
脑神经炎症对小鼠运动疲劳的影响及其可能机制
2023-10-25 08:48曹奕炜
中国药理学与毒理学杂志 2023年9期
曹奕炜,宋 睿,吴 宁,李 锦
(1.南京中医药大学研究生院,江苏 南京 210023;2.军事科学院军事医学研究院毒物药物研究所,神经精神药理学北京市重点实验室,抗毒药物与毒理学国家重点实验室,北京 100850)
运动疲劳通常被认为是由于长时间高强度的体力运动引起的肌肉活动度下降与主观疲劳感,在体力劳动、体育竞技及军事领域会产生重大负面影响,深入研究运动疲劳产生的机制并发现延缓运动疲劳的措施具有重要意义。目前关于运动疲劳机制的研究主要集中在骨骼肌等外周组织和心脏等器官,但中枢神经系统在其中也发挥重要作用。支配肌肉运动的运动相关脑区发放神经冲动减少是引起肌群活动度下降的根源,最终可导致动作发力终止和运动能力降低,而脑能量代谢平衡是中枢神经系统持续发放神经冲动的重要保障。
运动过程中伴随外周骨骼肌细胞乳酸堆积并释放进入血液循环。乳酸一直被认为是代谢终产物,但有研究显示,外源性乳酸可维持脑切片细胞突触活动[1],神经元从低氧状态下恢复也需乳酸作为必要的能量底物[2],提示乳酸可作为神经元的能量底物。近年来研究发现,大脑在执行高水平认知活动时,神经元主要依靠乳酸(而非葡萄糖)作为能量底物产生ATP,维持突触活动[3];在高强度运动情况下,脑葡萄糖摄取随着运动强度的增加而降低,提示大脑可能利用乳酸补偿在高强度运动期间维持神经元活动所需的能量[4]。星形胶质细胞-神经元乳酸穿梭是神经元乳酸的主要来源,而单羧酸转运体1(monocarboxylate transporter-1,MCT-1)、MCT-2 和MCT-4 分别是星形胶质细胞释放乳酸和神经元摄取乳酸的通道,其表达水平与乳酸转运能力正相关[5]。运动过程中脑神经元是依靠葡萄糖还是乳酸供能一直存在争议。2017 年Matsui 等[6]报道,阻断神经元乳酸摄取可缩短大鼠跑台力竭运动时间,提示高强度耐力运动中大脑神经元也是以乳酸作为能量底物,而提高神经元乳酸供给可能延缓运动疲劳的发生。但MCT 各亚型在外周组织中亦广泛表达,因此很难实现对星形胶质细胞-神经元乳酸穿梭的选择性干预。
研究报道,耐力运动伴随着外周和脑炎症水平的升高[7-9],降低运动引起的外周血、肌肉及肝炎症水平可延缓运动疲劳发生[10-12],但脑神经炎症是否是运动疲劳发生的原因尚未见报道。本研究采用小鼠跑台实验和负重游泳实验2种力竭运动模型研究脑神经炎症状态与运动疲劳之间的因果关联,并从对星形胶质细胞-神经元乳酸穿梭影响的角度探讨其可能机制,为深入了解运动疲劳的中枢机制、寻找延缓疲劳的新的潜在干预措施提供实验依据。
1 材料与方法
1.1 动物、试剂和主要仪器
雄性C57BL/6J 小鼠,体重20~24 g,购自北京斯贝福生物技术有限公司,许可证号:SCXK(京)2019-0010。所有实验小鼠均常规饲养于含有玉米芯垫料的鼠笼中,室温恒定于20~24 ℃,湿度为20%~25%,12 h 昼夜交替,不限制饮水进食。实验动物的使用遵守军事医学研究院动物管理细则。
炎症诱导剂脂多糖(lipopolysaccharides,LPS)、小胶质细胞激活抑制剂米诺环素(minocycline)和二甲亚砜(dimethyl sulfoxide,DMSO),美国Sigma-Aldrich 公司;MCT-1 和MCT-4 共抑制剂AZD3965 及MCT-2 抑制剂4-CIN,美国Med Chem Express 公司;Trizol 试剂,美国Invitrogen公司;逆转录试剂盒和实时定量PCR(real-time quantitative PCR,RT-qPCR)试剂盒,德国QIAGEN 公司。前炎性细胞因子白细胞介素1β(interleukin-1β,IL-1β)、IL-6和肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)及MCT-1、MCT-2和MCT-4引物由上海生工生物工程有限公司合成,序列见表1。

