
电子加速器源射线对小麦根尖染色体及生理特性的影响
2023-10-23 08:14管翊君吕美澄白俊青周春菊李吕金印
核农学报 2023年11期
管翊君 王 浩 吕美澄 白俊青 唐 燕 周春菊李 奎, 吕金印,
(1西北农林科技大学生命科学学院,陕西 杨凌 712100;2杨凌核盛辐照技术有限公司,陕西 杨凌 712100)
小麦是我国第二大谷类作物,随着全球气候和环境变化,提高小麦产量对我国保障粮食安全及社会稳定有重要意义[1]。在小麦新品种培育过程中,传统育种方式突变率低,而人工诱变将植物的突变率提高了10~1 000 倍[2],其中辐射育种周期短、突变率较高、扩大突变谱同时能够突破原有基因库的限制,产生更多样的突变表型[3-4]。因此,利用辐射诱变育种创制作物新品种的优势明显[5],育种进程加快,开启了作物改良的新纪元[6]。
辐射诱变育种是指利用物理辐射源处理植物材料,诱导植物的遗传物质发生改变,直接或间接地育成可供生产利用的新品种。辐射诱变包括电离辐射(γ、β、X 射线等)和非电离辐射(激光、紫外线等)。目前,应用较广的是γ射线[7],王泽港等[8]采用γ射线辐射处理小麦种子,结果表明剂量越大,苗高越低、生长受抑制的程度越明显;陈金跃等[9]采用137Cs-γ 处理玉米种子,其苗高和根系的生长受到抑制,并且发现玉米的半致死剂量小于450 Gy;李春牛等[10]采用60Co-γ 处理茉莉花果实,发现高剂量辐射可使茉莉花叶片中抗氧化酶活性下降。除此之外,紫外线、激光和混合离子场等诱变源的研究也有报导[11-13]。刘瑞媛等[14]研究表明,高能重离子束能够沿着粒子路径紧密地形成在局部区域,能量传递线密度较高,对目标生物的DNA 和蛋白质等分子造成严重破坏;谭瑗瑗等[15]利用高能碳离子束辐射水稻发现并获得粳稻两系不育系江79S;王雪等[16]采用碳离子束辐射处理大豆,结果表明大豆M1代叶绿素及丙二醛含量低于M2代;王鹏[17]采用紫外线处理小麦种子,发现处理后的小麦生长发育迟缓,具有“翘根”现象;崔玲艳等[18]利用紫外线辐射处理马铃薯,发现低剂量的辐射对马铃薯的生长不会产生危害,适当增加辐射剂量能够提高植株抗氧化酶活性及次生代谢产物。
电子加速器的射线主要包括电子束、电子照射在传输链及货物后发生韧致辐射产生的X 射线和光中子[19]。电子加速器所产生的射线作为辐射源用于诱变育种,可通过调节与射线源距离控制剂量率,调整辐射时间控制辐射总剂量,射线利用率更高,成本更低,安全性更强,不产生放射性废物[20-22]。杨震等[23]采用电子束辐射处理水稻,并与传统60Co-γ 辐射进行比较,结果表明两种辐射方法对水稻的影响接近,而电子束辐射易于控制,可重复性更强。
目前,电子束辐射在植物育种中的研究相对较少,对不同辐射剂量处理下小麦的生长发育过程的影响机理以及辐照适宜剂量仍不明确。鉴于此,本研究采用电子加速器所产生的源射线辐射处理小麦种子,分析不同剂量辐射处理下水培小麦幼苗叶片抗氧化酶活性等生理与生长特性,观察根尖染色体形态变化,筛选合适的辐射剂量,以期为阐明电子束辐射诱变机理、拓宽小麦育种途径及农作物物理诱变育种提供参考。
1 材料与方法
1.1 试验材料
1.1.1 辐射处理 试验选用小麦品种为普冰151,由西北农林科技大学农学院张正茂教授提供。挑选籽粒饱满、大小均匀的的小麦种子分为3 组,每组500 g,对照组未辐射,处理组使用DZ 型10 MeV 电子直线加速器(中广核辐照技术有限公司)进行辐射处理,辐射剂量分别为0、200、400 Gy,剂量率为1 Gy·min-1。
