
黄土丘陵区苜蓿地土壤线虫群落结构和代谢足迹对磷素的响应
2023-09-26 01:02李亮亮罗珠珠李玲玲牛伊宁何仁元刘家鹤
植物营养与肥料学报 2023年8期
李亮亮,罗珠珠,*,李玲玲,牛伊宁,何仁元,刘家鹤
(1 甘肃农业大学资源与环境学院,甘肃兰州 730070;2 省部共建干旱生境作物学国家重点实验室,甘肃兰州 730070;3 甘肃农业大学林学院, 甘肃兰州 730070)
线虫(Caenorhabditiselegans)是土壤中数量和种类最为丰富的生物,占领着土壤食物网多个营养级地位[1-3],对调控有机物的分解、养分转化和能量传递起到关键的作用,是维持土壤生态系统稳定的重要组成部分[4-5]。线虫对环境扰动十分灵敏,其群落组成和结构对土壤环境变化做出的反应,为土壤生态过程提供了独特信息,是评价生态系统恢复与扰动状况的生物学指标[6-7]。线虫群落易受土壤水分[8]、养分含量[9]、pH[10]等因子调控,耕作、施肥等农艺措施改变了土壤环境因子,进而也影响了土壤线虫群落。
黄土高原地区水土流失和土壤退化问题非常严重,生态环境十分脆弱[11]。紫花苜蓿(Medicagosativa)自汉代从西域引入我国,至今已有2000 多年的种植栽培历史[12]。紫花苜蓿作为推广种植的优良豆科牧草,具有产量高、营养丰富、生态适应性强的特点,其根系强大的根瘤固氮作用,对增加土壤有机质含量,改善区域生态环境和促进畜牧业发展发挥了重要作用[13-15]。目前,甘肃省种植面积74.67 万hm2,总产量高达603 万t,居全国首位[16]。由于苜蓿生长对土壤水分和磷(P)素需求量大[17-18],持续多年种植会导致苜蓿产草量较低且生长旺盛期短[19],苜蓿人工草地退化,需要增施磷肥以维持其可持续利用[20]。但磷肥过高或过低均不利于根瘤菌的正常发育[21],因此,选择适宜的磷肥用量有利于紫花苜蓿生长并提高土壤肥力。
有研究表明,施磷可提高食细菌性线虫、捕食/杂食性线虫和植食寄生性线虫的丰度[22-23],显著提升食真菌线虫的丰度[24]。土壤、气候和植被类型也影响着不同供磷水平下的线虫群落结构[25-26]。本研究假设黄绵土区苜蓿种植过程中土壤线虫密度、多样性、群落结构和代谢足迹增加均受到了施用磷肥的影响,为验证该假设,本研究基于黄土高原半干旱雨养区的一个长期定位试验,调查了多元化线虫群落特征度量指标(多样性指数、生态功能指数、代谢足迹)对施磷的响应,为黄土高原苜蓿人工草地的可持续利用提供理论依据。
1 研究区域与方法
1.1 研究区概况
试验设在甘肃省定西市安定区李家堡镇甘肃农业大学旱作农业综合试验站(35°28′N,104°44′E)内。试验区地处中温带半干旱区,年均日照时数2476.6 h,年均太阳辐射592.9 kJ/cm2,≥0℃积温2933.5℃,≥10℃积温2239.1℃,无霜期140 天,年均气温6.4℃,年均降水量390 mm,年均蒸发量1531 mm,属典型的一年一熟旱作雨养农业区。土壤类型为黄绵土,耕层土壤有机质8.04 g/kg,全氮0.82 g/kg,全磷1.07 g/kg。