Tab.1 Primer sequences for real-time quantitative PCR(RT-qPCR)
YLS-31A 大小鼠转棒测试仪,济南益延科技发展有限公司;SA101C动物跑台,江苏赛昂斯生物技术有限公司;梯度PCR 仪,德国PowerCycler 公司;LightCycler®96 实时荧光定量PCR 仪,瑞士罗氏公司。
1.2 药物配制、动物分组和处理
LPS和米诺环素溶于生理盐水,AZD3965溶于含10%DMSO生理盐水,4-CIN溶于含15%DMSO生理盐水,均采用侧脑室注射方式(intracerebroventricular injection,icv)给药,给药体积为每只小鼠5 μL。小鼠icv 给予炎症诱导剂LPS 诱导脑神经炎症状态,icv给予小胶质细胞激活抑制剂米诺环素抑制脑神经炎症状态;icv 给予MCT-1 和MCT-4 共抑制剂AZD3965 阻断星形胶质细胞乳酸释放,icv给予MCT-2抑制剂4-CIN阻断神经元乳酸摄取。
①药物对小鼠运动协调性的影响实验分组:分为溶剂对照和LPS 组(每只2.5 μg)、米诺环素组(每只12 μg)、AZD3965 组(每只50 nmol)、4-CIN组(每只40 nmol),每组12只。②米诺环素对小鼠运动疲劳的影响实验分组:小鼠跑台实验分为溶剂对照和米诺环素3 个剂量组(每只3,6 和12 μg),每组12只;小鼠负重游泳实验分为溶剂对照和米诺环素组(每只12 μg),每组12 只。③LPS 对小鼠运动疲劳及大脑运动皮质前炎性细胞因子和MCTmRNA 表达的影响及米诺环素的阻断作用实验分组:分为溶剂对照、LPS(每只2.5 μg)和LPS+米诺环素组(每只2.5 μg+12 μg),每组12 只。④AZD3965 和4-CIN 对小鼠运动疲劳的影响实验分组:小鼠跑台实验分为溶剂对照和AZD3965 3个剂量组(每只12.5,25 和50 nmol)或4-CIN 3 个剂量组(每只10,20 和40 nmol);小鼠负重游泳实验分为溶剂对照和AZD3965 组(每只50 nmol)或4-CIN组(每只40 nmol),每组12只。
参考文献[6,13-14],LPS 给药后12 h 和米诺环素给药后30 min 进行动物行为学测试或处死取大脑运动皮质;LPS 联合米诺环素给药为LPS 给予后12 h给予米诺环素,30 min后进行动物行为学测试或处死取大脑运动皮质;AZD3965 和4-CIN 给药后30 min进行动物行为学测试。
1.3 小鼠转棒实验
小鼠正式测试前预适应转棒3 d,每天1次。预适应时转棒起始转速为1 r·min-1,最终转速为10 r·min-1,转棒持续时间300 s。最终选择在第3 天适应时在300 s 内从转棒上落下次数<3 次的小鼠进行正式测试。正式测试时转棒起始转速为1 r·min-1,在15 s 内加速至25 r·min-1,测试截止时间为300 s。通过测试小鼠在转棒上保持平衡的时间(在棒时间),对小鼠的运动协调性进行评估。
1.4 小鼠负重游泳实验
使用高50 cm、直径15 cm 的玻璃水缸为容器,每次实验加入水,深度为25 cm;使用调节热水比例的方式调整水温为23~25 ℃。小鼠正式测试前进行预适应。预适应时不在尾部捆绑铅皮卷,自由游泳3~5 min。正式测试时小鼠尾根部捆绑5%体重的铅皮卷,完全打湿毛发后进行测试。小鼠保持平衡漂浮不动时采用玻璃棒轻推驱赶。当小鼠没于水面以下>8 s、无法继续维持游泳运动时判定小鼠力竭疲劳,同时记录小鼠负重游泳时间。
1.5 小鼠跑台实验
参考文献[15]的实验方法进行3 d 跑台训练预适应。第1 次训练前将小鼠置跑道内自由探索1~3 min 熟悉环境。然后采用逐级递增的速度进行跑台训练,每天1次。每次训练结束后使用抹布清理跑台通道,并喷涂乙醇、使用抹布擦拭去除气味。训练方案如下。
第1 天:2 m·min-1(1 min)→4 m·min-1(1 min)→6 m·min-1(1 min)→8 m·min-1(1 min)→9 m·min-1(2 min)→10 m·min-1(4 min)。
