
蒙古栎生长节律及开花结实过程观测
2023-10-30 04:53陈建伟康晓梅蔡艺玮李树君程广有
北华大学学报(自然科学版) 2023年6期
陈建伟,康晓梅,蔡艺玮,李树君,程广有
(1.吉林市丰满区林业稽查大队,吉林 吉林 132013;2.吉林市丰满区林业和畜牧业管理局,吉林 吉林 132013;3.北华大学林学院,吉林 吉林 132013;4.吉林市丰满区国有林总场,吉林 吉林 132013)
蒙古栎(QuercusmongolicaFisch.ex Ledeb)又称柞树,为壳斗科栎属落叶乔木.作为中国北方的乡土植物,蒙古栎不仅拥有极高的经济及生态价值,还具备巨大的改良潜力[1-6].研究[7]表明,水分、光照、温度等环境因子是蒙古栎营养生长和生殖生长的重要影响因素.曾伟等[8]研究发现,水分胁迫仍是蒙古栎叶片光合参数的重要影响因素;胡明新等[9]利用大规模人工气候室研究发现,不同增温幅度和光周期对蒙古栎幼苗生长量的影响存在很大差异;韩金生[10]通过研究气候变化对蒙古栎径向发育及导管的影响发现,径向生长受温度和降水的共同限制,且对温度响应更为敏感.目前,已开展了蒙古栎种群结构[4]、经济价值[5]、生态功能[11]、繁育技术[12]等研究,而有关开花结实的研究却鲜有报道.本研究旨在探讨蒙古栎生长发育过程对气候的响应,观测、分析蒙古栎种子园内部分无性系叶片,新梢,雌、雄花及果实的生长发育情况,为培育蒙古栎速生林和提高蒙古栎种子园种子产量提供依据.
1 材料与方法
1.1 试验材料
试验材料来自吉林省临江市金山种子园.该园总面积约为56.3 hm2,其中,蒙古栎划分为2个小区,分别为蒙古栎Ⅰ区、蒙古栎Ⅱ区.蒙古栎Ⅰ区无性系主要来源于长白山,面积约2.5 hm2,区组内有20个无性系,共624株母树;蒙古栎Ⅱ区无性系主要来源于小兴安岭,面积约2 hm2,区组内有20个无性系,共约500株母树.以上两个小区的蒙古栎均采用嫁接繁殖,各无性系分株数均大于20个,2003年5月定植.
1.2 观测方法
营养生长:在蒙古栎Ⅰ区、Ⅱ区内分别选择3个无性系,每个无性系选取3株生长健壮且无明显病虫害的单株,每个单株选定树冠东南西北4个方向、上下两层共8个芽挂牌标记,于4月末至9月初定期观测芽萌动、叶片和新梢生长进程.生殖生长:自5月初起,定期观测选定植株的雌、雄花和果实发育过程[13],并记录同期气象因子变化情况.
1.3 数据分析
利用Excel 2019软件对观测数据进行整理统计,用SAS 9.4软件进行方差及相关性分析.
2 结果与分析
2.1 蒙古栎营养生长对气候的响应
蒙古栎不同时期无性系平均芽长、芽宽见表1.由表1可知:5月初,当日均温达到6 ℃左右时,蒙古栎枝条逐渐返青,圆锥形混合芽体积不断膨大,芽长日均增长量约为0.75 mm/d,芽宽日均增长量约为0.215 mm/d.直至5月7日左右,多数混合芽逐渐开裂,顶端紧抱的硬质芽鳞逐渐开裂、褪去,可见略带绒毛的柔嫩绿色尖端,剥开鳞片可见叶序轴和幼叶.不同无性系单株进入芽开裂期的时间存在差异,如Ⅰ-23号无性系在5月10日左右混合芽芽鳞开始脱落,而Ⅱ-18号无性系在5月7日左右混合芽已开裂至可见颜色鲜嫩的幼叶.