1.2 试验设计
试验始于2014 年4 月6 日,紫花苜蓿采用条播方式建植,播种量为22.5 kg/hm2,每块样地播种10 行,行距为 30 cm。共设4 个P2O5用量处理,分别为0 (CK)、60 kg/hm2(P60)、120 kg/hm2(P120)和180 kg/hm2(P180),每个处理3 次重复,共12 个小区,随机排列,小区面积均为12 m2(3 m×4 m)。磷肥每3 年施1 次(分别于2014、2017 和2020 年施用),氮肥每年施用1 次,施用量为N 50 kg/hm2。供试肥料为尿素(N46%)、过磷酸钙(P2O512%)。试验采用完全雨养条件,不进行灌溉。
1.3 土壤样品采集
于2021 年6 月苜蓿头茬盛花期采集土壤样品,在每小区内随机取 5 株苜蓿,使用土钻采集苜蓿根围0—20 cm 深的土样,混合成1 个样品,去除土样中的石砾和植物残渣等杂物。将每个土样分成两份,1 份放置于4℃冰柜,用于线虫的提取与鉴定,1 份风干后用于测定土壤理化性质。
1.4 土壤理化指标测定方法
土壤含水量采用烘干法[27]测定;pH 采用电位法测定[27];全氮(TN) 采用H2SO4消煮—凯氏定氮法测定[27];全磷(TP)和速效磷(AP)均采用比色法[27]测定;土壤有机碳含量采用外加热重铬酸钾氧化法[27]测定。
1.5 土壤线虫测定
1.5.1 土壤线虫群落鉴定 采用浅盘法提取土样线虫[3]。称取100.0 g 鲜土样铺设于浅盘内部的滤纸上,沿着浅盘内壁缓缓地注入自来水,直至土样表面出现一层稀薄状的水膜,于室温培养48 h,使用孔径25 μm 筛分离获取线虫样品,在60℃水浴条件下将其杀死后,用4%的福尔马林溶液将线虫体型固定,并装置于提前标记好试区序号的标本瓶内等待鉴定。用微量取液器吸取0.2 mL 待定溶液滴加至载玻片上,滴加时从载玻片一侧缓缓释放溶液至载玻片另一端,形成条带状且均匀的涂片。将制作好的载玻片放置在体式显微镜下观察线虫形态并计数。土壤线虫的分类及鉴定参照《中国土壤动物检索图鉴》[28]、《植物寄生线虫分类学》[29]以及加州大学戴维斯分校Ferris 教授建立的网页http://Nemaplex.ucdavis.edu。同时依据线虫头部的形态特征及其取食生境将其划分为4 种营养类群(食细菌性线虫、食真菌性线虫、植物寄生性线虫、杂食/捕食性线虫),并根据生活史策略,将线虫划分成 Colonizer (机会主义者,r-对策者)和 Persister (竞争者,k-对策者),给线虫赋予不同的c-p 值,从极端的c-p 值为1 的r-对策者向c-p 值为 5 的k-对策者将其划分成5 种类群[30]。