1.1.2 水培小麦幼苗 辐射处理后的小麦种子在75%酒精溶液中消毒10 min 后用蒸馏水冲洗5~8 次,摆放在铺有滤纸的培养皿中,用纱布覆盖,使培养皿保持湿润状态,置于恒温培养箱内萌发。萌发3 d 后,将萌发的种子转移至1/2 Hoagland 营养液中进行光照培养,每隔2 d更换一次营养液。培养箱条件:光照强度为80 μmol·m-2·s-1,培养温度为22 ℃,光照周期为光/暗:16 h/8 h。待小麦长至两叶一心期时取样用于试验。
1.2 试验方法
1.2.1 幼苗生长特性的测定 将消毒后的种子转移到铺有滤纸的培养皿上培养,每日观察种子的发芽情况,7 d后按照公式计算种子的发芽率和发芽势。
发芽率=第7 天种子正常发芽数/供试种子数×100%
发芽势=每一天正常发芽的种子数/供试种子数×100%。
萌发3 d后的种子移至培养液中,水培10 d后利用直尺测量法测量小麦的株高和根长;利用天平称量法测量鲜重;利用烘箱将幼苗烘至恒重后使用天平对地上和地下生物量进行称重,其中地上生物量为小麦叶片,地下生物量包括种子内未被完全降解、利用的储藏物质及小麦根部。每个指标随机选取10 株幼苗用于统计分析。
1.2.2 生理指标的测定 叶绿素含量(chlorophyl,Chl)测定采用80%丙酮提取比色法。称取小麦新鲜叶片0.2 g,用剪刀剪成细丝状后放入装有15 mL 80%的丙酮的具塞试管中,黑暗处过夜提取至叶片变为白色,随后过滤。以80%丙酮为空白对照,分别在663 和645 nm 下测定过滤后得到的叶绿素提取液吸光值。每个处理设3个生物学重复。
丙二醛(malondialdehyde,MDA)含量测定采用硫代巴比妥酸比色法。称取小麦新鲜叶片0.3 g,在预冷过的研钵中加入适量石英砂和2 mL 0.05 mol·L-1pH值7.8 的磷酸缓冲液,研磨成匀浆,转移至10 mL 离心管中,加入5 mL 0.5% 硫代巴比妥酸溶液摇匀。沸水浴10 min 后迅速冷却,3 000 r·min-1离心15 min,以0.5%硫代巴比妥酸溶液为空白,测定上清液在532、600、450 nm处的吸光值。每个处理设3个生物学重复。
可溶性蛋白含量测定采用考马斯亮蓝G-250 法。称取小麦新鲜叶片0.2 g,以2 mL 0.05 mol·L-1pH 值7.8 的磷酸缓冲液为提取液在研钵中研磨至匀浆,5 000 r·min-1离心10 min。取0.9 mL 上清液加入考马斯亮蓝G-250 溶液2.5 mL,充分混匀后静置2 min,在595 nm 下测量吸光值,空白对照为加入考马斯亮蓝G-250的磷酸缓冲液。每个处理设3个生物学重复。
超氧阴离子测定采用NBT 染色法。取5片新鲜小麦叶片,用蒸馏水冲洗干净,擦干放入装有氮蓝四唑(nitro-blue tetrazolium,NBT)染液的具塞试管中,22 ℃条件下避光染色9 h,至阳性部位出现深蓝色,其他部位为淡蓝色或不变色。将染色叶片浸入95%乙醇制成的脱色液中,95 ℃中将未染色部分脱色至白色,蒸馏水冲洗干净后拍照。
抗氧化酶活性测定:称取小麦新鲜叶片0.3 g,在预冷过的研钵中加入适量石英砂和2 mL 0.05 mol·L-1pH 值7.8 的磷酸缓冲液,研磨成匀浆,转移至离心管中,12 000 r·min-1离心10 min,上清即为酶液。采用愈创木酚氧化法测定过氧化物酶(peroxidase,POD)活性,取0.2 mL 上清液加入2 mL 磷酸缓冲液、0.5 mL 愈创木酚溶液、0.2 mL 30%的H2O2溶液于试管中,每隔1 min 测定470 nm 下吸光值,共测3 次;采用氯化硝基四氮唑蓝(nitrotetrazolium blue chloride,NBT)光化还原法测定超氧化物歧化酶(superoxide dismutase,SOD)活性,取0.2 mL 上清液加入2.7 mL 甲硫氨酸溶液、0.6 mL 乙二胺四乙酸二钠(EDTA-Na2)溶液、0.1 mL核黄素溶液于试管中,在5 000 lux 的光照培养箱中光照20 min 后,遮光保存,测量560 nm 下吸光值;采用紫外吸收法测定过氧化氢酶(catalase,CAT)活性,取0.2 mL 上清液加入2 mL 磷酸盐缓冲液(phosphate buffered saline,PBS)溶液、0.3 mL 30%的H2O2于试管中,每隔1 min 测定240 nm 下吸光值,共测3 次。每个处理设3个生物学重复。