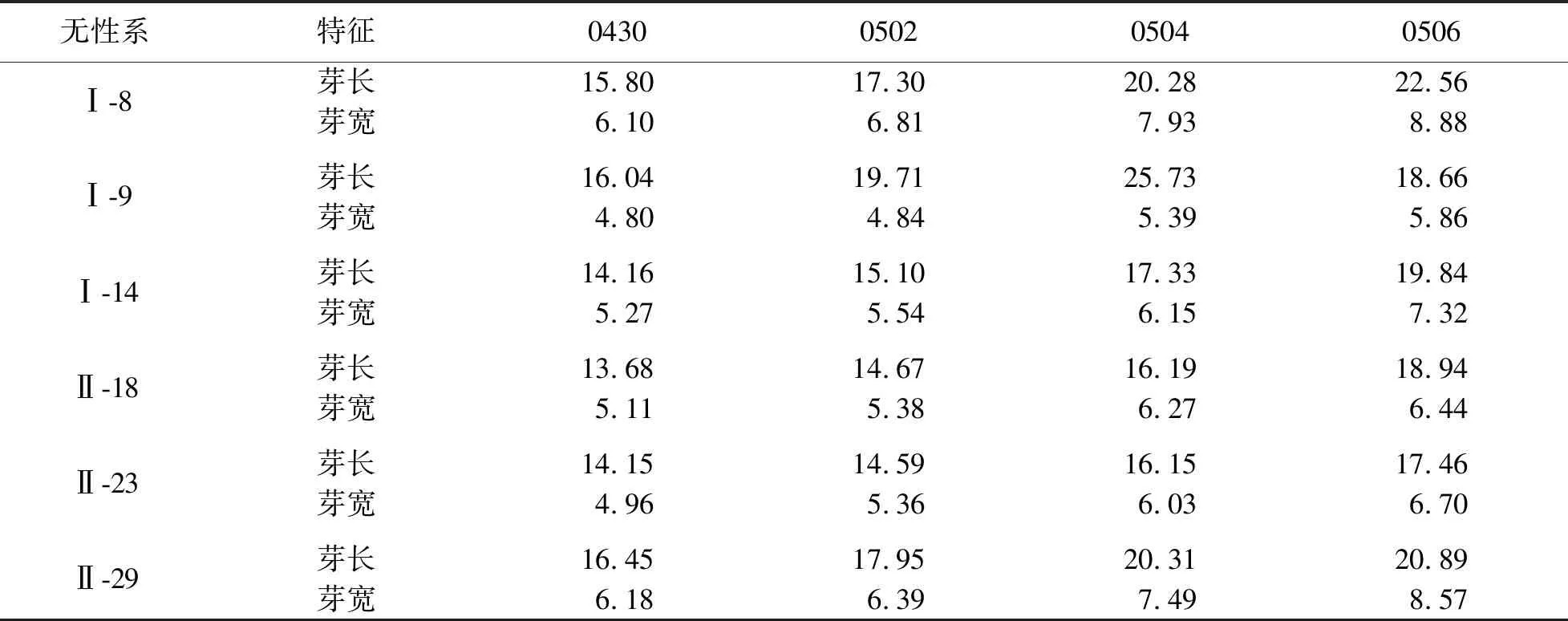
表1 蒙古栎不同时期无性系平均芽长、芽宽
蒙古栎叶片生长曲线见图1.由图1可见:叶片在最初的3周生长迅速,然后进入缓慢生长期.5月7日,当日均温达到7.5 ℃时,新叶开始生长.初期芽苞中伸出卷曲或按叶脉折叠着的小叶,叶长平均为19.38 mm,叶宽平均为5.49 mm,叶长生长速率为2.81 mm/d,叶宽生长速率为1.56 mm/d,此时叶片生长量占总生长量的18.31%(见表2).5月13日,当日均温达到10.5 ℃时,叶片半展,生长速度加快,此时叶片仍被松散的芽鳞包裹,且呈半聚拢状态,叶长为40.94 mm,叶宽为14.85 mm,叶长生长速率为4.43 mm/d,叶宽生长速率为5.17 mm/d,此时生长量占总生长量的20.93%.5月15日,第1批小叶从被芽鳞包裹的直立状态逐渐转至平展.5月16日,当日均温达到14 ℃时,半数以上枝条的混合芽芽鳞几乎全部脱落,叶面积随着气温的升高不断增大,进入展叶盛期,叶片颜色也逐渐由嫩绿色转变为深绿色,且不断增厚,此时叶长为54.24 mm,叶宽为30.35 mm,叶长生长速率为11.05 mm/d,叶宽生长速率为6.82 mm/d,生长速度达到全年最高水平.5月23日,当日均温达到18.5 ℃时,进入春色叶变色期,新叶从鲜嫩的绿色转为暗绿色,叶长平均达到 131.6 mm,叶宽达到78.06 mm,叶面积达到6.88 cm2,此时生长量占总生长量的79.07%.

