放入装有湿纸的黑色塑料袋中,立即带回实验室复水1 夜,使叶水势接近于0。于第2 天将复水后的枝条放入超纯水中,切取1 片无病虫害的带叶柄的大叶片,将叶柄迅速连接至EFM 装置中。待叶片蒸腾至少30 min 流速稳定后记录叶温,蒸腾速率(E)为最后读取的5 个数据的平均值,温度矫正为25 ℃。测定完成后迅速取下叶片,放入提前充满水蒸气的塑封袋中(以防止叶片蒸腾失水),并放置于黑暗中平衡30 min 后测定叶水势(Ψleaf)(Scoffoniet al.,2008;2016),最后将叶片放入根系扫描仪上(Epson v750,J221A,Japan)上进行扫描,用WinRHIZO 软件(Regent Instruments Inc.,Canada)测量叶面积(LA),KleafEFM的计算公式如下,之后用LA 标准化。
式中:KleafEFM为叶片水力导度(mmol·m-2s-1Mpa-1);E为蒸腾速率(mmol ·s-1)Ψleaf为叶水势(-MPa);LA为叶面积(m2)。
2.2.2 叶脉密度(VD)的测定 VD 指单位面积内叶脉的总长度(Brodribbet al.,2007)。将每个叶片平整地放于根系扫描仪上进行扫描。使用WinRHIZO软件对扫描图片沿主脉在上、中、下3 个不同位置绘制测定区域(含次脉),进行VD(mm·mm-2)的测定,最后求取平均值代表1 个叶片的VD 值。
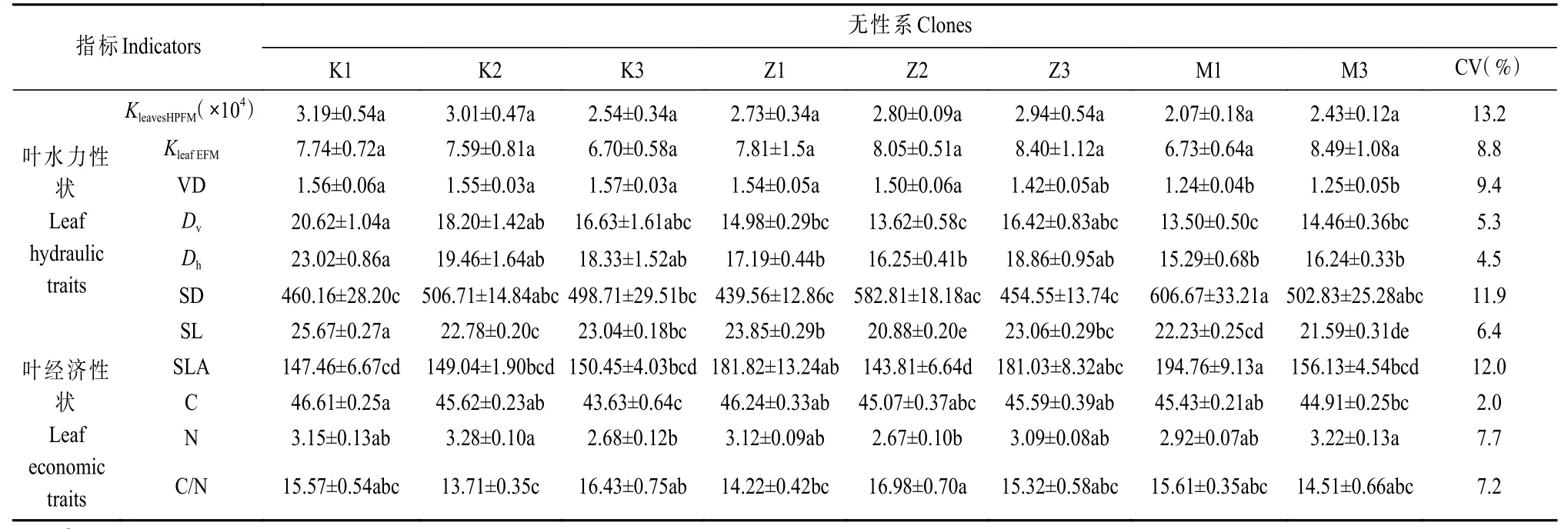
表1 8 个白杨无性系叶片水力和经济性状特征值①Tab. 1 Values of the leaf hydraulic and economic traits of the eight poplar clones
2.2.3 导管直径(Dv)和导管水力直径(Dh)的测定将扫描完的叶片沿主脉从叶尖至叶基部分为上、中、下3 个区域,在每个区域切取1 个1 cm×1 cm 的正方形小块,放入FAA 固定液中[甲醛∶冰醋酸∶70%酒精 (V∶V∶V)= 1∶1∶18],于4 ℃下保存,制石蜡切片(李和平,2009)。每个叶片制作3 个样片,并在正置荧光显微镜(LeicaDM4000B,Germany)100×或200×下观察,当所有导管出现在视野时拍照,使用WinCell Pro version 2012a(Regent Instruments Inc.,Canada)图像分析软件,测定图像中每个导管腔面积(AV),导管数目(N),计算Dv、Dh。其计算公式为:
式中:Dv为导管直径(µm);AV为导管腔面积(µm2);N为导管数目;Dh为导管水力直径(µm)。