以上测定方法均参照《植物生理学实验指导》[24],并稍有改进。
1.2.3 小麦根尖细胞染色体观察 小麦根尖临时装片的制备方法参考王明玉等[25]的方法并有所改进。萌发的小麦根长至约2.0 cm 时切下约1.0 cm 根尖,在95%乙醇和冰乙酸以3∶1 比例配成的卡诺固定液中4 ℃下固定24 h后取出。蒸馏水洗涤3次,转移至预热的酸解液(1 mol·L-1HCl),60 ℃条件下酸解8~10 min。蒸馏水洗涤3 次,吸干根尖表面水分,在石碳酸复红染液中染色10 min,切取根尖分生组织,放置于载玻片上,盖上盖玻片,用镊子敲至雾状后镜检。每个剂量观察14 个根尖,每个根尖选取70 个细胞进行统计,每个处理选取根尖细胞1 000 个。按下列公式计算有丝分裂指数、染色体畸变率和微核率:
有丝分裂指数=有丝分裂细胞数/观察细胞总数×100%
染色体畸变率=染色体畸变细胞数/有丝分裂细胞数×100%
微核率=微核细胞数/观察细胞总数×100%。
1.3 统计分析
数据首先使用Excel 2013 进行分析,用SPSS 20.0对数据进行单因素方差分析(one-way ANOVA),采用P<0.05、P<0.01、P<0.001和P<0.000 1水平的t检验方法进行显著性分析,采用Graphpad Prism 7.0进行绘图。
2 结果与分析
2.1 电子加速器源射线对小麦生长特性的影响
不同辐射剂量处理下小麦种子发芽率无显著差异(图1-A)。开始萌发时,与对照相比,辐射处理种子发芽势较低,随着辐射处理剂量的增加,萌发第1 天发芽势分别下降49.0 和60.06 个百分点;萌发第2 天发芽势分别下降16.88 和27.54 个百分点;第3 天后各辐射剂量下发芽势无明显差异(图1-B)。表明开始时种子的生活力随辐射剂量增加而减弱,即高剂量的辐射处理可以延缓小麦种子的发芽。
随着辐射剂量的增加,小麦幼苗株高和根长显著降低(P<0.000 1,图2),200 Gy处理下小麦幼苗株高和根长与对照组相比分别降低49.67%和74.79%;400 Gy处理下分别降低了81.32%和89.67%说明辐射处理对小麦的生长有抑制作用,其中400 Gy 严重影响小麦幼苗的正常生长。

图2 不同剂量辐射处理对小麦幼苗株高与根长的影响Fig.2 Effects of different doses of irradiation on plant height and root length of wheat seedlings
辐射处理后,水培10 d 小麦的鲜重和地上生物量与对照组相比均有降低(图3)。200 Gy 处理下,小麦幼苗的鲜重和地上生物量分别降低50.93%和59.27%;400 Gy 处理下小麦幼苗的鲜重和地上生物量分别降低62.01%和75.26%。表明随着辐射剂量的增加,小麦幼苗的生长随之减缓,而地下生物量与对照组相比有小幅上升,分别升高12.89% 和23.22%。表明高剂量辐射处理下小麦种子的营养物质向地上的转化和运输受到明显抑制,因而表现在影响生长。
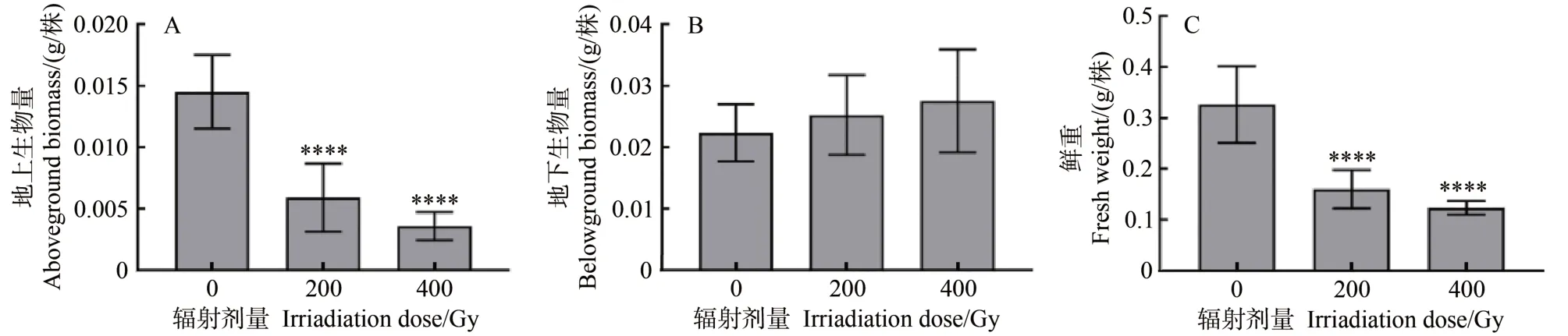
图3 不同剂量辐射处理对小麦幼苗生物量的影响Fig.3 Effects of different doses of irradiation on the biomass of wheat seedlings
2.2 电子加速器源射线对小麦生理特性的影响
