
莱州湾芙蓉岛人工鱼礁区大型底栖动物的营养结构特征
2023-11-05 08:46董建宇张宇洋詹启鹏张宗航申丰源丁夏阳张沛东张秀梅
水产学报 2023年9期
张 镇,董建宇,孙 昕,张宇洋,詹启鹏,张宗航,申丰源,丁夏阳,张沛东,张秀梅
(1.中国海洋大学海水养殖教育部重点实验室,山东 青岛 266003;2.浙江海洋大学水产学院,浙江 舟山 316022)
食物网及营养结构关系是海洋生态系统研究的重要内容,是研究各物种间营养关系的重要途径。大型底栖动物是海洋食物网中至关重要的生态类群,在海洋底栖食物网中扮演着重要角色,对研究食物网中各物种营养结构和关系具有重要作用[1-2]。研究海洋生物食性及食物网营养关系的方法主要有胃含物分析法、碳氮稳定同位素法、特征脂肪酸组成分析、特定化合物同位素分析技术、基于高通量测序的DNA 条形码技术以及基于理论模型的食物网结构预测方法等[3-4]。其中,碳氮稳定同位素法不仅能够反映近一段时间内生物摄食的食物类型,而且能够追溯生物长期生命活动的主要营养来源,具有反映时间长、准确度高等优点,现已成为海洋动物营养生态位和食物网结构研究的重要手段,被国内外学者广泛应用[5-10]。
大型底栖动物作为人工鱼礁底栖生境的重要组成部分,在食物网物质循环和能量流动中发挥重要作用,其营养关系会直接影响人工鱼礁的生态效应。刘春云等[11]利用稳定同位素技术研究了烟台养马岛潮间带大型底栖动物食物网结构特征。刘晓收等[12]利用碳氮稳定同位素技术,对黄海大型底栖动物的食物源、营养级和底栖食物网结构进行了研究,并初步构建了黄海底栖食物网。Lee 等[13]分析了韩国江华潮滩不同生境中大型底栖动物及其潜在食物来源的碳氮稳定同位素比值,对其底栖食物网动态变化进行了研究。以上这些研究揭示了底栖动物的食物网结构,此外针对海洋牧场及人工鱼礁区食物网及营养关系也有部分研究[14-18],但鱼礁建设时间的长短对大型底栖动物的营养关系和生态效应是否具有直接的影响,目前仍不明确。
因此,实验利用碳、氮稳定同位素分析法对莱州湾芙蓉岛海域人工鱼礁区大型底栖动物食物网进行研究,探讨主要底栖动物种类及其潜在食源的碳、氮稳定同位素特征;分析不同潜在碳源对底栖动物的食物贡献率;研究不同底栖动物在食物网中的营养级关系,以期为评价人工鱼礁的生态效应提供基础数据。
1 材料与方法
1.1 研究区域
研究区域位于烟台莱州太平湾海域明波海洋牧场,该区域平均水深3~4 m。目前该海域已建成人工鱼礁区4 处,投放人工鱼礁约35.47 万空方。其中礁区A 和礁区B 为3 年礁龄的礁区 (建设时间为2018 年5—8 月),礁区C 和礁区D 为7~8 年礁龄的礁区 (建设时间为2013—2014 年)。本研究将礁区A 和礁区B 合并归为短礁龄区 (ARAB)、将礁区C 和礁区D 合并归为长礁龄区 (ARCD)。每处礁区内设置3 个采样点,另按照我国人工鱼礁建设技术规范 (SCT 9 416—2014)在礁区附近200~1 000 m 设置3 个对照点。采样站点如图1 所示。
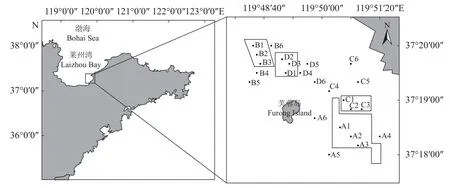
图1 人工鱼礁区底栖动物调查站位示意图A1~A3、B1~B3 为短礁龄区点位,A4~A6、B4~B6 为短礁龄区对照点位;C1~C3、D1~D3 为长礁龄区点位,C4~C6、D4~D6 为长礁龄区对照点位。Fig.1 Sampling sites of macrobenthos in artificial reefsA1-A3,B1-B3 are short-age reef area sites,A4-A6,B4-B6 are short-age reef area control sites;C1-C3,D1-D3 are long-age reef area sites,C4-C6,D4-D6 are long-age reef area control sites.