2.2.4 气孔性状的测定 气孔性状主要包括气孔密度(SD,mm-2)和气孔长度(SL,µm),SD 为单位面积内的气孔数量;SL 为保卫细胞的长度(Sacket al.,2003)。气孔性状采用指甲油印迹法制片(Russoet al.,2010),每个叶片制作3 个样片,使用正置荧光显微镜在400×下观察,并选择3 个视野进行拍照。之后,用Image J软件统计每张照片中的气孔数量,求取SD。接着在每张照片中选取4 个清晰的气孔进行SL 的测定。
2.2.5 叶片经济性状的测定 叶片经济性状主要包括比叶面积(SLA),叶片全碳(C)、全氮(N)含量及碳氮比(C/N)。使用直径为1.5 cm 的打孔器从每个叶片上(避开中脉)取5 个小圆片,记录其叶面积(LA,cm2),后放入烘箱,在65 ℃下烘至恒重,测定其干质量(DW,g),求取SLA,SLA=LA / DW。将测定过干质量的叶片,经研钵磨碎、过筛后,用元素分析仪(vario MACRO cube,elementar)测定C、N 含量和C/N 值(Marronet al.,2006)(无性系叶片数量选取见2.1)。
2.2.6 地上部分生物量的测定 在生长季结束,树木开始落叶时(2020 年10 月20 日)测定所有无性系距地面15 cm 处的地径(D)(精确至0.01 cm),每个无性系测定30 株,再使用前期研究已建立的异速生长方程(与本试验材料相同)(Zhaoet al.,2021)计算地上部分生物量(AGB),并取平均值作为最终结果。具体公式如下:
式中,AGB 为地上各部分生物量(kg);D为距地面15 cm处的直径(cm)。
2.3 数据处理
利用Excel 2013 对所有数据进行初步分析处理,使用SPSS 21.0 软件对数据进行Shapiro-Wilkinson 残差正态分布检验和Levene’s 方差齐性检验,对不符合的数据进行对数转换,以符合参数检验要求。采用单因素方差分析(ANOVA)检验无性系间指标的差异性;皮尔逊相关分析(Pearson)的一元线性和多元线性回归分析无性系叶片水力和经济性状与生物量的关系和其影响大小;主成分分析阐明三者间耦联关系。利用Origin 2019b 软件绘制相关图表。
3 结果与分析
3.1 无性系间生物量、叶水力性状和经济性状的差异
8 个无性系AGB 大小排序为:K1>K2>K3>Z2>Z1>M3>Z3>M1,最大值比最小值高出96.3%(图1)。无性系M1 的AGB 显著小于其他无性系;无性系Z3的AGB 显著小于Z1、Z2,而与M3 无显著差异性。叶水力性状在无性系间的变异系数大小排序为:KleavesHPFM>SD>VD>KleafEFM>SL>Dv>Dh。KleavesHPFM的变异系数最大,为13.2%,变化范围为2.07~3.19 kg·m-2s-1MPa-1;Dh的变异系数最小,为4.5%,变化范围为15.29~23.02 µm(表1)。