1.5.2 生态功能指数计算公式
式中,Pi指第i个线虫类群丰度与线虫总丰度的比值[31]。
式中,pi指第i个线虫类群丰度与线虫总丰度的比值[32]。
式中,BF 指食细菌性线虫丰度,FF 指食真菌性线虫丰度[33]。
式中,PP 指植物寄生性线虫丰度[33]。
式中,vi指自由生活线虫的c-p 值,fi指第i类线虫类群丰度占自由生活线虫丰度的比值[30]。
式中,fi指第i类PP 类线虫丰度占线虫总丰度的比例[30]。
1.5.3 线虫代谢足迹和区系分析 利用“线虫-植物专家信息系统”(http://plpnemweb.ucdavis.edu/nemaplex)中列出的线虫生物量 (鲜重,W)计算线虫代谢足迹。
式中,Nt指t类线虫类群丰度;mt指t类线虫类群cp 值;Wt指t类线虫类群生物量[34]。
根据代谢足迹 (Fe) 和结构代谢足迹 (Fs),以(SI,EI) 坐标点为中心,在A、B、C、D 象限内绘制4 个坐标点:(SI,EI-0.5Fe/k)、(SI+0.5Fs/k,EI)、(SI,EI+0.5Fe/k)和(SI-0.5Fs/k,EI),其中k 为转换常数[35]。线虫的功能代谢足迹为依次连接4 个坐标点所圈定的四边形面积。
1.5.4 线虫物种选择性 线虫物种选择性以IndVal值表示,IndVal 值根据以下公式进行计算[36]:
式中:Aij是线虫类群i在样地组j的平均多度(个体数)与该线虫类群在所有样地组的平均多度之和的比值;Bij是样地组j中存在线虫类群i的样地数与该样地组的所有样地数的比值。以线虫对环境的专一性和忠实性为基础,采用物种指示值分析法[37]计算土壤线虫指示值(soil nematode indicated value),并通过IndVal 值确定不同施磷素处理选择性物种(物种通常仅生长在添加磷素或未添加磷素处理)和非选择性物种(物种在添加磷素和未添加磷素处理均可生长)。如果IndVal 值大于0.6,则认为该物种是磷素选择性物种,否则为磷素非选择性物种;施磷素处理IndVal值大于0.6 为施肥正选择物种,CK 处理IndVal 值大于0.6 为施肥负选择物种。
1.6 数据处理
线虫生态功能指数、多度、营养类群和理化指标的差异显著性分析,利用SPSS 26.0 单因素方差分析(ANOVA) 和多重比较(LSD 法,P=0.05) 完成。主成分分析(PCA)采用R 软件“stats”程序包进行,通过降维定性分析样本间物种组成的相似性和差异性,找出影响样本物种组成差异的潜在主成分;为了进一步判断组间差异是否具有统计学意义,采用R 软件“vegan”程序包进行ANOSIM 分析检验差异显著性。Pearson 检验主要是通过相关性热图来分析线虫各营养类群与理化因子之间的相关关系。柱状图和小提琴图采用Original 2021 作图,主要分析线虫各营养类群代谢足迹大小,表明线虫对碳的利用强弱,AI (Adobe Illustrator CS6) 软件进行图形修饰。