第2天:4 m·min-1(1 min)→6 m·min-1(1 min)→8 m·min-1(1 min)→10 m·min-1(2 min)→11 m·min-1(3 min)→12 m·min-1(5 min)。
第3天:8 m·min-1(1 min)→10 m·min-1(1 min)→11 m·min-1(2 min)→12 m·min-1(3 min)→13 m·min-1(5 min)→14 m·min-1(8 min)。
正式测试时跑台速度设置为:14 m·min-1(2 min)→16 m · min-1(3 min)→18 m · min-1(25 min)→20 m·min-1(15 min)→22 m·min-1(15 min)→24 m·min-1(15 min)→26 m·min-1(≥60 min)。跑台速度程序最高速度为26 m·min-1。
当小鼠无法在跑台通道上继续维持跑步,长时间处在跑道末端或伏地式前进,且跑台电击与人为驱赶也无法使其跑至跑道前端时判定为小鼠力竭疲劳,记录跑步持续时间和距离。为确保疲劳判断的准确性,采用盲法判定小鼠疲劳状态。
1.6 RT-qPCR 测定小鼠大脑运动皮质前炎性细胞因子和MCT mRNA水平
给药结束后处死小鼠取脑,于冰上快速分离运动皮质脑区组织,保存于盛有预冷Trizol 试剂的1.5 mL EP管内,于-80 ℃冰箱保存。
参考RNA 提取试剂盒的流程提取组织总RNA,用蛋白核酸分析仪对RNA 的浓度和质量进行测定。A260nm/A280nm值在1.9~2.1 范围内,表明RNA 样品合格,用于进行后续RT-qPCR 实验。逆转录和RT-qPCR 实验分别按说明书步骤进行,测定IL-1β,IL-6和TNF-α及MCT-1,MCT-2和MCT-4mRNA 表达。以β 肌动蛋白为内参基因,用2-ΔΔCt法计算待测基因mRNA相对表达水平。
1.7 统计学分析
对满足正态性与方差齐性的实验结果数据用±s表示,否则采用中位数±四分位间距(median±interquartile)表示。用SPSS 24.0软件进行统计学分析。首先采用Levene 检验和Shapiro-Wilk 检验对数据的方差齐性和正态性进行检验;若数据满足正态性与方差齐性,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析结合Bonferroni检验;若数据不满足上述条件则采用Mann-Whitney非参数检验(两组间比较)或Kruskal-Wallis 非参数检验(多组间比较)进行分析。P<0.05 表示差异具有统计学意义。
2 结果
2.1 LPS、米诺环素、AZD3965 和4-ClN 对小鼠运动协调性的影响
与溶剂对照组比较,LPS(2.5 μg)、米诺环素(12 μg)、AZD3965(50 nmol)和4-CIN(40 nmol)对小鼠转棒实验中在棒时间均无明显影响(图1),提示上述药物处理不影响小鼠的运动协调性,可排除对小鼠跑台实验和负重游泳实验的干扰。
Fig.1 Effects of lipopolysaccharides(LPS)(A),minocycline(B),AZD3965(C)and 4-ClN(D)administration on motor coordination of mice. Mice were intracerebroventricularly injected(icv)with LPS(2.5 μg per mouse)12 h before the rotate bar test,while minocycline(12 μg per mouse),AZD3965(50 nmol per mouse)and 4-CIN(40 nmol per mouse)were injected 30 min before the rotate bar test.The injection volume was 5 μL per mouse.Median±interquartile,n=12.