1.2 样品采集与处理
于2021 年4 月对芙蓉岛海域人工鱼礁区进行取样调查,采集大型底栖无脊椎动物样品,并同时采集其潜在食源样品,包括浮游植物、浮游动物、沉积物有机物 (SOM)、水体悬浮颗粒有机物(POM)用于稳定同位素分析。大型底栖动物使用面积为0.05 m2的抓斗式采泥器进行取样,每个站位重复取样3 次,用0.5 mm 网筛淘洗收集底栖动物,样品经冷冻后带回实验室,用体视显微镜(Nikon SMZ1000,日本)进行鉴定,通过WoRMS网站 (http://marinespecies.org/)对鉴定出的物种名录进行校对和复核。样品的处理、鉴定和称重严格按照《海洋调查规范》(GB/T 12763.6—2007)进行。浮游动植物分别使用大、小型浮游生物网采用水平拖网方式来采集。使用抓斗式采泥器采集SOM,收集表层1~2 cm 的泥样作为待测样品。POM 通过使用水样瓶采集水样进行收集。
样品在实验室内的处理方法参考刘晓收等[12]的研究报道,但本实验将其方法中用浓盐酸酸熏改为用0.1~1.0 mol/L 的稀盐酸进行酸化,以得到更好的处理效果,具体过程:在实验室内,浮游植物样品使用100 目筛绢过滤掉样品中的浮游动物和悬浮碎屑,然后将浮游植物样品抽滤到0.45 μm 孔径的GF/F 膜上 (提前450 °C 预燃6 h 以除去无机碳)。将滤膜使用0.1~1.0 mol/L 的稀盐酸 (下同)进行酸化,之后烘干并装入锡囊,用于碳氮稳定同位素含量的测定;POM 样品处理方法与浮游植物相同。浮游动物样品将其静养24 h 排除消化道内含物,用去离子水清洗,再使用稀盐酸进行酸化,之后烘干并装入锡囊,用于碳氮稳定同位素含量的测定。SOM 样品烘干24 h 后,取0.5 g于10 mL 离心管中,加入1 mol/L 的稀盐酸酸化之后再次烘干并装入锡囊中,用于碳氮稳定同位素含量的测定。底栖动物样品经鉴定种后用于同位素样品前处理:多毛类以及较小个体的类群选取整体,双壳类选取闭壳肌,腹足类选取腹足部分,甲壳类等选取肌肉组织。将动物组织用去离子水清洗后,放入-80 °C 冷冻干燥机中冷冻干燥48 h,使用酸化后的玻璃棒将冻干后的样品研磨成粉状,接下来进行酸化处理,将1 mol/L的稀盐酸逐滴加入样品管中,直至样品停止产生CO2,反应后水洗至中性,将样品烘干48 h,再次研磨装入锡囊用于碳氮稳定同位素含量测定。
1.3 碳氮稳定同位素的测定
所有样品的碳氮稳定同位素比值使用同位素质谱仪进行测定。碳、氮稳定同位素比值用国际通用的δ值表示,分别以VPDB (Vienna Pee Dee Belemnite)国际标准和大气氮作为参考标准。δ13C、δ15N 分别按以下公式计算:
式中,13C/12CVPDB为国际标准物VPDB 的碳同位素比值,15N/14Nair为标准大气氮同位素比值。