叶片经济性状指标的变异系数大小排序为:SLA>N>C/N>C。SLA 的变异系数最大,为12.0%,变化幅度达到1.35 倍。C 的变异系数最小,为2.0%。N 和C/N 的变异系数比较接近,分别为7.7%和7.2%。相比叶经济性状的变异范围(2.0%~12.0%),水力性状(4.5%~13.2%)在无性系间变异较大。差异分析表明,叶水力性状指标KleavesHPFM和KleafEFM在无性系间无显著差异,其他指标均差异显著;叶经济性状指标在无性系间均差异显著(表1)。
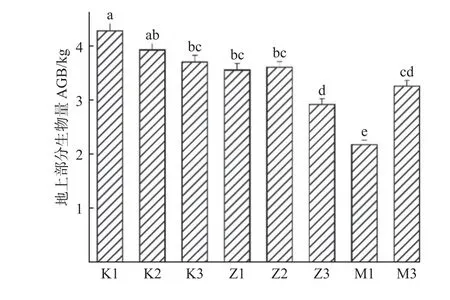
图1 8 个无性系间地上部分生物量(AGB)Fig. 1 Above-ground biomass of eight clones (AGB)
3.2 叶水力性状与经济性状的关系
无性系叶片水力性状和经济性状无显著相关性(P>0.05)(表2)。叶水力性状中,KleavesHPFM与VD、Dv、Dh呈显著正相关(P<0.05),SL 与Dv和Dh呈显著正相关(P<0.05)。叶片经济性状中N 与C/N 呈极显著负相关(P<0.01),其余指标间均无显著相关性(P>0.05)(表2)。

表2 叶片水力及经济性状间相关系数①Tab. 2 Pearson’s coefficient between leaf hydraulic and economic traits of clones
3.3 叶片水力及经济性状与生物量的关系
叶片水力性状中的KleavesHPFM、VD、Dv和Dh与AGB 均呈显著正相关性(P<0.05),(图2a, c, d, e),KleafEFM、SD 和SL 与AGB 均无显著相关性(P>0.05)(图2b, f,g);叶经济性状中的SLA 与AGB 呈极显著负相关(P<0.01)(图3a),C、N 和C/N 与AGB 均无显著相关性(P>0.05)(图3b, c, d)。
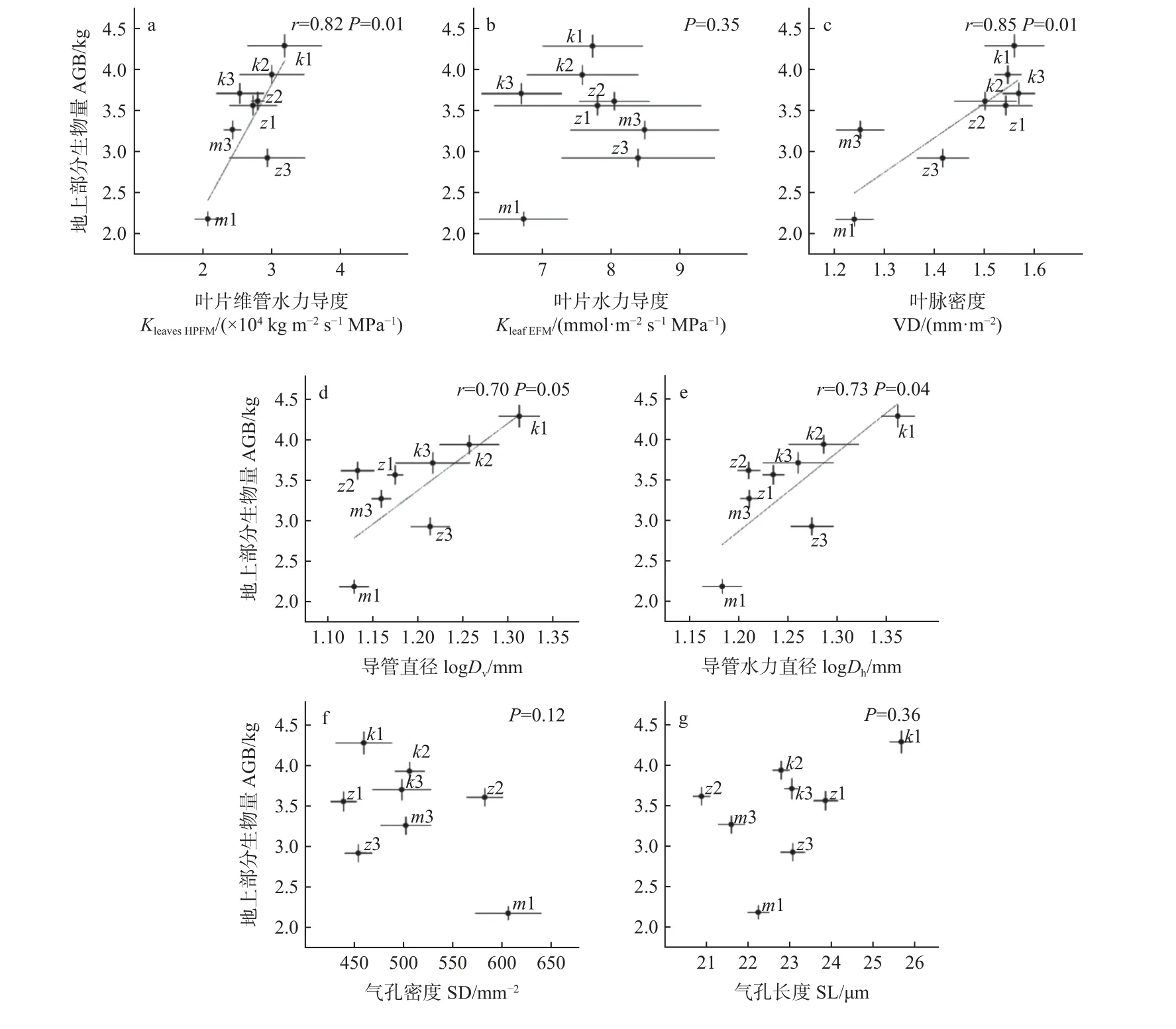
图2 叶片水力性状与地上部分生物量(AGB)的关系Fig. 2 Relationship between leaf hydraulic traits and above-ground biomass (AGB)
利用多元线性回归分析了叶水力及经济性状与生物量的关系,发现VD 和SLA 的回归方程可解释AGB 变异的87.6%,且达到显著水平(P<0.05)。其中,VD 对AGB 的影响最大(t=2.957)(表3)。

表3 无性系地上部分生物量多元线性回归①Tab. 3 Multiple linear regression of above-ground biomass of clone