2.2 米诺环素对小鼠运动疲劳的影响
小鼠跑台实验结果表明,与溶剂对照组比较,米诺环素(12 μg)组小鼠运动持续时间和运动距离显著增加(P<0.01),3 和6 μg 组两指标均无明显变化(图2A1和A2)。小鼠负重游泳实验结果表明,与溶剂对照组比较,米诺环素(12 μg)组小鼠游泳持续时间显著增加(P<0.01,图2B)。
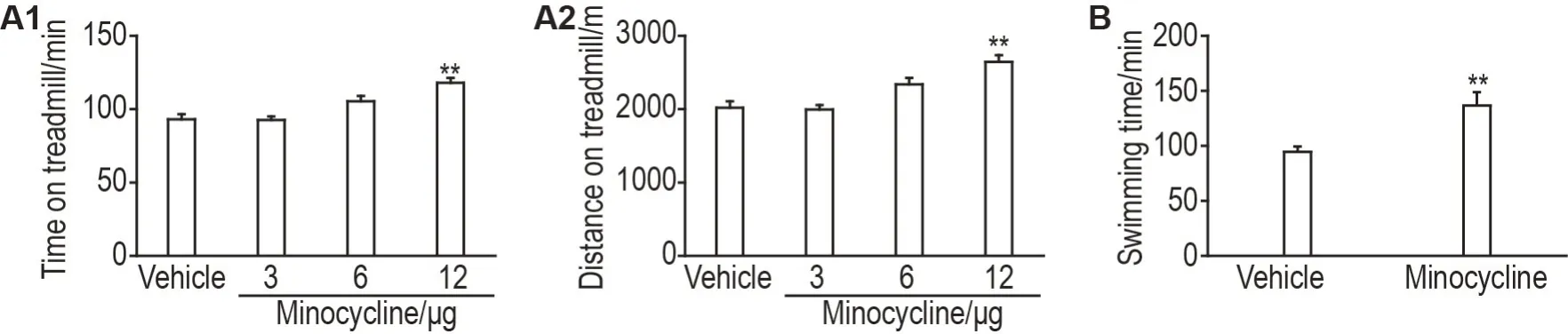
Fig.2 Effects of minocycline on exercise capacity of mice.Mice were icv with minocycline(3,6 and 12 μg per mouse)30 min before the treadmill test.Mice were icv with minocycline(12 μg per mouse)30 min before the load swimming test.The injection volume was 5 μL per mouse. A1 and A2:running duration and distance covered by mice on the treadmill,respectively;B:swimming time of mice.±s,n=12.**P<0.01,compared with vehicle group.
2.3 LPS 对小鼠运动疲劳的影响及米诺环素的阻断作用
在小鼠跑台实验中,与溶剂对照组比较,LPS(2.5 μg)组小鼠运动持续时间和运动距离显著降低(P<0.01);与LPS 组比较,LPS+米诺环素(2.5 μg+12 μg)组小鼠运动持续时间和运动距离显著增加(P<0.01)(图3A1 和A2)。在小鼠负重游泳实验中,与溶剂对照组比较,LPS 组小鼠游泳持续时间显著减少(P<0.01);与LPS 组比较,LPS+米诺环素组小鼠游泳持续时间显著增加(P<0.05)(图3B)。
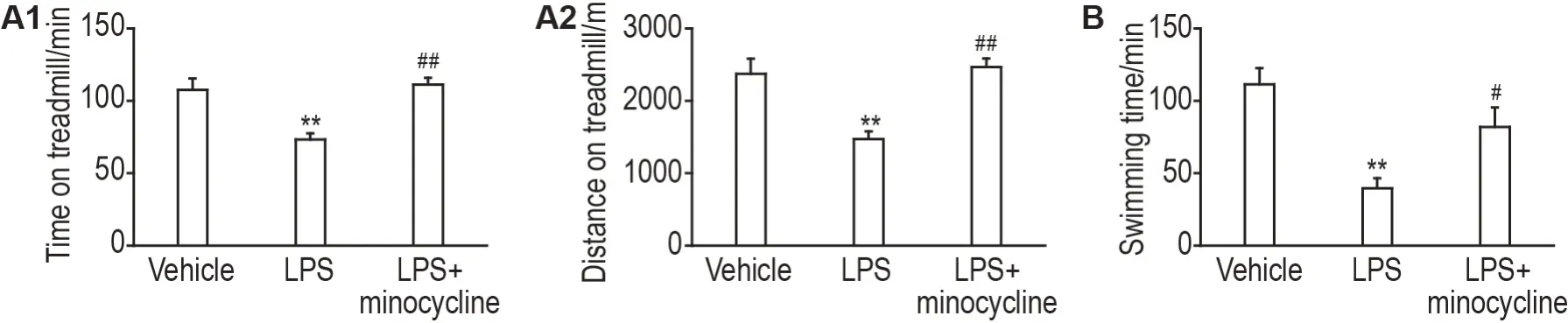
Fig.3 Effects of minocycline on exercise capacity of mice. Mice were icv with LPS(2.5 μg per mouse)12 h,minocycline(12 μg per mouse)30 min or icv minocycline(12 μg per mouse)30 min after icv with LPS(2.5 μg per mouse)12 h before the treadmill test and the load swimming test. The injection volume was 5 μL per mouse. A1 and A2:running duration and distance covered by mice on the treadmill,respectively;B:swimming time of mice.