1.4 营养级及食源贡献率计算
δ15N 值的高低可以确定消费者在食物网中的营养级,通过下列公式计算:
式中,TL 代表消费者的营养级,δ15N消费者代表消费者的氮稳定同位素比值,δ15N基准代表基准生物的氮稳定同位素平均比值。λ代表所选取基准生物的营养级,基准生物为初级生产者时,λ=1;基准生物为初级消费者时,λ=2。Δδ15N 代表营养级间的氮稳定同位素富集水平,平均值约为3.4‰。本实验选取在人工鱼礁区分布较为广泛且频繁出现的浮游动物作为基准生物计算大型底栖动物的营养级[19]。
利用基于R 程序的Bayesian 稳定同位素混合模型 (SIAR)计算不同饵料生物的贡献率:经过同海域研究所得营养富集因子校正 (δ13C 和δ15N 值的营养富集因子分别为0.4‰±1.3‰和3.4‰±1.0‰)拟合得出摄食来源[20]。由于在消费者组织中测量的稳定同位素比值与其食物中的值紧密相关,即稳定同位素生态位可能与营养生态位紧密相关,因此可以用消费者的稳定同位素值来描述它们生态位的变化:利用R 语言SIBER 包对比底栖动物群落食物网结构和营养冗余程度:①δ15N 范围(NR)提供了群落营养长度的信息;②δ13C 范围(CR)用以估计基础资源的多样性;③包含数据点(TA)的凸包的总面积,以指示生物对营养生态空间的利用情况;④ 到中心点的平均距离 (CD)提供了关于生态位宽度和物种间距的额外信息;⑤ 物种聚集度密度参数 (MNND)提供群落内物种密度和聚集度的信息;⑥ 物种聚集度均匀度参数(SDNND)给出了空间密度和填充均匀性的度量[21]。本实验过程中操作人员严格遵守动物实验伦理规范,并按照相关的规章制度执行。
对上述公式的计算和数据处理通过Microsoft Excel、SPSS 22.0、OriginPro 9.0 和R 3.3.2[22]等软件完成。各数值以平均值±标准差 (mean±SD)表示。
2 结果
2.1 碳、氮稳定同位素特征
潜在食源碳、氮稳定同位素特征 由于在人工鱼礁区内未发现大型海藻 (草)的存在,所以实验主要采集了POM、SOM、浮游植物和浮游动物等4 种潜在有机碳源样品。潜在食源的δ13C和δ15N 值稳定同位素值见表1。经单因素方差分析 (ANOVA),对照区潜在食源SOM 的δ13C 值显著高于人工鱼礁区 (P<0.05),而礁区之间则无显著差异,其余3 种潜在食源之间的δ13C 值在不同区域均无显著差异。潜在食源SOM 的δ15N 值在短礁龄区显著低于长礁龄区和对照区 (P<0.05),POM 的δ15N 值在长礁龄区显著低于对照区(P<0.05),其余δ15N 值在不同区域间无显著差异。