对所有性状进行主成分分析,提取了2 个主成分,共解释71.1%的变异,第一主成分和第二主成分分别解释49.0%和22.1%的变异(图4)。第一主成分主要与AGB 和叶水力性状(KleavesHPFM、Dv、Dh)有关,而第二主成分主要与叶经济性状(SLA、C、N、C/N)有关(表4),表明无性系叶片水力性状和经济性状解耦,无性系AGB 与大部分叶水力性状具有相关性,说明无性系AGB 受叶片水力性状的影响较大。
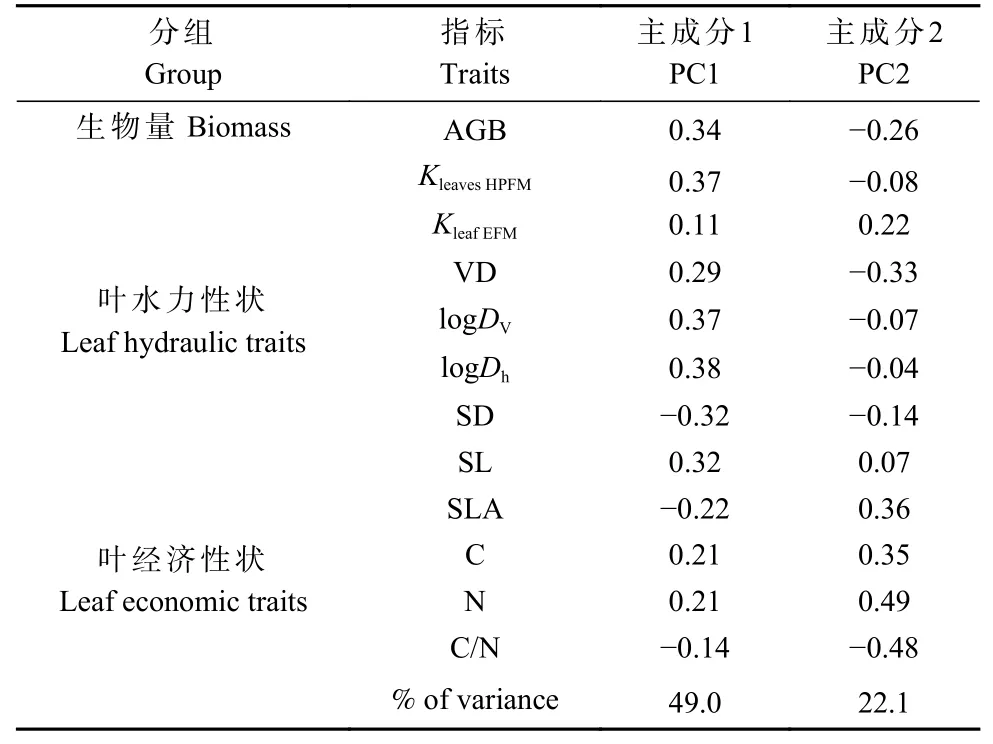
表4 主成分分析中不同叶功能性状的载荷Tab. 4 Loading scores of leaf functional traits in the PCA
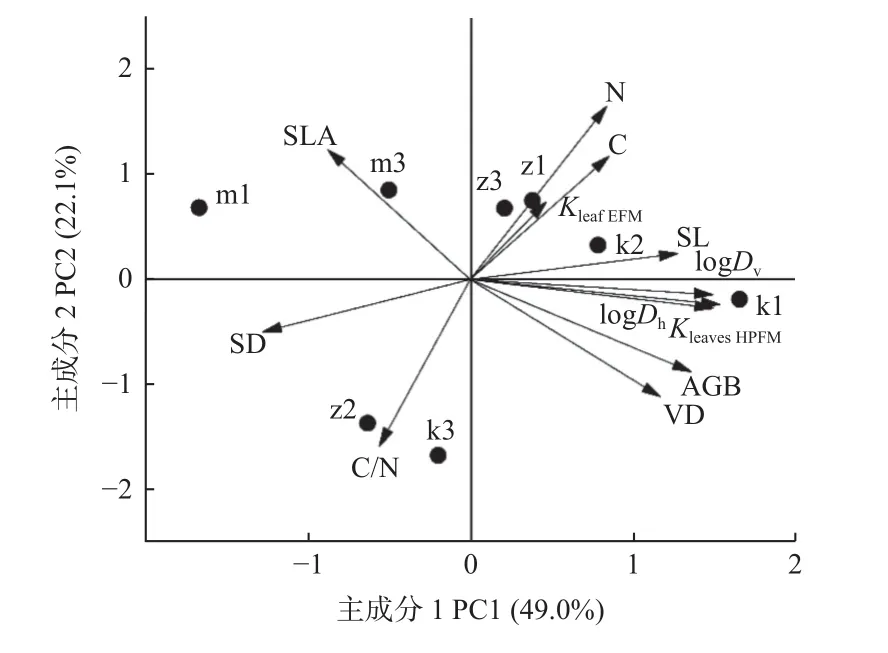
图4 无性系间生物量和叶功能性状的主成分分析Fig. 4 Principal component analysis of biomass and leaf functional traits among 8 clones
4 讨论
4.1 无性系间生物量、叶水力性状和经济性状的差异
试验所用无性系在扦插后3 年间(2017—2019 年)的生长中表现出了地上部分生长速率和生物量积累的差异(Zhaoet al.,2021),本研究于2020 年生长季开始试验,测得的所有无性系的AGB 排序与前3 年一致,且大部分无性系的生长速率表现与8 年生的母树一致。虽然部分无性系,如M1 和M3,与8 年生的母树生长差异有起伏,但总体来说,仍低于K1、K2、K3以及Z1、Z2。由于所有无性系均生长在环境条件基本一致的同质园内,这些结果说明无性系间的AGB差异是由遗传因素决定的,具有时间稳定性。
无性系间叶片性状的显著差异是选育高产基因型的前提(Gebaueret al.,2016)。本研究无性系间具有显著差异的叶片性状包括,水力性状中的VD、Dv、Dh、SD 和SL 以及经济性状中的SLA、C、N 含量和C/N。Gleason 等(2016)发现被子植物的不同物种中,当VD 低于5~10 mm·mm-2时,VD 与Kleaf和gs之间有很强的协调性。本研究无性系间VD 值均在2 mm·mm-2以下,且VD 与KleavesHPFM呈正相关。因此,它可能与全孔导度(gs)有关,影响植物生长。无性系叶的Dv和Dh值均在13~23 um 之间,比茎的Dv和Dh值小(25~35 um)(Zhaoet al.,2021)。狭窄的叶导管直径不容易形成栓塞,降低了茎木质部功能发生障碍的风险(Lechthaleret al.,2020),保护了植物生长。无性系SD 和SL 的变异幅度(分别为:440~607 mm-2,21~26 um)与不同亲本杨树无性系相比(Yu,2001;Alafaset al.,2006;Fichotet al.,2011),SD 值受基因的影响较大,相差3 倍左右;但SL 值比较稳定,均在25 um 左右波动。