2 结果与分析
2.1 土壤线虫群落结构组成
12 个样品共鉴定出土壤线虫(Caenorhabditis elegans) 4575 条,每个样品线虫密度平均达381.25条/(100 g,干土),隶属于19 科27 属,包括食细菌性线虫和植物寄生性线虫各9 属,食真菌性线虫4属,捕食/杂食性线虫5 属 (表1),以植物寄生性线虫占比最高(43.00%~57.33%)。不同处理土壤线虫优势属(>10%)、常见属(1%~10%)和稀有属(<1%)分布有所差异,螺旋属为各处理共有优势属(20.67%~39.33%)。除此之外,CK 处理的优势属还包括拟丽突属(10.33%)、艾普鲁斯属(11.00%)和丝尾垫刃属(10.00%);P60 处理优势属还有拟丽突属(11.00%);P120 处理优势属有艾普鲁斯属(10.00%);P180 处理优势属有中杆属(10.00%)。土壤线虫总多度各处理间无显著差异(图1),但是不同营养类群相对丰度在处理间差异显著(图1),其中CK 和P60 处理食细菌性线虫相对丰度显著高于P120 和P180;P60 处理食真菌性线虫相对丰度显著低于其他处理;P180 处理植物寄生性线虫相对丰度显著高于其他处理;P60和P120 处理杂食/捕食性线虫相对丰度显著高于CK和P180 处理。另外,P60 处理的线虫丰富度显著高于其他处理(图1)。
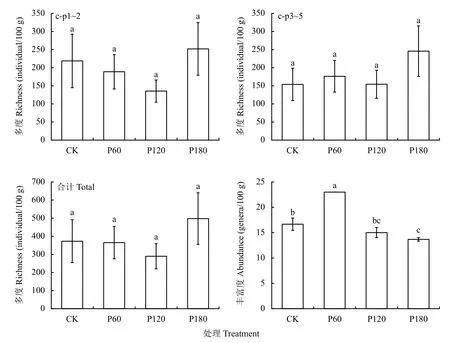
图1 土壤线虫丰富度和多度Fig.1 The richness and abundance of soil nematodes
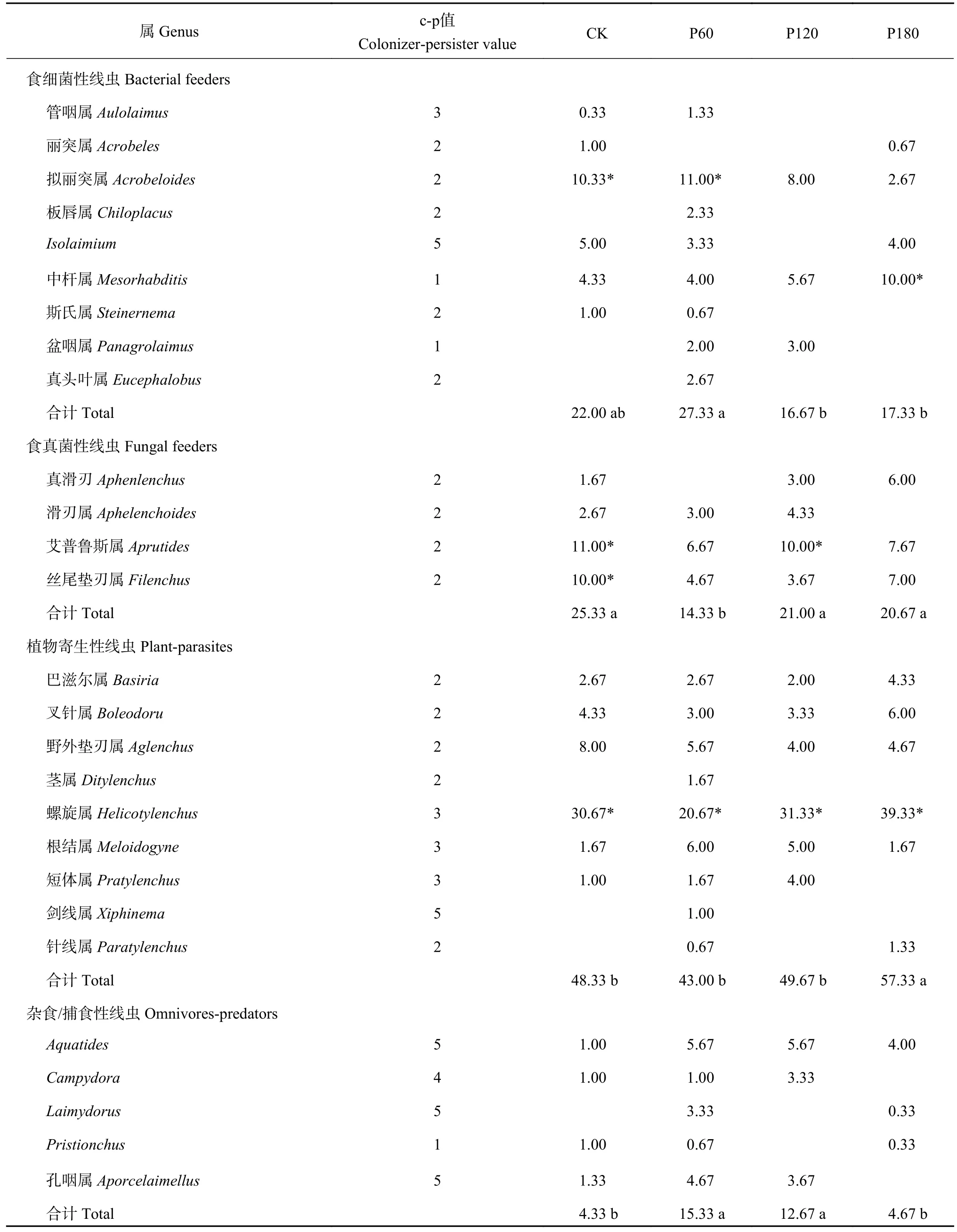
表1 土壤线虫群落组成(%)Table 1 Composition of soil nematode community
主成分分析结果(图2)表明,土壤线虫各营养类群、Colonizer Persister (c-p) 类群和总线虫群落的差异总解释度均大于80%,且R>0,说明不同供磷素水平下土壤线虫群落结构存在差异。ANOSIM检验发现,P 素添加显著影响食微类群、杂食/捕食类群、c-p1~2 类群和总线虫群落。
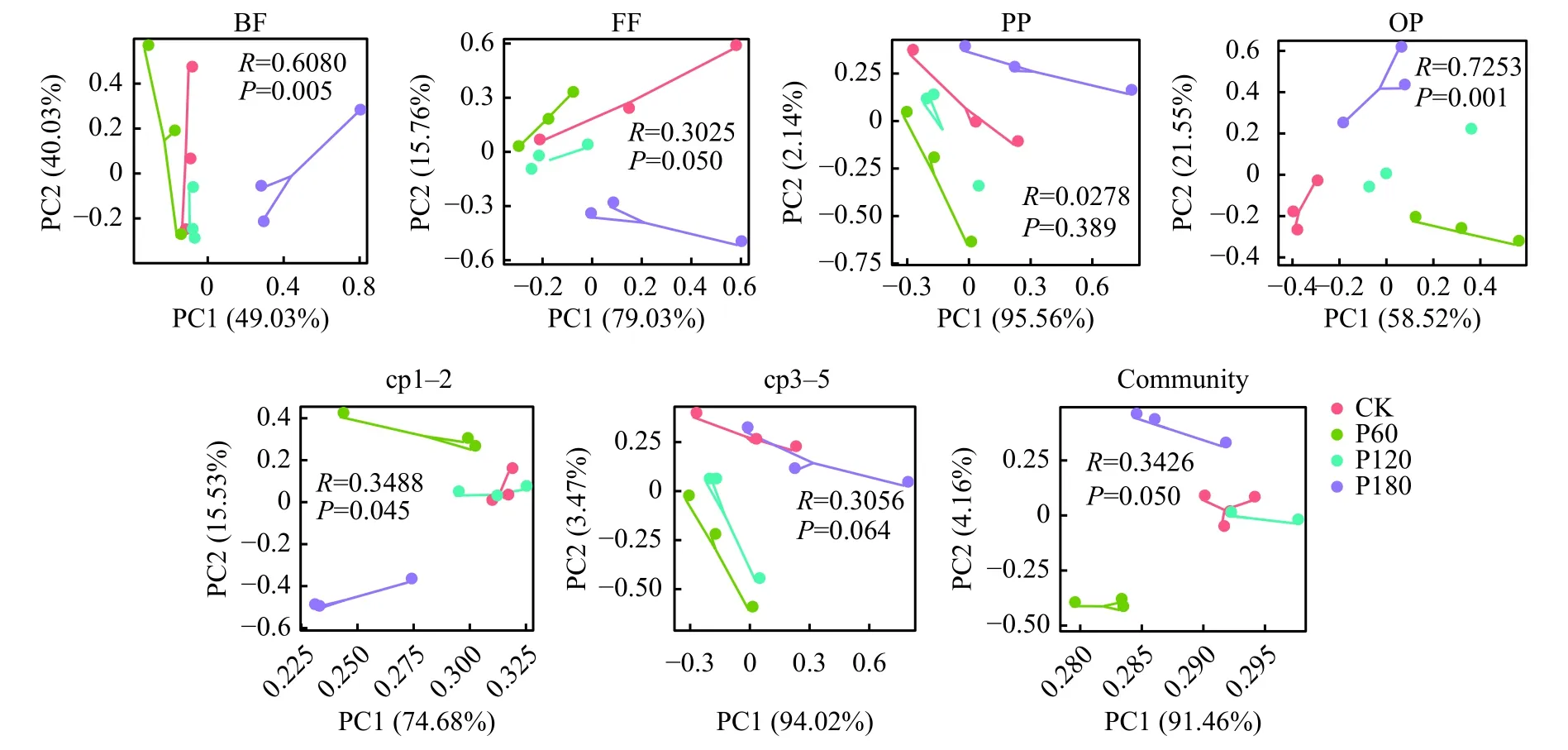
图2 土壤线虫营养类群、c-p 类群和总线虫群落的PCA 分析Fig.2 PCA analysis of soil nematode trophic groups, c-p groups and total nematode communities
2.2 土壤线虫生态功能指数