经ANOVA 方差分析发现,在整个研究区域中4 种潜在食源之间的δ13C (F=29.686,P<0.001)和δ15N (F=99.836,P<0.001)存在极显著差异:SOM和浮游植物的δ13C 稳定同位素平均值相近 (浮游植物为-17.98‰±1.98‰;SOM 为-18.92‰±3.40‰),并高于POM (-26.12 ‰±0.47 ‰)和浮游动物(-25.23‰±0.84‰)。浮游动植物的δ15N 稳定同位素平均值最高 (浮游动物为9.56‰±0.57‰,浮游植物为9.30‰±0.32‰),最低值为POM (5.12‰±0.73‰),SOM 平均δ15N 值居中 (5.93‰±0.59‰)。
消费者碳、氮稳定同位素特征 实验以食性为依据,将大型底栖动物分为包括浮游生物食者 (planktonophagous group,Pl)、肉食者 (carnivorous group,C)、碎屑食者 (detritivorous group,D)和杂食者 (omnivorous group,O)在内的4 类摄食功能群,并分析了它们之间的关系 (图2,表2)。经过分析,浮游生物食者δ13C 值在长礁龄区中(-16.97‰±0.00‰)显著高于短礁龄区 (-23.08‰±0.89‰) (P<0.05),不同区域间浮游生物食者δ15N值差异不显著。此外,其余3 个类群的碳、氮稳定同位素值在不同区域间均无显著差异。
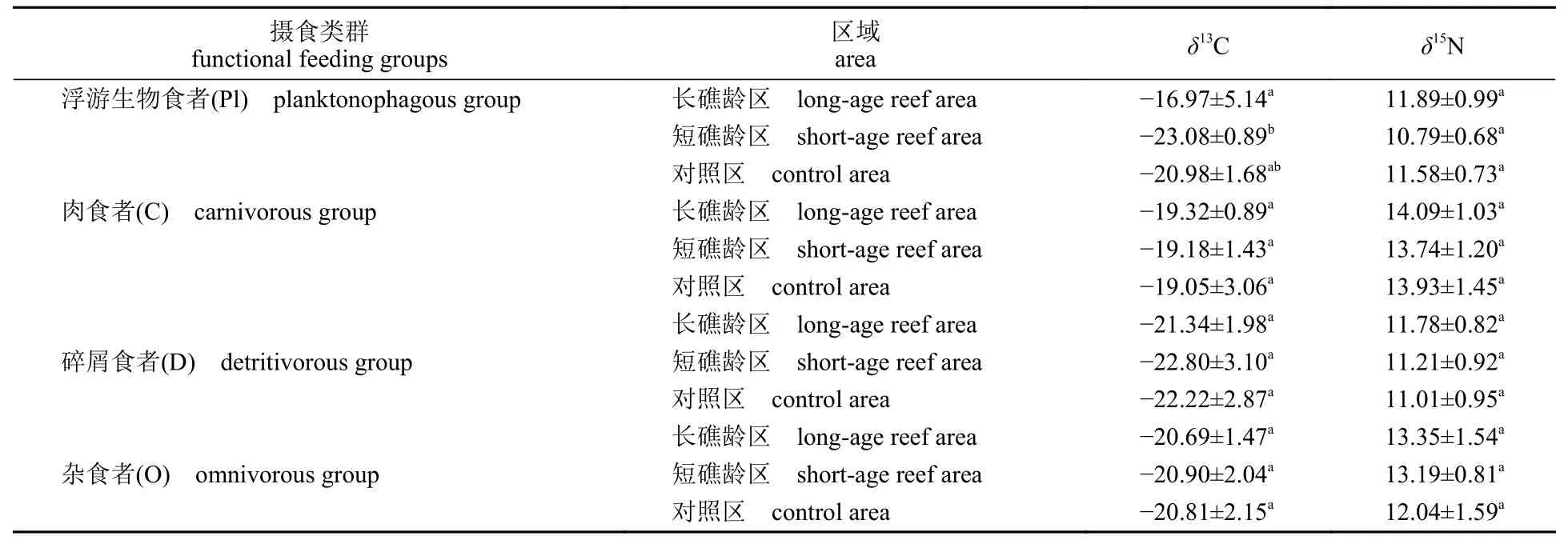
表2 不同区域间不同摄食类群的碳氮稳定同位素特征Tab.2 Carbon and nitrogen stable isotope characteristics of different functional feeding groups in different areas
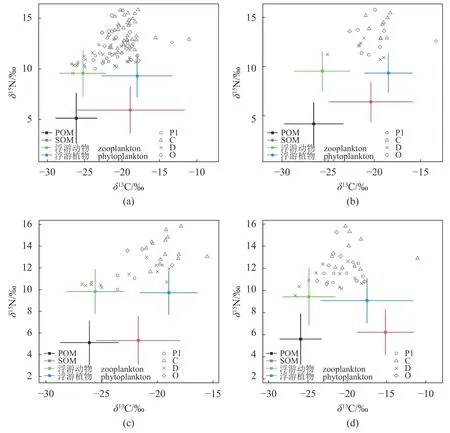
图2 芙蓉岛大型底栖动物摄食功能群和潜在食源碳氮稳定同位素双标图(a) 总体,(b) 长礁龄区,(c) 短礁龄区,(d) 对照区,Pl.浮游生物食者,C.肉食者,D.碎屑食者,O.杂食者,下同。Fig.2 Biplot of carbon and nitrogen stable isotope values of each functional feeding groups of macrobenthos and their potential feeding sources in Furong Island(a) overall,(b) long-age reef area,(c) short-age reef area,(d) control area,Pl.planktonophagous group,C.carnivorous group,D.detritivorous group,O.omnivorous group,the same below.