图1 蒙古栎无性系叶长、叶宽
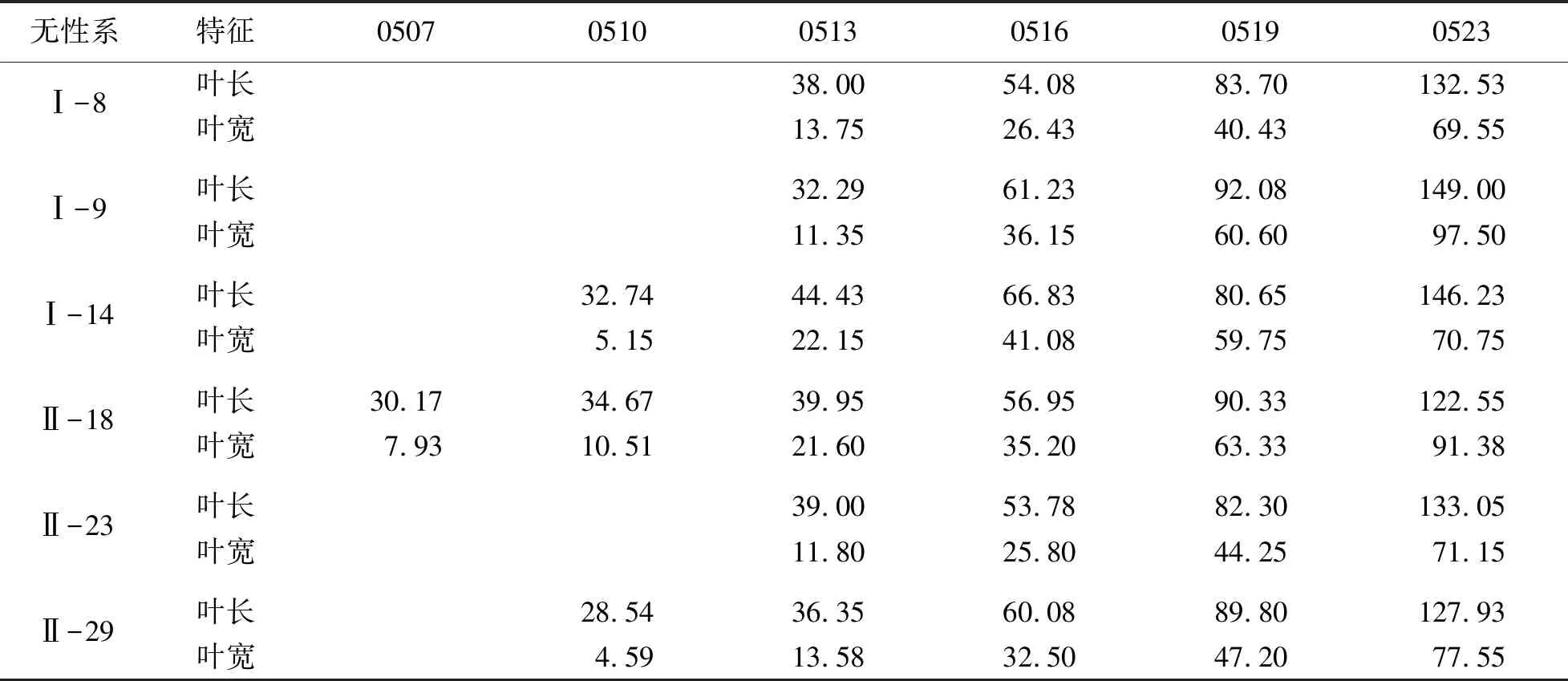
表2 蒙古栎无性系叶片生长性状特征
花期后,叶长、叶宽仍缓慢增加,逐渐加厚,革质化,叶色由深绿色变为墨绿色,淡黄色叶脉逐渐清晰、凸起.直至9月中旬,叶片边缘发黄、干枯、变脆,叶面逐渐布满锈色斑点,进入变色期.9月末至10月初,进入落叶期,多数叶片干枯、凋落,部分宿存.