±s,n=10-12. **P<0.01,compared with vehicle group;#P<0.05,##P<0.01,compared with LPS group.
2.4 AZD3965和4-ClN对小鼠运动疲劳的影响
小鼠跑台实验结果表明,与溶剂对照组比较,AZD3965(50 nmol)组小鼠运动持续时间和运动距离显著降低(P<0.01),12.5 和25 nmol 组无明显变化(图4A1 和A2);4-CIN(40 nmol)组小鼠运动持续时间和运动距离显著降低(P<0.05),10 和20 nmol 组无明显变化(图4B1 和B2)。小鼠负重游泳实验结果表明,与溶剂对照组比较,AZD3965(50 nmol)组小鼠游泳持续时间显著降低(P<0.01,图4C);4-CIN(40 nmol)组小鼠游泳持续时间显著降低(P<0.05,图4D)。
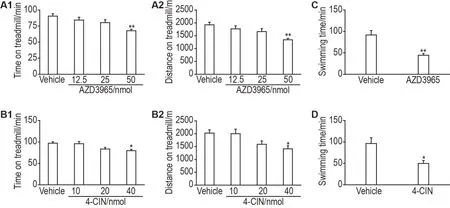
Fig.4 Effects of AZD3965 and 4-ClN on exercise capacity of mice. Mice were icv with AZD3965(12.5,25 and 50 nmol per mouse)or 4-CIN(10,20 and 40 nmol per mouse)30 min before the treadmill test. Mice were icv with AZD3965(50 nmol per mouse)or 4-CIN(40 nmol per mouse)30 min before the load swimming test. The injection volume was 5 μL per mouse. A1 and A2:running duration and distance covered by mice on the treadmill,respectively,after icv AZD3965;B1 and B2:running duration and distance covered by mice on the treadmill,respectively,after icv 4-CIN;C:swimming time of mice after icv AZD3965;D:swimming time of mice after icv 4-CIN.±s,n=12.*P<0.05,**P<0.01,compared with vehicle group.