ANOVA 方差分析发现,在整个研究区域中,消费者的δ13C 在4 种摄食类群中存在极显著差异 (F=9.233,P<0.001)。C 类群的平均δ13C 值 (-19.18‰±1.91‰)高于其余3 个类群 (Pl,-20.84‰±2.86‰;D,-22.26±2.77‰;O,-20.78‰±1.82‰);同时统计了4 类摄食功能群动物δ15N 的变化情况,采用ANOVA 方差分析将大型底栖动物分为4 组:C 类群的平均δ15N 值 (13.73‰±1.23‰)最高,Pl和D 类群最低 (Pl,11.44‰±0.79‰;D,11.27‰±0.92‰),O 类群平均δ15N 值 (12.67‰±1.54‰)居中 (F=27.208,P<0.001),表明肉食者类群在底栖生境中具有最高的氮富集值,是处于较高营养级的一个类群。
2.2 不同区域食物网结构和营养冗余程度对比
利用R 语言SIBER 包对3 个不同区域内底栖动物碳氮稳定同位素进行分析,各区域底栖动物同位素特征见图3,参数见表3。结果显示,营养结构生态量化指标在不同区域之间存在一定差异:短礁龄区底栖动物群落食物网营养长度(NR)最长(2.95)。长礁龄区底栖动物群落中食源多样性(CR)最高 (4.36),其次为短礁龄区,对照区最低。底栖动物对营养生态空间利用 (TA)方面,长礁龄区数值最高 (5.77),对照区利用程度最低 (0.56)。长礁龄区与短礁龄区平均营养级多样性 (CD)相当且高于对照区 (1.12)。在营养冗余程度方面,物种聚集度密度参数 (MNND)长礁龄区 (2.01)高于短礁龄区和对照区,物种聚集度均匀度参数(SDNND)在人工鱼礁区与对照区相近。
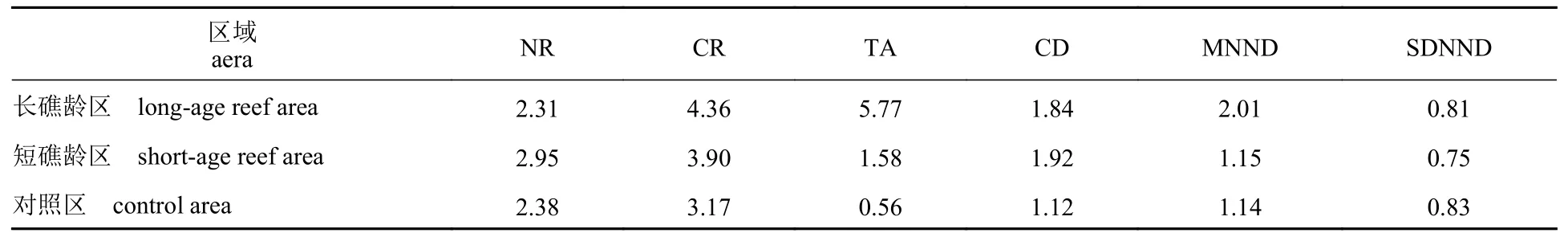
表3 不同区域生物群落指标Tab.3 Biological community indices in different areas
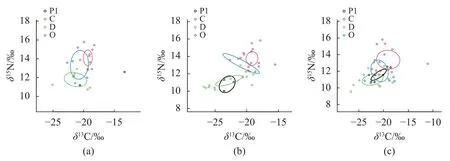
图3 不同区域摄食功能群贝叶斯图(a) 长礁龄区,(b) 短礁龄区,(c) 对照区。Fig.3 Bayesian plot of functional feeding groups in different regions(a) long-age reef area,(b) short-age reef area,(c) control area.