5月中旬,蒙古栎新梢萌发,混合芽萌发后10 d左右,1 a生枝顶部新梢开始生长.新梢生长初期,纵向生长速度较快,生长速率为3.28 mm/d,5月18日进入花期后,生长速度明显减缓,生长量为1.61 mm/d,径向生长在短期内无明显变化.5月24日,蒙古栎新梢长度平均为32.18 mm,宽度为2.6 mm(图2、表3).

图2 蒙古栎无性系新梢长、新梢宽
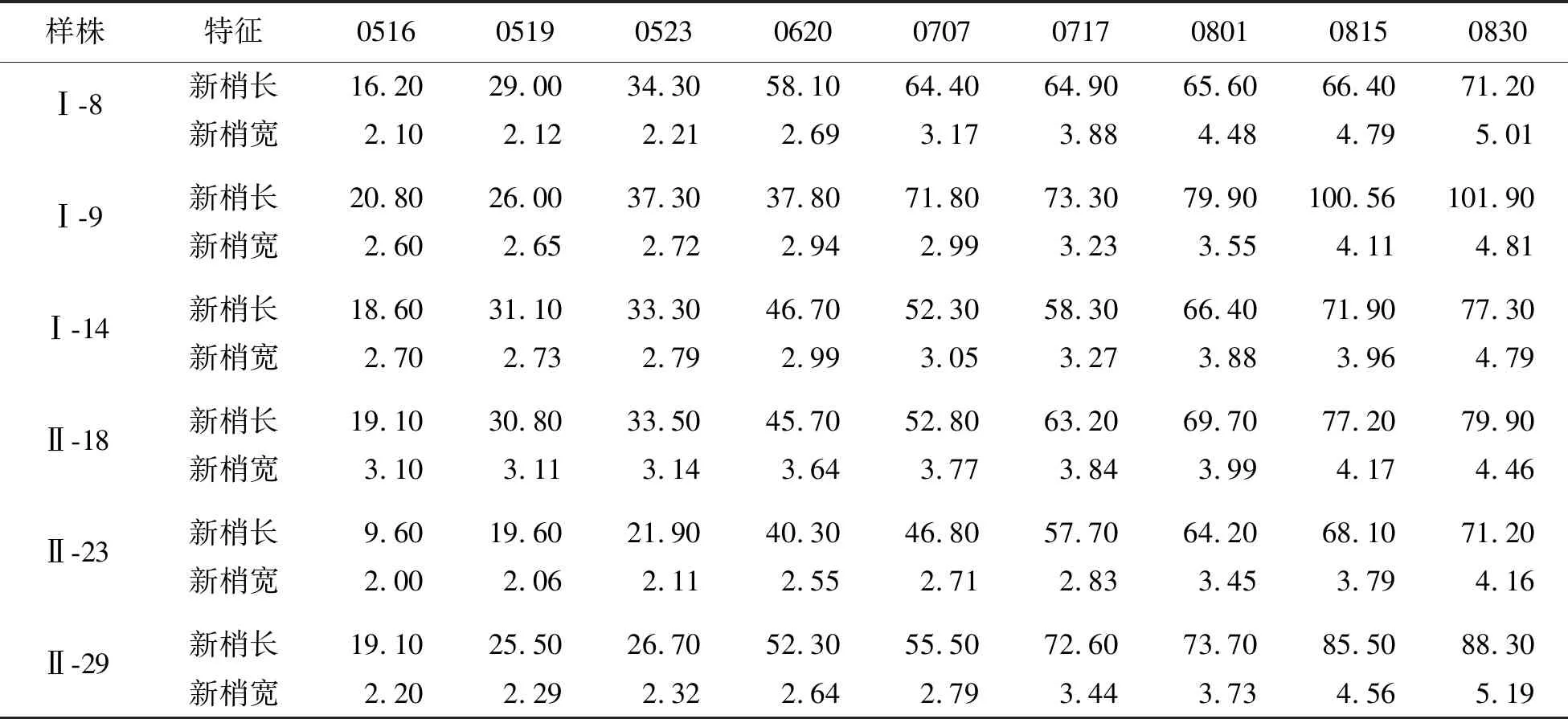
表3 蒙古栎无性系新梢生长性状数量特征
花期结束后,蒙古栎新梢继续生长,此时新梢长增长量为0.74 mm/d,新梢宽增长量为0.03 mm/d,8月末至9月初逐渐停止生长并达到最大.同时,柔软的新梢逐渐木质化,颜色由翠绿色逐渐变成深绿色,最后呈棕褐色,进入越冬休眠期.