土壤线虫群落生态功能指数分析结果(表2)发现,除瓦斯乐卡指数(WI)之外,其余生态功能指数在各处理之间均存在显著差异(P<0.05)。P60 处理多样性指数(H′)、线虫通路指数(NCR)和自由生活线虫成熟度指数(MI)均显著高于其他处理(P<0.05),植物寄生性线虫与自由生活线虫成熟度指数比(PPI/MI)和优势度指数(λ)显著低于其它处理(P<0.05);P120 处理植物寄生性线虫成熟度指数(PPI)显著高于其它处理(P<0.05)。
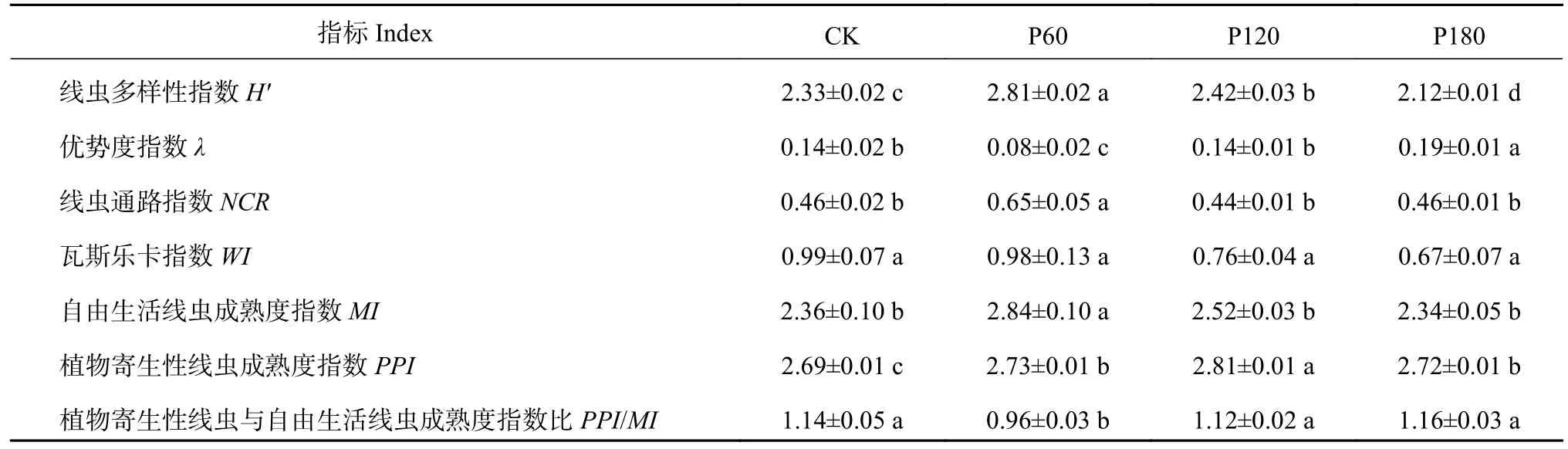
表2 不同施磷水平下土壤线虫群落指数Table 2 Soil nematode community indices as affected by P2O5 application rate
2.3 土壤线虫代谢足迹和区系分析
施磷量对杂食/捕食性线虫代谢足迹(OPMF)有显著影响,对其余各营养类群代谢足迹均无显著影响(图3)。P60 处理的OPMF 显著高于其他处理(P<0.05),P120 处理的OPMF 显著高于CK。区系分析显示(图4),CK 处理土壤线虫分布在A、B、C 三象限,P60、P120 和P180 处理均处于B 象限。土壤线虫功能足迹(菱形面积)表现为P60>CK>P180>P120;土壤线虫富集指数(菱形中心位置)随施P 素量增加而提高。
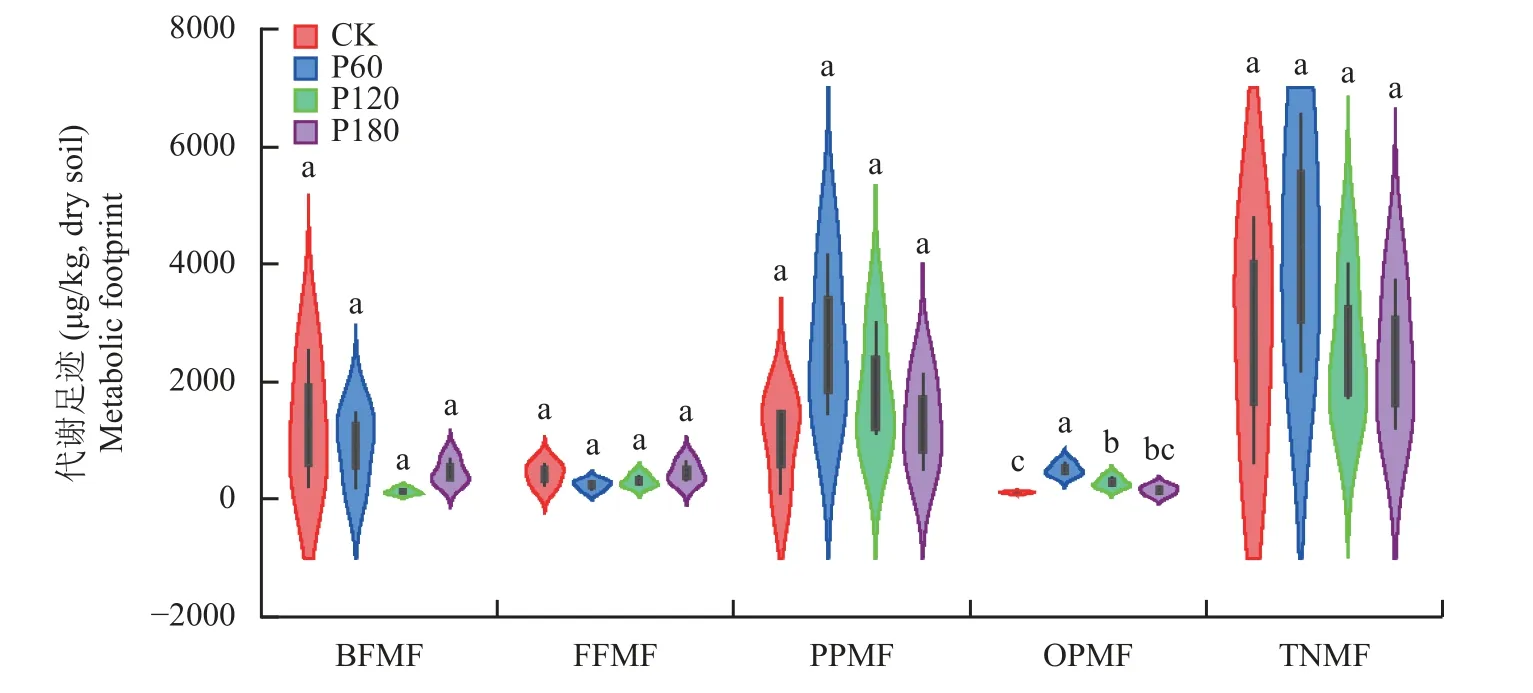
图3 土壤线虫代谢足迹Fig.3 Metabolic footprint of soil nematodes
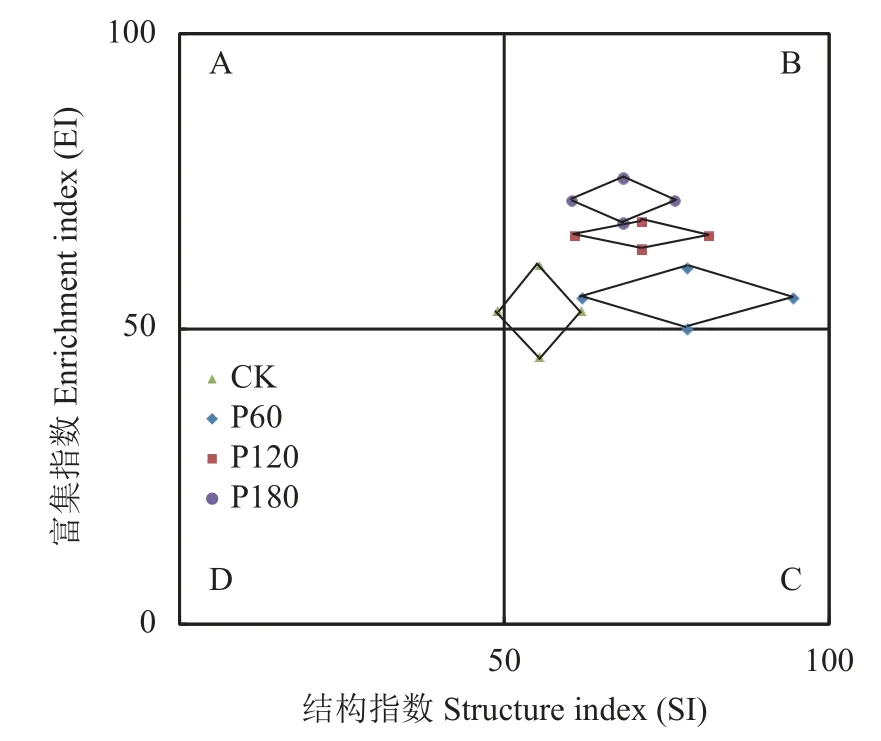
图4 土壤线虫群落区系分析Fig.4 Fauna analysis of soil nematode communities
2.4 不同生境对土壤线虫群落物种的选择性
施磷对线虫营养类群和c-p 类群具有选择性(表3),其中P60 处理除了食真菌类线虫外,其余各类线虫正选择性物种比例大于负选择性物种比例;P120 处理各类线虫正选择性物种比例大于负选择性物种比例,且食细菌类线虫负选择性物种比例为零;P180 处理食细菌类和植物寄生类线虫的正选择性物种比例和负选择性物种比例均为零,杂食/捕食类和c-p3~5 类线虫负选择性物种比例为零。食细菌类、杂食/捕食类和植物寄生类表现为P60 处理正选择性物种比例最大;食真菌类表现为P180 处理正选择性物种比例最大;c-p 类群均表现为P60 处理正选择性物种比例最大。
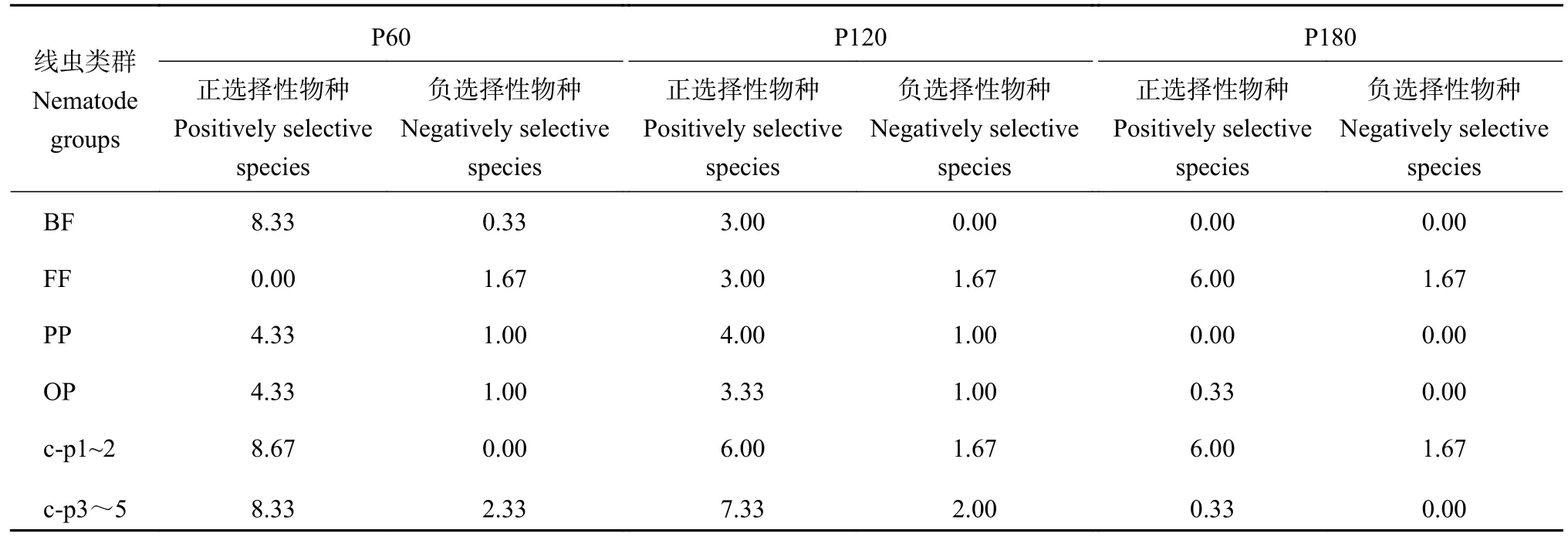
表3 线虫选择性物种比例(%)Table 3 Proportion of selective species of nematodes