校正的标准椭圆面积 (SEAc)表征核心同位素生态位,其数值的高低象征着营养生态位宽幅的宽窄。不同摄食功能群对资源利用情况如表4 所示,在长礁龄区,O 的营养生态位宽幅较大,其次为D,最低为C,且O 与C 生态空间相互重叠明显。在短礁龄区,D 营养生态位宽幅最大,且与其余3 个类群生态空间相互重叠。在对照区,C 营养生态位宽幅最大,其次为O 和D 类群,Pl类群营养生态位宽度最小,且4 个类群的生态空间重叠明显,说明在对照区中底栖动物的营养关系差异较小,营养生态空间利用情况较为相似。

表4 不同摄食功能群生态位利用情况Tab.4 Utilization of ecological niche of the different functional feeding groups
2.3 潜在食源对大型底栖动物摄食功能群食物贡献率
近岸水域生态系统中的消费者,其潜在碳源主要为浮游动植物、沉积物有机物、悬浮颗粒有机物、大型海藻、海草等。由于芙蓉岛人工鱼礁海域大型海藻和海草缺失,本研究在计算潜在碳源对消费者的贡献率时,只考虑浮游动物、浮游植物、SOM 和POM 等4 种潜在食源。潜在食源对人工鱼礁区底栖动物消费者摄食功能群的平均贡献率如图4 所示。
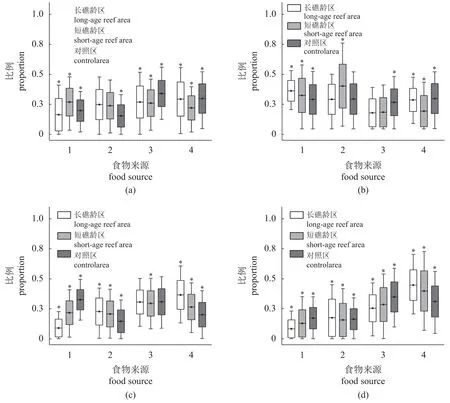
图4 四类摄食功能群食物来源及比例(a) 浮游生物食者(Pl);(b) 肉食者(C);(c) 碎屑食者(D);(d) 杂食者(O)。1.POM,2.SOM,3.浮游动物,4.浮游植物。*表示差异显著。Fig.4 Food sources and proportions of four functional feeding groups(a) planktonophagous group;(b) carnivorous group;(c) detritivorous group;(d) omnivorous group.1.POM,2.SOM,3.zooplankton,4.phytoplankton.* indicates significant difference.
将Pl、D 以及O 类群以浮游动物、浮游植物、SOM 和POM 作为潜在食源进行食物贡献率分析(图4)。在长礁龄区,Pl 类群中POM 占比最低(18.7%),其余3 种潜在食源贡献率大致相同。浮游植物碳源对D 类群的贡献率最高 (36.5%),浮游动物 (30.9%)与其相近,SOM (22.7%)占比居中,POM 占比最小 (9.9%)。O 类群中浮游植物碳源占比最高(44.5%),POM 最低(8.4%)。在短礁龄区,Pl 类群中4 种潜在食源贡献率大致相同,POM 最高(27.2%)。浮游动物碳源对D 类群的贡献率最高(29.5%),最低为SOM (21.7%)。O 类群中浮游植物占比最高(40.6%),POM 最低(13.3%)。在对照区,Pl 类群潜在食源贡献率最高为浮游动物(33.9%),SOM 占比最低(16.2%)。O 类群中,浮游动物碳源贡献率最高为34.8%,最低为SOM(16.2%)。综上所述,随着人工鱼礁建设时间的变化,长礁龄区除C 类群之外,其余3 种摄食功能群的食物贡献率中均为浮游植物碳源占比最高,贡献最少的均为POM。短礁龄区中,浮游植物碳源不再占比最高,但仍占据较大比例。对照区中,4 种摄食类群的食物来源与人工鱼礁区之间具有较大差异。
C 类群由于食源特殊,因此以Pl、D 和O 类群为潜在食源计算食物贡献率 (图4-b)。在长礁龄区,潜在食源贡献最高为Pl 类群 (33.37%),最低为O 类群 (16.04%)。在短礁龄区,D 类群贡献率最高,为37.84%,最低为O 类群 (16.22%)以及Pl 类群 (17.43%)。在对照区,4 种潜在食源贡献率大致相同。总体上,C 类群在长礁龄区更多的以Pl 类群为潜在碳来源,短礁龄区中C 类群更多以底栖D 类群为食,同时对照区中C 类群摄食O类群的比例相比于鱼礁区有所提高。
2.4 大型底栖动物连续营养谱