叶片经济性状中,叶片SLA、C、N 含量、C/N 的变异幅度(分别为:144~195 cm2·g-1,44%~47%,2.7%~3.3%,14~17)与自然条件下生长的其他品种杨树无性系的指标范围保持一致(Moncluset al.,2005;Guetet al.,2015)。但C 和C/N 的变异幅度比温室中生长的P. deltoides×P. nigra黑杨无性系的指标变异幅度(分别为:46%~48%,16~25)略小(Marronet al.,2006),而N 和SLA 的变异幅度比温室中(分别为:1.9%~2.9%,84~118 cm2g-1)的略大。这可能是因为本研究无性系生长主要依靠自然降水,而温室中无性系进行了充足的水分灌溉,说明除受基因差异外,环境水分差异也可能是造成2 种杨树叶经济性状差异的主要原因。
前人研究认为某些叶片性状可用于反映杨树基因型的高产潜质(Bunnet al.,2004;Marronet al.,2005;Gebaueret al.,2016)。本研究揭示了同一父母本的子代无性系叶水力性状和经济性状存在差异,且这两类性状与无性系地上生物量之间均存在显著相关性,此外无性系间稳定的生长趋势,使在无性系生育早期选定预测高产性能的叶片性状成为可能。
4.2 无性系叶水力性状与经济性状的解耦
由于叶水力性状和经济性状都参与水分和二氧化碳的交换过程,有人认为这2 组性状是紧密联系的(Nardiniet al.,2012;Jinet al.,2016;Yinet al.,2018)。然而,本研究无性系叶片水力性状和经济性状不相关,此结果也与前人在种间水平上的研究结果一致(Liet al.,2015;Liuet al.,2020)。Li 等(2015)从理论上分析叶片水力性状和经济性状的解耦主要与相关叶片解剖结构在物理位置上的分离有关:叶脉主要分布在栅栏组织和海绵组织中间,气孔主要分布在下表皮,因此水力性状大多与海绵组织有关;而叶经济性状(如N 含量)主要与栅栏组织有关。本研究发现,叶片水力性状指标KleavesHPFM、Dv、Dh与海绵组织厚度(ST)呈显著正相关;叶经济性状指标SLA 与栅栏组织厚度(PT)和叶片厚度(T)呈显著负相关,其他指标与解剖结构均无显著相关性。所以大部分叶片水力性状与经济性状确实与解剖结构关联的组织位置不同,符合Li 的假设。然而,叶片水力性状中VD 比较特殊,VD与SLA 无直接关系,但二者均与PT 有联系。有研究表明,叶脉密度与叶片构建成本无关(Liet al.,2015;Kawaiet al.,2017;Kawaiet al.,2020)。