2.5 LPS 对小鼠大脑运动皮质前炎性细胞因子mRNA表达水平的影响及米诺环素的逆转作用
与溶剂对照组比较,LPS(2.5 μg)组小鼠大脑运动皮质IL-1β,IL-6和TNF-αmRNA 水平均显著升高(P<0.01),提示icv 给予LPS 可诱导小鼠处于高水平的脑神经炎症状态。与LPS组比较,LPS+米诺环素(2.5 μg+12 μg)组小鼠大脑运动皮质IL-1β和TNF-αmRNA 水平显著下降(P<0.05),IL-6mRNA水平无明显变化(图5),提示米诺环素可降低LPS诱导的神经炎症水平升高。
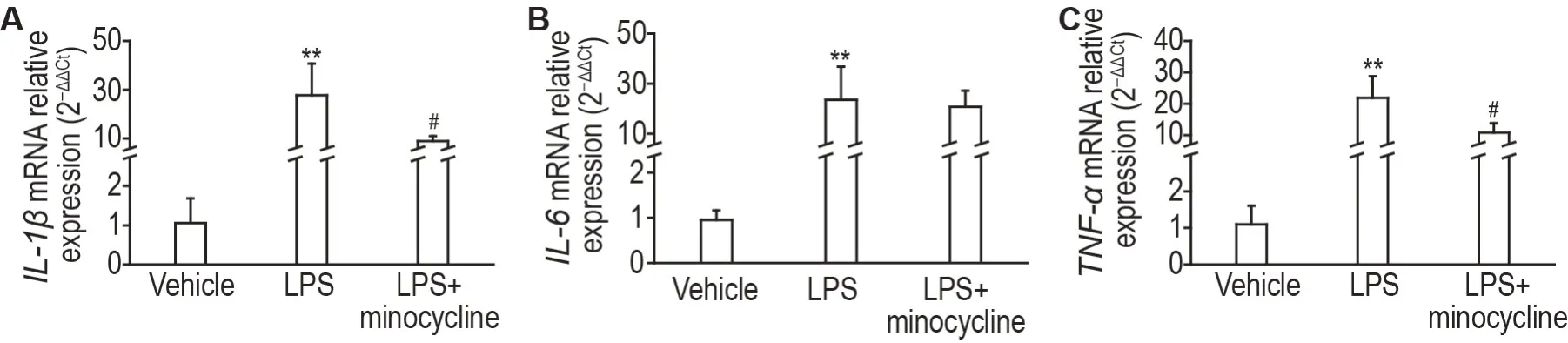
Fig.5 Effect of minocycline on IL-1β(A),IL-6(B)and TNF-α(C)mRNA expressions of motor cortex of mice.Mice were icv with LPS(2.5 μg per mouse)12 h or minocycline(12 μg per mouse)30 min before the mice were sacrificed. The injection volume was 5 μL per mouse.±s,n=10-12.**P<0.01,compared with vehicle group;#P<0.05,compared with LPS group.
2.6 LPS 对小鼠大脑运动皮质MCT mRNA 表达水平的影响及米诺环素的逆转作用
与溶剂对照组比较,LPS(2.5 μg)组小鼠大脑运动皮质MCT-1,MCT-2和MCT-4mRNA 水平均显著降低(P<0.05);与LPS 组比较,LPS+米诺环素(2.5 μg+12 μg)组小鼠大脑运动皮质MCT-1,MCT-2和MCT-4mRNA 水平均显著升高(P<0.01),且接近溶剂对照组水平(图6)。
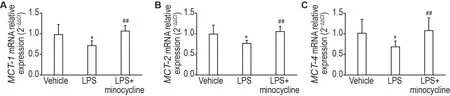
Fig.6 Effect of minocycline on MCT-1(A),MCT-2(B)and MCT-4(C)mRNA expressions of motor cortex of mice.See Fig.5 for the mouse treatment.±s,n=10-12.*P<0.05,compared with vehicle group;##P<0.01,compared with LPS group.