2.2 蒙古栎营养生长与气象因子的相关性
蒙古栎营养生长与主要气候因子的相关性见表4.由表4可知:芽长与最高温度、最低温度、平均气温呈显著正相关,与降雨量呈极显著负相关;芽宽与最高温度、最低温度、平均气温呈显著正相关,与降雨量呈极显著负相关;叶长与最高温度、昼夜温差、风速、紫外线强度呈显著正相关,与最低温度和平均气温呈极显著正相关,与相对湿度呈显著负相关;叶宽与最高温度、昼夜温差、风速、紫外线强度呈显著正相关,与最低温度和平均气温呈极显著正相关,与相对湿度呈显著负相关;新梢长与最高温度、最低温度、平均气温呈极显著正相关,与紫外线强度呈显著正相关,与相对湿度呈极显著负相关;新梢宽与最高温度、平均气温呈极显著正相关,与最低温度呈显著正相关.由此可知,温度和光照是影响蒙古栎营养生长的最主要因素.

表4 蒙古栎营养生长与气象因子的相关性
2.3 蒙古栎生殖生长对气候的响应
2.3.1 雄花发育
蒙古栎花芽于上一年8月底完成分化,翌年春季继续发育.蒙古栎雄花序发育经过6个时期,包括混合芽萌动期、雄花序形成期、雄花序伸长期、花药形成期、雄花序散粉期、雄花序枯萎期.蒙古栎雄花序5月初开始生长,伸长生长较快(图3).5月13日,混合芽开裂后5 d,即可见由多个花粉囊组成的雄性柔荑花序,此时花序原基生长较快,花药紧凑,颜色呈嫩绿色,花序长为24.42 mm,宽度为3.32 mm,生长速率为9.61 mm/d,为雄花序形成期(表5).随着气温的升高,雄花序顶端微微发红,花轴迅速伸长,平均生长速率为7 mm/d,单日最大生长量为20.6 mm.5月18日雄花序长达到最大,平均为71.39 mm,最长可达117.5 mm,最短为40.1 mm,宽度为5.28 mm,并形成饱满的、较为分散的黄绿色花药.同时,部分花药开裂散粉,开裂顺序是从花序基部逐渐到顶部直至整个花序完全开放,花粉为金黄色.散粉后花药迅速萎缩干瘪,雄花序枯萎,逐渐变为灰褐色并凋落.