VD 与PT 呈正相关,可能是因叶脉密度越高,叶片水分运输和储存能力也就越大,越有利于栅栏叶肉保持良好的水分状况,从而提高光合能力(Thomaset al.,2015)。而SLA与PT 呈负相关,意味着SLA 越小的植物,叶片干质量越多,具有更厚的叶片或更密的叶肉组织(Milla-Morenoet al.,2016)。综上,本研究认为白杨无性系叶片水力性状和经济性状的解耦确实与叶片解剖结构具有一定的关系,水力性状主要与海绵组织有关,叶经济性状与栅栏组织有关。此结论是否具有普遍性仍需要更多种间及种内的试验证据。
4.3 无性系叶水力及经济性状的变异对生物量的驱动作用
叶片水力性状中,KleavesHPFM在无性系间的变异系数CV 值是KleafEFM的1.5 倍,KleavesHPFM与AGB 呈显著正相关,而KleafEFM与AGB 无相关性。由于KleavesHPFM和KleafEFM的主要区别是有无计算叶片非维管水力导度,二者与AGB 的关系不一致可能是叶非维管部分的水力导度对AGB 的影响造成的。前人研究指出叶片水分运输的阻力主要存在于非维管路径(Cochardet al.,2004),并且叶片水力导度与植物气体交换过程密切相关(Sacket al.,2006;Xionget al.,2020)。但近年来对于叶片不同水流路径与气体交换关系的研究发现,叶片非维管水力导度会削弱叶片水力导度与气孔导度(gs)的协调,且自身与gs的关系比叶维管水力导度与gs的关系弱(Liuet al.,2019)。因此,叶维管水力导度可能通过影响植物与外界气体交换从而影响AGB,但由于该指标在无性系间无显著差异,所以无法作为本研究中的预测指标。
然而,叶维管路径的导水效率通常与叶脉网络结构(VD、Dv、Dh)有关,如较大的叶脉密度VD 使叶片有较多的水分运输通道,同时减少了叶脉末梢到蒸腾位置的距离,提高了叶片的水分运输能力(Brociouset al.,2016);导管是植物长距离水分运输中重要的木质部细胞(Scholzet al.,2013),Dv和Dh越大,越能满足叶片的水分供应(龚容等,2015)。本研究的结果同样支持了此观点,即KleavesHPFM与叶脉网络结构的3 个指标(VD、Dv、Dh)均呈显著正相关。此外,VD、Dv、Dh均与AGB 呈显著正相关,其中VD 是影响最大的因子。由驱动叶片气体交换的水力理论模式图(Scoffoniet al.,2016)可知,VD 主要通过KleavesHPFM对气孔导度gs和光合速率(Amax)产生影响,进而影响生物量积累。