一般将土壤线虫群落指示值大于0.6 的线虫属作为指示物种[37-39],在鉴定的27 个物种中,有9 个可以作为指示物种(表4)。P60 处理的指示物种有6个,分别为管咽属(Aulolaimus)、板唇属(Chiloplacus)、真头叶属(Eucephalobus)、茎属(Ditylenchus)、剑线属(Xiphinema)和Laimydorus;P120 处理指示物种只有3 个,分别是盆咽属(Panagrolaimus)、短体属(Pratylenchus)和Campydora,CK 和P180 处理无指示物种。
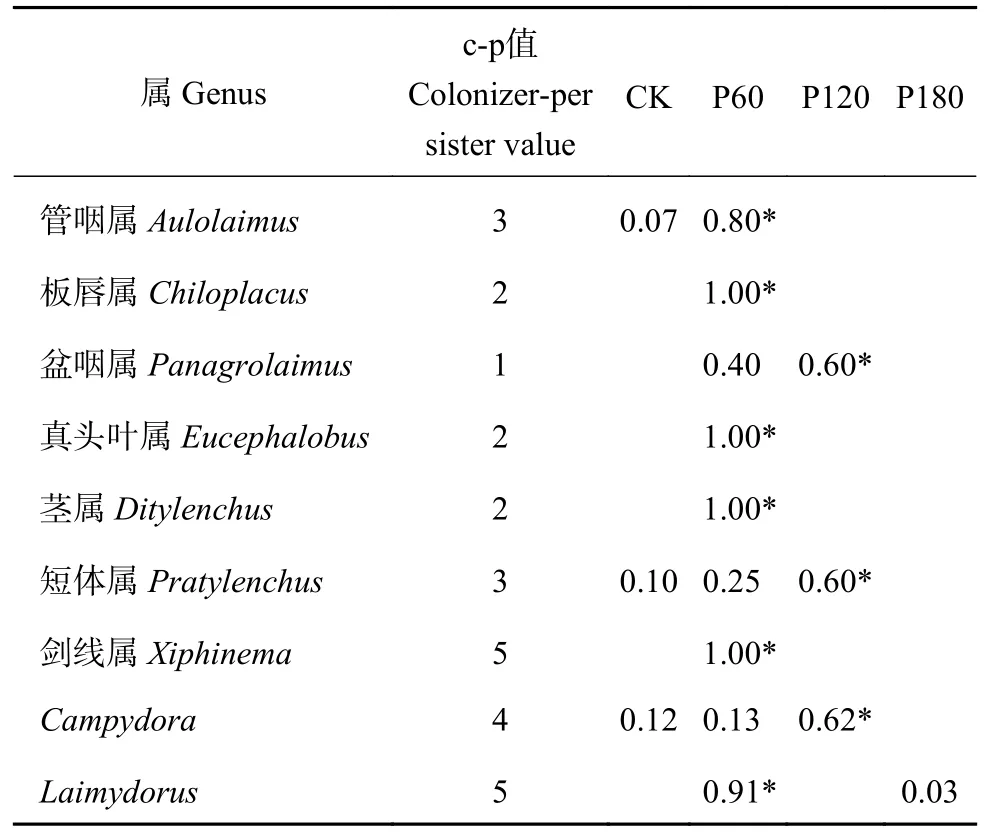
表4 不同处理中土壤线虫群落指示属及其物种指示值Table 4 Indicator genera and the indicated values (IndVal)of soil nematodes community in each treatment
2.5 土壤线虫群落与土壤理化性质的相关性
表5 显示,施磷量对土壤水分和全氮含量无显著影响,但显著提高了土壤中有机碳、全磷和速效磷含量。与CK 相比,P60 显著提高了土壤有机碳、全磷和速效磷含量,而对pH 无显著影响。P120 和P180 之间土壤有机碳、全磷和速效磷含量无显著差异,但均显著高于P60 处理,且P180 处理的pH 显著低于CK 和P60 处理(P<0.05)。
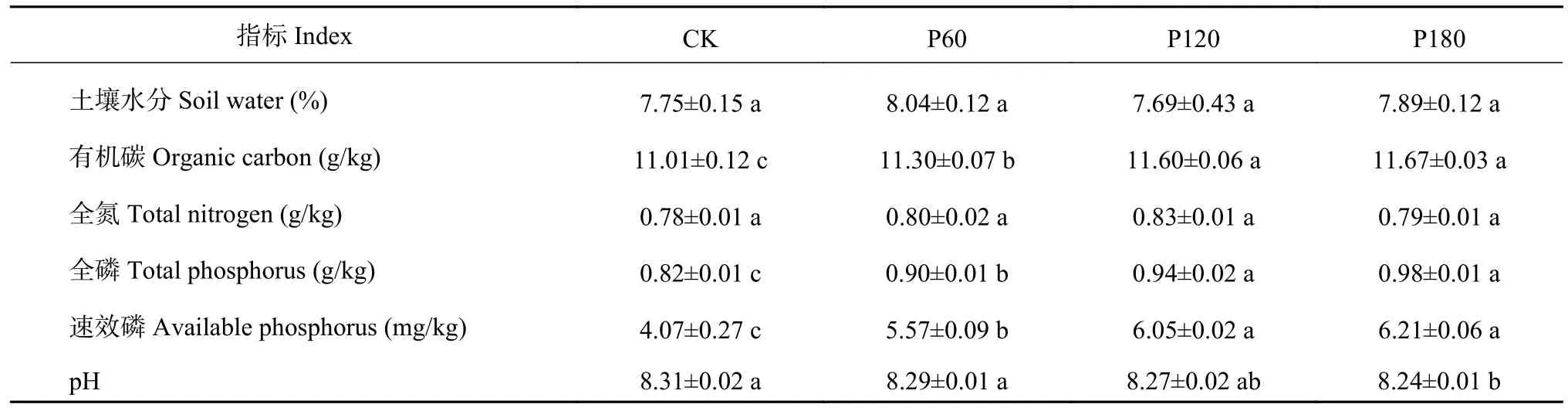
表5 不同施磷量下的土壤理化性质Table 5 Soil physiochemical properties under different phosphorus treatments
线虫群落相对丰度和土壤理化因子相关性分析(图5)表明,植物寄生类线虫与pH (P<0.01)呈显著负相关;杂食/捕食类线虫与全氮(P<0.05)呈显著正相关,单个土壤理化因子与食微线虫相关性均未达显著水平, 说明土壤食微线虫群落受到多种土壤理化性质的共同影响。
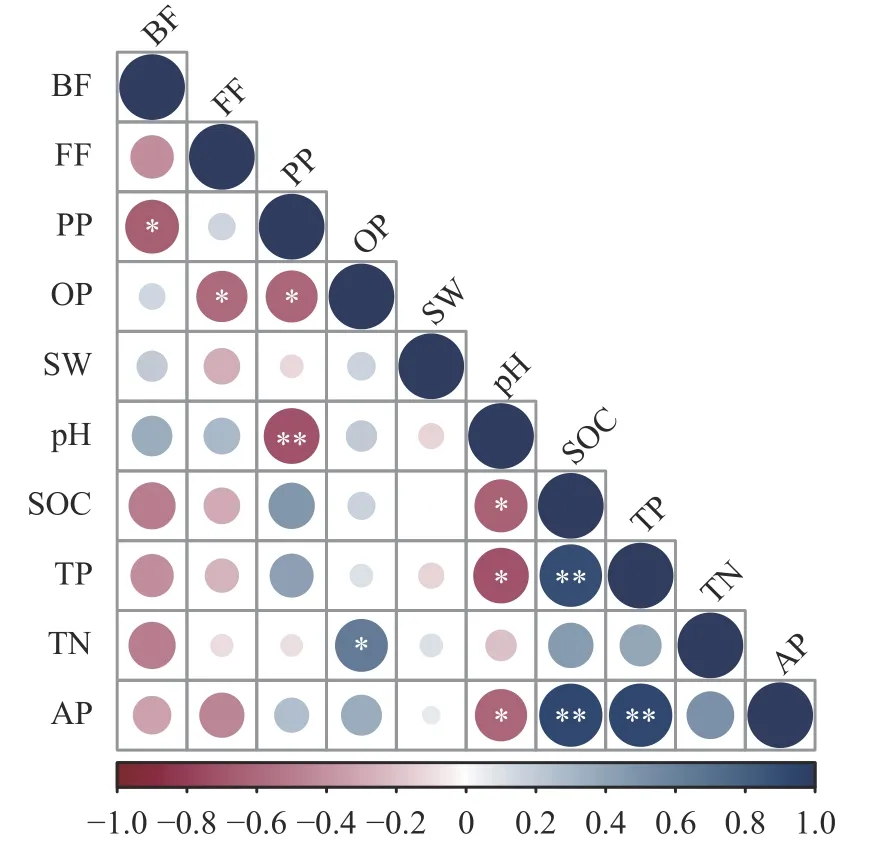
图5 土壤线虫群落和理化性质相关性分析Fig.5 Correlation analysis between soil nematode community and physical and chemical properties
3 讨论
本研究结果表明长期施磷对土壤线虫多度无显著影响,但影响着土壤线虫多样性、丰富度和群落组成,因为施磷引起土壤理化性质变化,而不同线虫群落对环境具有物种选择性亦即具有偏好或排斥性,导致土壤线虫群落组成差异较大,进一步的ANOSIM 检验证实施磷显著影响食细菌类群、食真菌类群、杂食/捕食类群和c-p1~2 类群线虫,这也验证了本研究假设。CK 处理无指示物种,而添加P 素后具有明显的指示物种,这表明长期P 素添加会改变土壤特定环境,线虫会寻找自己适合生存的地方,从而导致指示物种丰度在不同P 素处理间的差异。