辐射处理下小麦叶片中SOD活性随辐射剂量增加而降低,POD 活性和CAT 活性随辐射剂量增加呈先升后降趋势(图4)。与对照组相比,200 Gy 处理下叶片中POD活性和CAT活性分别升高89.77%和171.01%,SOD活性降低34.91%;400 Gy处理下POD活性和CAT活性分别升高59.74% 和37.69%,SOD 活性降低45.45%。由此可见,POD、CAT 为辐射处理后小麦幼苗清除活性氧的主要要抗氧化酶。

图4 不同剂量辐射处理对小麦叶片抗氧化酶活性的影响Fig.4 Effects of different doses of irradiation on antioxidant enzyme activity in wheat leaves
随着辐射剂量增大,叶绿素a和叶绿素b含量均呈降低趋势(图5)。与对照组相比,200 Gy 处理下小麦叶片中叶绿素a 与叶绿素b 含量分别降低42.55%和28.03%;400 Gy 处理下小麦叶片叶绿素a 与叶绿素b含量分别降低54.69%和54.44%。与200 Gy 处理相比,400 Gy处理下小麦叶片中叶绿素a与叶绿素b含量分别降低21.13%和36.70%。

图5 不同剂量辐射处理对小麦叶片叶绿素含量的影响Fig.5 Effects of different doses of irradiation on chlorophyll content of wheat leaves
随着辐射剂量的增大,小麦叶片中可溶性蛋白、MDA 和H2O2含量呈上升趋势(图6)。与对照相比,200 Gy 处理下小麦叶片中可溶性蛋白、MDA、H2O2含量分别升高63.20%、44.85%和120.64%;400 Gy处理下小麦叶片中可溶性蛋白、MDA、H2O2含量分别升高102.45%、75.36%和175.78%。

图6 不同剂量辐射处理对小麦幼苗生理特性变化的影响Fig.6 Effects of different doses of irradiation on physiological characteristics of wheat seedlings
随辐射剂量的增加,叶片超氧阴离子剂量升高(图7)。对照组中超氧阴离子含量较低,未发现明显染色部分,但200与400 Gy处理下可见蓝色色斑明显增大、增多。说明高剂量的辐射导致超氧阴离子积累。
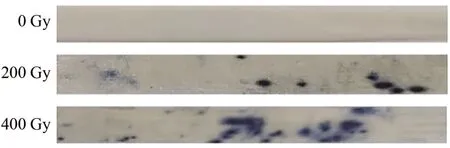
图7 不同剂量辐射处理对小麦叶片超氧阴离子含量的影响Fig.7 Effects of different doses of irradiation on the content of superoxide anion in wheat leaves
2.3 电子加速器源射线对小麦根尖细胞染色体的影响
辐射处理下小麦根尖细胞染色体发生断裂、染色体未排列在赤道板、染色体滞后、染色体桥、多极分裂及微核等突变现象(图8)。

图8 不同剂量辐射处理对小麦根尖染色体畸变的影响Fig.8 Effects of different doses of irradiation on chromosome aberration of wheat root tip
与对照组相比,随着辐射剂量增加,200 Gy处理下小麦根尖细胞染色体畸变率、微核率分别升高6.80 和6.02 个百分点,有丝分裂指数降低4.37 个百分点;400 Gy 处理下小麦根尖细胞染色体畸变率、微核率分别升高16.19 和14.22 个百分点,有丝分裂指数降低6.00 个百分点。表明辐射对染色体根尖有丝分裂有抑制作用,对染色体畸变有促进作用,通过影响根尖细胞正常分裂,抑制小麦幼苗正常生长。
3 讨论