图3 蒙古栎无性系雄花序长、花序宽
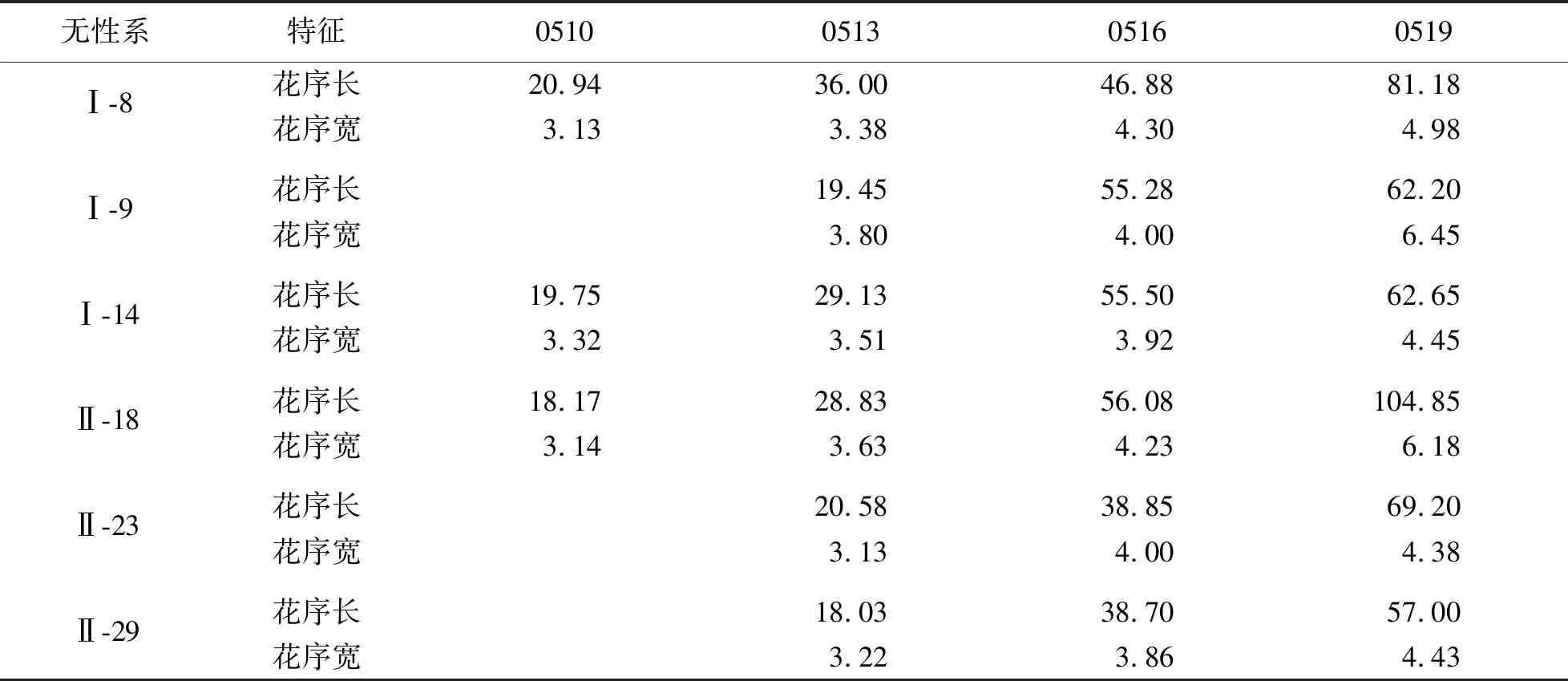
表5 蒙古栎无性系不同时期雄花序长、花序宽
2.3.2 雌花发育
蒙古栎雌花多生于新枝顶端,由花柱、总苞和胚珠3部分组成.依据形态变化,雌花自萌动起至开花分为5个时期,即萌动期、柱现期、柱裂期、柱干期、子房膨大期(表6).5月14日,混合芽萌发后7 d左右,可见新梢顶端出现3~5个微小的红色圆点状凸起,为雌花萌动期.5月16日,当日均温达到14 ℃左右时,雌花进入柱现期,多个雌花向上伸长,可见淡绿色细长花柱,并伴有淡黄色绒毛,此时柱头尚未开裂.5月18日,当日均温达16.5 ℃时,柱头开始大量开裂,期初开裂角度约为45°,随着雌花的进一步发育,柱头逐渐平展直至向外翻卷,开裂角度可达120°,表面较为湿润,呈嫩绿色,宽度约为2 mm,此时雌花直径为2 mm,最适宜接受花粉.授粉后,柱头转变为暗红褐色,表面光滑、干燥,进入柱干期,随后子房迅速膨大.

表6 蒙古栎雌、雄花发育过程
2.3.3 授 粉
当日平均温度达到16 ℃时,蒙古栎雄花序开始散粉,此时雌花柱头开裂,开裂角度多在45°,呈淡绿色花瓣状、湿润,易于接受花粉,进入开花始期.5月19日,进入开花盛期,50%以上植株雄花序散粉,此时雌花柱头开裂角度可达到120°,达到开裂最大角度,并向外翻卷.散粉时雄花序长为71.39 mm,宽为5.33 mm,达到可授粉状态的雌花直径为2 mm.5月23日,半数以上的蒙古栎雄花序散粉结束,干枯脱落,雌花柱头表面也变得光滑,并呈暗紫色,标志着蒙古栎进入开花末期.