以浮游动物为基线生物,基于营养级计算公式测定了芙蓉岛人工鱼礁区主要底栖动物的营养级,并基于此构建了该海域大型底栖动物的连续营养谱图 (图5)。长礁龄区的营养级范围为2.00(浮游动物)~3.83 [(颗粒拟关公蟹(Paradorippe granulata)]。其中位于2~3 营养级上的动物种类最多,有13 种,占总量的59.1%;营养级高于3 的物种有8 种。短礁龄区的营养级范围为2.00 (浮游动物)~3.49 [岩虫 (Marphysa sanguinea)],其中位于2~3 营养级上的动物种类最多,有13 种,占总量的65.0%;营养级高于3 的物种有6 种。对照区的营养级范围为2.00 (浮游动物)~3.87 [头吻沙蚕 (Glycera capitata)],位于2~3 营养级上的动物种类最多,有15 种,占总量的68.2%;营养级高于3 的物种有6 种。研究表明,在人工鱼礁区与对照区高营养级的底栖动物中以C 和O 类群为主。
3 讨论
本研究选取了POM、SOM、浮游植物和浮游动物作为人工鱼礁区大型底栖动物的潜在食物来源。POM 是食物网中重要的物质与能量来源,它是一个非均质有机质库,主要由浮游植物和其他水体碎屑组成[23],本研究中不同区域POM 的碳稳定同位素值在空间上无显著差异,且与中国近海各海域 (如黄海灵山湾海域[24]、舟山群岛枸杞岛海域[25]、大亚湾海域[26]、天津大神堂海域[18]等)相比均偏低,推测可能是由于研究区域离岸较近且属于内湾,水交换能力较弱,受陆源物质的影响较大,从而导致POM 较低的δ13C 值[27-28]。SOM来源比较复杂,如POM 的自然沉降、大型海藻(草)以及海洋生物死亡后形成的碎屑沉积等[11]。本研究中SOM 的δ13C 值与浮游植物相似,表明SOM 的有机质可能主要来源于浮游植物等的沉降[29-30]。此外,SOM 在人工鱼礁区与对照区之间的碳稳定同位素值具有显著差异,这可能是由于人工鱼礁建设引起礁体及礁区周围水流的变化,进而影响礁区沉积物的组成和有机营养物质的沉积效率[2,23,31-32]。该人工鱼礁区海域浮游植物的δ15N 值偏高,推测可能是由于在莱州湾等人类活动密集、富营养化的沿海海域,浮游植物可能获得较高的人为来源的δ15N 同位素所导致[30,33]。通常,碳稳定同位素可以用来指示生物种的食物来源和组成,本研究中潜在食源的碳、氮稳定同位素值在不同类群以及不同区域的同一类群间具有差异,也由此说明了本研究区域底栖动物食物来源的复杂性和多样性,此外不同摄食类群间的碳、氮稳定同位素差异更多的是与其摄食习性相关,而不是与其分类关系或生境特征有关[23]。
本研究中,长礁龄鱼礁区CR 值、TA 值以及MNND 水平均高于短礁龄区和对照区,这意味着该区域底栖动物群落营养生态位更广,食物网中占据相似营养生态位的动物比例更小,营养冗余程度变低,这表明长礁龄区底栖动物群落营养多样性增加,群落内物种具有更加多样的营养生态位,群落结构稳定性较好[34-36],这可能是由于随着投放礁体时间的延长,长礁龄区底栖生境生物资源增加,食源多样性水平有所提升,从而使得占据同种营养生态位的底栖动物比例减小,种间竞争强度降低,最终促进群落稳定性的提高。同时短礁龄区和对照区食物来源多样性水平较低,进而使处于相同生态位物种数量增加,最终导致群落营养冗余程度的增加[37]。此外,短礁龄区和对照区间的MNND 和SDNND 值相近,表明在建设时间较短的人工鱼礁区与对照区,底栖动物群落的营养状况与营养冗余的关系趋于一致[38]。人工鱼礁的生态功能主要在于改善底栖生境,促进生物群落结构改善和生物量提高。研究发现,在人工鱼礁区与对照区之间,不同摄食功能群所占据的生态空间发生变化,说明人工鱼礁建设影响到了底栖生境的食物网结构及营养关系。Zhang等[39]的研究也证实了这一点,即人工鱼礁建设会引起鱼类及无脊椎动物等营养水平的变化。