从营养类群水平而言,磷素添加之后各营养类线虫正选择物种比例大于负选择性物种比例,P60处理食细菌类、植物寄生类和捕食/杂食类线虫正选择物种比例最大,P180 处理食细菌类、植物寄生类和捕食/杂食类线虫正选择物种比例最小。这说明低浓度P 素可能更有利于食细菌类、植物寄生类和捕食/杂食类线虫生活,尤其是管咽属(Aulolaimus)、板唇属(Chiloplacus)、真头叶属(Eucephalobus)、茎属(Ditylenchus)、剑线属(Xiphinema)和Laimydorus,而高浓度磷素不利于这类线虫生存。这可能与高浓度磷素对土壤微生物的调控有关,高浓度P 素会抑制细菌群落的生长代谢,从而降低细菌的代谢功能活性,使土壤中细菌数量减少[40],因此食细菌性线虫的食物资源减少,所以食细菌性线虫比例有所降低。同样,高浓度P 素既抑制紫花苜蓿生长和叶片的光合能力,又抑制根瘤固氮能力,所以高浓度磷素不利于紫花苜蓿根系生长[41],更不利于植物寄生类和捕食/杂食类线虫生活。另外,对于食真菌类线虫,P180处理正选择物种比例最大,负选择物种比例与P60和P120 处理无差异,这可能是因为高浓度磷素会导致土壤中真菌数量增加[42],而真菌是食真菌性线虫的食物资源,所以食真菌性线虫比例有所增高。土壤线虫营养类群对土壤理化因子的响应不一致,影响植物寄生类线虫的主要理化因子为pH,因为植物寄生类线虫对土壤pH 的变化比其他类群线虫更敏感[43];影响捕食/杂食类线虫的主要理化因子为全氮,这可能是因为土壤线虫栖息环境偏好不同,食性不同以及生态位所导致的营养层次不同。
土壤线虫生态功能和代谢足迹发生变化主要是长期P 素添加造成土壤线虫群落结构差异所导致的。P60 处理土壤线虫多样性指数(H′)显著高于其他处理,优势度指数(λ) 显著低于其他处理,P180处理与之相反,这一结果与 Aerts 等[44]研究结果类似,说明适宜的磷素添加可以改善土壤线虫群落多样性,而过高浓度磷素则抑制土壤线虫群落的多样性。这可能是低浓度磷素能够促进紫花苜蓿根系生长,来吸收土壤中更多有效磷,同时根系会产生分泌物使酸性磷酸酶活性增强,复杂的有机磷转换为有效磷以适应和改善低磷生长环境[45],同时根系分泌物中含有一种黄酮醇能够促进根瘤菌固氮能力和植物的生长发育[46]。另外,根系分泌物会对土壤微生物产生影响,微生物多样性进一步影响线虫多样性[47],但二者明确的互作关系还有待后续联合代谢组和基因组进一步研究。尽管土壤线虫不直接以土壤C、N、P 素为营养来源,但低浓度磷素(导致C/P 和N/P 升高)显著增加了土壤线虫多样性,因此化学计量比表征的土壤环境和土壤微生物的改变会间接影响土壤线虫可用资源多样性[48],最终影响土壤线虫多样性。这可能是因为土壤C、N、P 素的循环不仅仅影响土壤食物网中的生物类群,而且对植物群落和以植物作为营养来源的土壤生物的生长、繁殖以及活动也产生了较大影响。