2.3.4 果实发育
5月24日,授粉开始后7 d左右,雌花子房开始膨大,进入坐果期.蒙古栎幼果呈棕褐色,成熟时多为淡黄色.6月20日,叶腋可见黄绿色壳斗包裹的蒙古栎幼果,淡绿色花柱依然留存于壳斗中央,柱头干枯呈黑褐色,此时壳斗直径为5.52 mm,尚未见果实.随后壳斗不断增大,直至7月7日左右,壳斗外壁的小苞片更加凸起,呈三角状,壳斗略有增大,直径为8.05 mm,完全包裹住花柱,柱头呈硬革质三角状,分3~4片留存于壳斗底部中央,并微微向外翻卷.8月1日左右,壳斗外表面小苞片呈半球型瘤状凸起,底部向中间凹陷呈圆形,边缘呈粉红色,并可见壳斗内果实,此时壳斗直径为15.62 mm.至8月中旬,淡黄绿色壳斗内可见圆形光滑的深绿色果实,壳斗底部边缘变为灰白色,直径为20.02 mm,果实略凸起于壳斗底部.8月底,果实直径生长至最大,此时果实明显向外隆起,呈墨绿色,被绒毛并有光泽,果实底部向内凹陷,壳斗包裹种实的1/2,直径为26.99 mm.至9月中旬,壳斗逐渐干枯呈黄褐色,果实脱落,壳斗部分随果实脱落,部分宿存.
2.3.5 蒙古栎生殖生长与气象因子的相关性
蒙古栎生殖生长与气象因子的相关性见表7.由表7可知:雄花序长与最高温度、最低温度、平均气温呈极显著正相关,与昼夜温差、紫外线强度呈显著正相关,与相对湿度和降雨量呈显著负相关;雄花序宽与最高温度、最低温度、平均气温、昼夜温差呈极显著正相关,与紫外线强度呈显著正相关,与降雨量和相对湿度呈极显著负相关;雌花长与最高温度、最低温度、平均气温呈极显著正相关,与昼夜温差呈显著正相关,与相对湿度和降雨量呈显著负相关;果实长与最高温度、最低温度、平均气温呈极显著正相关,与昼夜温差呈显著正相关,与相对湿度和降雨量呈显著负相关;果实宽与最高温度、最低温度、平均气温呈极显著正相关,与相对湿度呈极显著负相关,与降雨量呈显著负相关.
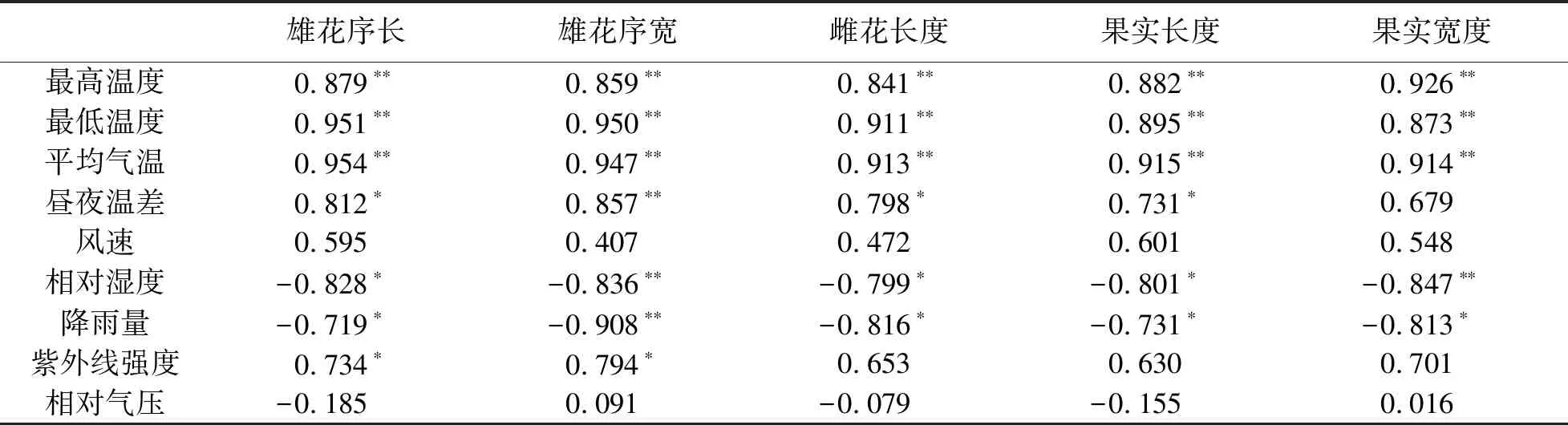
表7 蒙古栎生殖生长与气象因子的相关性
3 小结与讨论
3.1 小 结
1)通过营养生长观测发现,蒙古栎营养生长进程主要分为萌芽期、展叶期、新梢生长期和木质化等阶段.5月初,当日均温达到7.5 ℃左右时,蒙古栎混合芽鳞片开始脱落,并露出柔嫩绿色尖端,新叶进入生长阶段.直至5月中旬,小叶从被芽鳞包裹的直立状态逐渐转至平展,半数以上枝条的混合芽芽鳞几乎全部脱落,进入展叶期,叶片颜色也逐渐由嫩绿色转变为深绿色,且不断增厚.同时,新梢也进入快速生长阶段,翠绿柔嫩的新梢逐渐变得坚硬,直至8月末,新梢木质化呈棕褐色,此时叶片呈硬革质,颜色为深绿色,并逐渐从边缘开始枯萎、凋落.