浮游植物是重要的海洋初级生产者,也是海洋食物网的基础碳来源之一[40-41]。本研究利用稳定同位素混合模型 (SIAR)得出,浮游植物碳源对3 类消费者的平均贡献率保持在31.40%以上,为该区域主要碳源供应者,并且在人工鱼礁区中3类消费者食物中的浮游植物碳源比例有所上升,说明浮游植物碳源作为基础的食物来源在人工鱼礁区得到了更加充分的利用,也表明在人工鱼礁建设之后海域的初级生产力有所提高,这与多处海域研究报道的浮游植物碳源是消费者主要碳来源的结果相同[15-16,42]。浮游动物作为海洋的初级消费者,在食物链中起着关键的作用,它能将营养物质输送到较低的营养级并保证较高营养级的捕食者能够获得食物,因此在海洋生态系统食物网中占据重要地位[43-45]。有研究表明,芙蓉岛海域人工鱼礁建设使得浮游动物丰度和群落结构稳定性均有显著提升,有效促进了该海域生产力水平的提高[46]。本研究中浮游动物碳源在整个鱼礁区的贡献率较对照区略有提升也证明了这一观点。此外,肉食者类群在3 个不同研究区域之间食物来源的变化,也说明人工鱼礁的建设改变了底栖生境食物网的营养结构和能量的传递链。随着礁龄的增加,浮游生物食者承担了更多食物链的能量传递过程,反映出人工鱼礁的生态效应在礁龄上的差异。采用碳、氮稳定同位素混合模型研究食物网的主要能量和物质来源及动态,对揭示食物网能量和物质循环变动规律具有重要意义。本研究中4 类摄食功能群消费者食物来源在鱼礁区与对照区以及在长、短礁龄鱼礁区之间都存在差异,说明底栖动物的食物来源及组成受到了人工鱼礁建设的影响,但由于海洋生态系统有机物质来源复杂,影响基础碳源同位素值的因素较多,因此在进行食物来源以及贡献率研究时还需综合各种因素做更系统的探讨。
环境差异是造成底栖动物群落结构与功能存在差异的重要因素,并且食物来源不同,不同区域间底栖动物也会呈现出不同的营养水平。本研究发现,由于人工鱼礁的建设导致不同区域各营养级底栖动物数量发生变化,长礁龄区底栖动物群落中处于第三营养级的动物较对照区有所增加,且长礁龄区出现了蟹类等较高营养级的捕食者,表明底栖生态系统中高营养级动物的比例增加,这与徐勤增等[47]以及陈晨等[48]的研究结果一致。人工鱼礁建设能增加海区的空间异质性[49-50],同时为多种生物提供觅食及庇护场所,形成利于海洋生物栖息和繁育的适宜生境,使得底栖动物物种丰富度增加,进而促进食物网中不同生态位物种的同域共存[51-53]。
4 结论
本研究利用碳氮稳定同位素技术探究了莱州湾芙蓉岛人工鱼礁区大型底栖动物的营养结构以及营养关系,揭示了人工鱼礁建设时长对底栖动物营养结构的影响。随着人工鱼礁建设时间的增加,不同区域之间摄食类群的稳定同位素值略有变化,人工鱼礁建设能加强大型底栖动物对浮游植物碳源的利用,增加底栖动物群落的营养多样性以及丰度,促进群落中较高营养级动物的增加。研究表明,在芙蓉岛海域,长礁龄 (7~8 年)人工鱼礁产生了良好的生态效应,而短礁龄 (3 年)人工鱼礁生态效应尚不显著,说明在经历了礁体投放等人为扰动后,人工鱼礁的生态效应会随着建设时间的增加而逐步显现。本研究结果有助于了解人工鱼礁建设的生态效应以及底栖动物群落的营养结构,为底栖生境食物网物质循环和能量流动评价提供了基础资料。今后应探究不同季节间底栖动物稳定同位素的差异,同时增加对底栖高营养级捕食者的研究,另外也应利用其他食性分析方法对大型底栖无脊椎动物营养关系进行系统研究,以进一步明确和完善人工鱼礁建设对底栖生境的影响,以及对底栖动物群落结构和营养关系的生态效应。
(作者声明本文无实际或潜在的利益冲突)
猜你喜欢
海洋通报(2022年5期)2022-11-30
海洋通报(2020年5期)2021-01-14
江苏农业科学(2019年5期)2019-09-02
珠江水运(2018年21期)2018-12-20
广东农业科学(2017年5期)2017-08-29
电源技术(2015年7期)2015-08-22
中国海洋大学学报(自然科学版)(2015年8期)2015-03-18
应用海洋学学报(2014年1期)2014-11-22
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