植物的遗传变异特性最终通过植株的表型生长性状体现。本研究中电子加速器源射线对小麦种子的发芽率影响不明显,但随着辐射处理剂量的增加,小麦发芽及生长减缓,幼苗株高、根长、地上生物量及鲜重均呈现下降趋势,而地下生物量略有上升,表明高剂量(400 Gy)辐照处理下小麦幼苗的生长受到抑制,可能是萌发过程中种子中储存的营养物质向外转运减少,胚芽生长延缓。
植物细胞中丙二醛(MDA)含量可反映细胞膜受伤害程度,叶绿素和可溶性蛋白含量则分别是反映光合与渗透调节代谢的重要指标。本试验中随着辐射剂量的增加,小麦幼苗MDA 和可溶性蛋白含量增加,叶绿素和含量降低,这与李永锋[26]和韩榕等[27]辐射处理小麦的研究结果一致,说明电子束辐射处理小麦后引起细胞膜损伤及代谢紊乱,导致细胞中由多种酶系构成的非膜结合蛋白合成体系被激活,叶绿素合成受阻,对植物光合作用产生了不利影响。
正常生长的植物细胞体内活性氧的产生与清除处于平衡状态,而逆境下这种平衡被打破。本研究中随着辐射剂量的增加,小麦幼苗细胞中H2O2和超氧阴离子含量升高,表明辐射导致体内大量活性氧积累,细胞膜产生不可逆的氧化损伤。此外,随着辐射剂量的增加,SOD 活性持续降低;在200 Gy 辐射下CAT 和POD活性呈现上升趋势,400 Gy辐射下该酶活性略降,但仍大于对照组。说明低剂量(200 Gy)辐射处理下,细胞内产生的活性氧主要通过刺激CAT 和POD 活性来清除,以提高自身防御能力,保护细胞膜的完整性和正常代谢;但高剂量(400 Gy)辐射处理下,小麦叶片细胞膜损伤加剧,影响正常的生理代谢,导致抗氧化酶活性降低。综上所述,相对而言,200 Gy可作为为电子加速器源射线处理小麦的的参考剂量。
细胞染色体是遗传物质的载体,植物的遗传变异也体现在染色体的形态变化。本研究中在观察小麦根尖染色体时发现,200、400 Gy处理下,细胞有丝分裂指数分别下降4.37 和6.00 个百分点,畸变率分别升高6.80和16.19个百分点,微核率分别升高6.02和14.22个百分点,表明电子束辐射不仅能提高染色体畸变的频率,同时也可获得大量染色体畸变的类型,辐射剂量越大,对根尖细胞染色体的损伤越大。陆璃等[28]在γ辐射小麦的研究中指出,γ 辐射能够诱变小麦根尖染色体畸变,微核率也随辐射剂量增加而升高,与本研究结果相符。由此可见,高剂量辐射处理小麦种子抑制小麦生长,对小麦根尖细胞染色体造成了损伤,表明尖有丝分裂、提高染色体畸变率从而提高小麦的突变频率。综上所述,众多的变异类型则为品种选育增加新材料和新的遗传资源,为创制新品种提供可能。
本试验仅限于实验室水培植株,处理剂量有限,而在后续的研究中还应结合大田小区试验,通过观察小麦的生长表型,筛选有益突变体,缩短育种周期,提高育种效率。
4 结论
随着辐射剂量的增加,突变一代(M1)代小麦幼苗生长发育在一定程度上受到抑制和损伤,生长发育减慢,叶绿素含量降低,MDA 含量增高。辐射处理减少了小麦根尖细胞有丝分裂指数,减缓小麦根尖增长速率,增加小麦根尖细胞畸变率及微核率。本试验选取的辐射剂量中,200 Gy 可作为电子加速器源射线辐射小麦诱变育种的参考剂量。
猜你喜欢
今日农业(2021年10期)2021-11-27
阅读(科学探秘)(2020年8期)2020-11-06
科学之谜(2019年3期)2019-03-28
中国果业信息(2019年1期)2019-01-05
科学之谜(2018年8期)2018-09-29
生物学教学(2017年9期)2017-08-20
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
中国火炬(2015年12期)2015-07-31
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
食品工业科技(2014年6期)2014-05-10