2)蒙古栎花芽在7月开始形成,并于8月底至9月初完成分化.雄花序发育经过6个时期,包括混合芽萌动期、雄花序形成期、雄花序伸长期、花药形成期、雄花序散粉期、雄花序枯萎期.蒙古栎雌花多生于新枝顶端,自萌动起至开花,大致可分为萌动期、柱现期、柱裂期、柱干期、子房膨大期.5月18日左右雄花序开放散粉,雌花进入可授期,开花约持续7 d左右.授粉后的雌花子房开始膨大,进入坐果期.8月底,果实直径生长至最大,明显向外隆起,呈墨绿色,被绒毛,有光泽,果实底部向内凹陷,壳斗包裹种实的1/2,直径为26.99 mm.9月中旬,壳斗逐渐干枯,呈黄褐色,果实脱落,壳斗部分随果实脱落,部分宿存.
3.2 讨 论
植物生长发育不仅与自身遗传特性有关,还与外界环境条件(如温度、湿度、光照、土壤等)密切相关[14].温度升高与光周期增加的协同作用促进了蒙古栎春季物候提前,短光周期则明显抑制花芽的生长发育[15-16],长时间降雨易使花粉霉变,不利于花粉扩散[17].花期常伴随大风天气,频繁的空气流动有利于花粉扩散和雌花授粉,有助于花粉混合,缩短雌、雄花期,降低自交概率[18].在整个蒙古栎雌、雄花发育阶段,光照条件较好的晴天仅占整个阶段的20%,雨天占40%,日均空气湿度70%,特别是花期的强降水导致种子园内花粉受潮,极大影响了花粉传播和授粉.本研究发现,气温和降水是影响金山种子园蒙古栎营养和生殖生长的主要因素.提前收集和保存花粉[19]、人工辅助授粉可避免降雨以及长时间弱光照对授粉的不利影响,有效提高授粉成功率[20].
猜你喜欢
沙漠与绿洲气象(2022年5期)2022-02-03
科技风(2021年31期)2021-11-30
科技风(2021年32期)2021-11-30
新疆农业科学(2020年1期)2020-02-14
亚热带植物科学(2019年2期)2019-07-22
江苏农业学报(2018年5期)2018-09-10
农业知识(2018年18期)2018-05-28
江苏农业科学(2016年10期)2017-02-05
创新作文(3-4年级)(2016年9期)2016-12-22
科学种养(2016年2期)2016-02-23