3 讨论
疲劳的定义为“机体生理过程不能将其机能持续在特定水平上或器官不能维持其预定的运动强度”。疲劳发生的学说主要包括能量耗竭学说、代谢产物堆积学说、内环境稳态失调学说、自由基影响理论等,其中“能量耗竭学说”占主导地位。现有运动疲劳机制的研究主要集中在外周组织器官。本研究以中枢神经系统为切入点,进一步探究运动疲劳产生的中枢机制。脑能量代谢具有与外周组织器官显著不同的特点。尽管依然存在一些争议,但越来越多的研究倾向于认为在大脑进行高强度认知活动时神经元以乳酸作为主要的能量底物。神经元很少储存能量底物,星形胶质细胞糖酵解产生的乳酸经由星形胶质细胞-神经元乳酸穿梭进入神经元中,通过三羧酸循环及线粒体氧化磷酸化生成ATP,维持神经元的兴奋性及突触活动[16]。但运动过程中脑神经元是否依赖于乳酸作为能量底物、维持运动相关脑区神经元兴奋性及机体运动状态,尚存在争议。本研究采用2种经典的啮齿类动物耐力运动实验(小鼠跑台实验和小鼠负重游泳实验)发现,阻断脑星形胶质细胞释放乳酸(icv 给予MCT-1 和MCT-4 共抑制剂AZD3965)或阻断神经元摄取乳酸(icv 给予MCT-2 抑制剂4-CIN)均可显著降低小鼠的耐力运动持续时间和运动距离,提示由星形胶质细胞-神经元乳酸穿梭供给的乳酸是小鼠维持耐力运动所需的神经元能量底物,与Matsui等[6]研究结果一致。
研究发现,大鼠跑台运动至力竭时海马脑区IL-1β 和TNF-α 水平显著增加[17-18],大鼠力竭游泳显著增加杏仁核IL-1β 免疫阳性细胞数量[19],提示耐力运动至力竭伴随脑神经炎症水平的升高,但升高的神经炎症是否是导致力竭疲劳的原因尚不明确。本研究采用icv 给予LPS 诱导小鼠处于高水平的脑神经炎症状态(运动皮质前炎性细胞因子IL-1β,IL-6和TNF-α水平显著升高)后,小鼠耐力运动持续时间和运动距离明显缩短,而icv 给予米诺环素降低脑炎症水平则延长小鼠耐力运动持续时间和运动距离,为小鼠大脑神经炎症状态与运动疲劳的因果关系提供了实验证据。另外,本课题组尚未发表的研究显示,小鼠耐力运动至力竭进程中运动相关脑区的神经炎症水平逐渐升高。结合本研究结果推测,降低脑神经炎症水平可能是延缓运动疲劳发生的有效措施。
相关研究报道,阿尔茨海默病导致脑内呈现高水平的神经炎症,同时星形胶质细胞葡萄糖利用率降低、MCT-1 下调及乳酸释放减少[20],使用趋化因子受体4 可逆拮抗剂减轻神经炎症可上调MCT-1并显著改善认知与记忆功能[21],提示疾病状态下的神经炎症可能通过影响MCT 表达及星形胶质细胞-神经元乳酸穿梭过程而导致神经元能量底物乳酸供给障碍。本研究结果表明,LPS诱导小鼠大脑运动皮质前炎性细胞因子IL-1β,IL-6和TNF-αmRNA 表达水平升高,同时降低MCT-1,MCT-2和MCT-4mRNA 表达水平;而给予米诺环素可降低运动皮质IL-1β和TNF-αmRNA表达的同时,上述MCTmRNA 表达也恢复至溶剂对照组水平,表明高水平的神经炎症状态可导致星形胶质细胞-神经元乳酸穿梭中乳酸转运通道——MCT 表达下调。另外,本课题组尚未发表的实验研究显示,前炎性细胞因子及其下游信号通路导致MCT 蛋白表达下降;而icv前炎性细胞因子受体的中和抗体亦可延缓小鼠运动疲劳的发生。因此推测,脑神经炎症驱动运动疲劳发生的机制可能是通过下调MCT 表达而影响星形胶质细胞-神经元乳酸穿梭实现的。
综上所述,本研究结果表明,星形胶质细胞-神经元乳酸穿梭是小鼠维持耐力运动的神经元能量底物供给途径,且脑高水平的神经炎症状态是驱动运动疲劳发生的重要原因,其机制可能与下调星形胶质细胞-神经元乳酸穿梭中的关键分子MCT表达有关。
猜你喜欢
传染病信息(2022年2期)2022-07-15
当代水产(2021年3期)2021-07-20
数学物理学报(2019年3期)2019-07-23
当代医药论丛(2017年22期)2017-04-12
中国卫生标准管理(2015年8期)2016-01-15
淮海医药(2015年2期)2016-01-12
现代检验医学杂志(2015年1期)2015-02-06
中国药业(2014年17期)2014-05-26
东北师大学报(自然科学版)(2014年1期)2014-02-27
东北石油大学学报(2013年6期)2013-03-11