MI指数越大,PPI指数越小,表明土壤线虫的健康状况越好,环境受干扰强度越小,线虫群落更为稳定[49-50]。值得注意的是,磷素添加后PPI指数显著增加,这可能是因为磷素添加后增加了土壤养分,有利于苜蓿根系生长以及促进植食性线虫繁殖,从而使PPI 指数升高;但P180处理PPI有所降低,可能是因为高浓度磷素使土壤酸化,对植物寄生性线虫造成了毒害作用使其数量减少[51],特别是短体属(c-p3)线虫缺失是植物寄生性线虫成熟度指数PPI降低的主要因素。但是,自由生活线虫成熟度指数MI随磷素添加量的增加先升后降,这也直接导致P60 处理植物寄生性线虫与自由生活线虫成熟度指数比(PPI/MI) 显著低于其他处理,说明与P120 和P180 处理相比,P60 处理土壤线虫群落受干扰程度显著降低,有利于其线虫群落向成熟稳定方向发展。
线虫代谢足迹能够指示土壤食物网碳代谢途径及其能流状态[35]。本研究植物寄生性线虫代谢足迹大于食微线虫足迹,表明食物网中从植物通道进入土壤的养分最多。进一步的土壤线虫区系分析结果表明,CK 处理横跨A、B、C 三象限,添加P 素处理均在B 象限,且土壤线虫富集指数(菱形中心位置)随磷素添加量增加而提高,说明P 素添加具有富集土壤养分和改善土壤环境效应,使得土壤干扰程度降低,食物网结构更成熟稳定。土壤线虫功能足迹表现为P60>CK>P180>P120,说明P60 处理土壤线虫群落对碳的利用率更高,其对食物网的调控以及维持生境中被捕食者与捕食者种群间代谢平衡关系的能力最强,这与其较高的杂食/捕食性线虫丰度有关。
4 结论
黄土高原半干旱区苜蓿地以植物寄生性线虫在数量上占据优势地位。施磷量提高了土壤有机碳、全磷、速效磷水平,而降低了土壤pH,引起土壤线虫群落结构、生态功能指数和杂食/捕食性线虫代谢足迹的改变。随着P 素添加量增加,植物寄生性线虫成熟度指数和线虫富集指数显著增加,但低磷量显著增加土壤线虫丰富度、多样性指数、线虫通路指数和自由生活线虫成熟度指数。土壤线虫功能足迹表现为P60 处理高于其他处理,说明P60 处理土壤线虫群落对碳的利用率更高,这与其较高的杂食/捕食性线虫丰度有关。因此,每3 年施用P 60 kg/hm2可以促进土壤食物网向复杂稳定方向发展,对于苜蓿人工草地的可持续利用具有积极意义。
猜你喜欢
文化遗产(2023年2期)2023-10-06
农业工程学报(2022年5期)2022-06-22
中国服饰(2021年5期)2021-06-30
江苏农业科学(2019年5期)2019-09-02
广东农业科学(2017年5期)2017-08-29
商情(2017年10期)2017-04-30
应用海洋学学报(2014年1期)2014-11-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
土壤与作物(2013年4期)2013-03-11
植物营养与肥料学报(2012年3期)2012-10-26