另外,虽然无性系间SD 和SL 有显著差异,且一般SD 和SL 的相互作用影响气孔导度大小(Frankset al.,2009),调节生物量积累。但本研究未发现SD 和SL 与AGB 有关。Dillen 等(2008)和Monclus 等(2006)对黑杨无性系的研究也得到了一致的结果。然而也有研究认为SD 和SL 与植物生长有关,例如Alafas 等(2006)发现12 个不同亲本和杂交群的杨树无性系SD 和SL 与生物量呈显著正相关;Yu(2001)发现P.tremula×P. tremuloides的生长与SD 呈负相关。因此,SD 和SL 与杨树无性系生长的关系,受杨树亲本的影响较大。
叶经济性状中,SLA 与AGB 呈显著负相关,而C、N 含量、C/N 与AGB 无相关性。Marron 等(2005)对P. deltoids×P. nigra无性系研究表明:在温室生长的3 月龄生根插条其生物量与SLA 呈负相关,也与N 呈负相关;Marron 和Ceulemans(2006)在第二年的研究中发现该杨树无性系生物量与叶片C、N 含量无相关性,但并没有发现其与SLA 的相关性。此外,在对田间生长的同种杂交杨无性系(P. deltoids×P. nigra)的研究中发现,不管是2 年生还是3 年生的无性系均与SLA 无相关性(Moncluset al.,2005;Gebaueret al.,2016)。可见,叶经济性状与生物量之间有无相关性,主要取决于物种、生长条件和植株年龄等。虽然本试验对象是白杨无性系,SLA 与AGB 有关,但还需延长试验研究时间,来进一步验证预测指标的可靠性。
从PCA 和载荷表中分析可知,PC1 主要与水力性状相关,PC2 主要与经济性状相关,水力性状和大部分经济性状关系不紧密。另外,AGB 主要与水力性状在一个轴上,而与大部分经济性状解耦,说明早期杨树无性系生长差异主要是由水资源获取策略的不同造成的。
5 结论
白杨无性系具有稳定的生长趋势,且叶片水力性状和经济性状在无性系间存在差异,使在无性系生育早期选定某一生理指标作为预测高产指标成为可能。叶水力性状与经济性状不相关,但二者均与无性系生物量相关,二者可能是驱动白杨无性系生长的独立的两个轴。大多数叶水力性状指标与生物量密切相关,叶片维管水力导度、叶脉密度、导管直径和导管水力直径越大的无性系,生物量越高。叶片经济性状中,只有比叶面积与生物量有相关性,比叶面积越小的无性系,生物量越高。相比比叶面积,叶脉密度是该无性系主要的预测因子。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
动漫界·幼教365(大班)(2021年11期)2021-11-20
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
发明与创新(2020年26期)2020-06-27
装备制造技术(2019年12期)2019-12-25
小学生学习指导(低年级)(2019年9期)2019-09-25
科技创新与品牌(2019年12期)2019-02-06
