
模拟增温对无芒雀麦生长特性的影响
2023-09-25 03:42:24宫珂靳瑰丽刘文昊马建刘智彪李嘉欣李莹
草业学报 2023年9期
宫珂,靳瑰丽,刘文昊,马建,刘智彪,李嘉欣,李莹
(新疆农业大学草业学院,新疆草地资源与生态重点实验室,西部干旱荒漠区草地资源与生态教育部重点实验室,新疆 乌鲁木齐 830052)
全球气候正在发生波动性变化,并加剧了极端气候出现,政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)2021 年报道中指出,由于人为活动致使全球地表气温升高的局面已无可争议,从1850 至1900 年气温约升高1.1 ℃,如果二氧化碳排放不加以控制,未来20 年增温将达到或超过1.5 ℃[1]。野生牧草是我国重要的生物种质资源,对维持草原生物多样性和保持草原生态平衡具有十分重要的作用,其种质资源具有适应性强、抗性耐性较好等栽培品种无法比拟的优点[2]。近20 年来我国草地生态不断受到过度放牧的人为破坏,以及气候变化带来的严重影响,使得种质资源及生物物种多样性面临严峻考验[3-4],有效地保护和利用野生牧草的遗传资源显得格外重要。已有大量研究表明,温度是植物生长繁殖过程中的关键环境因子,进一步研究发现增温对垂穗披碱草(Elymus nutans)、羊茅(Festuca ovina)、矮生嵩草(Kobresia humilis)等野生牧草的生长产生促进或抑制作用[5-6]。因此,研究野生牧草生长对温度升高的响应表现,可以为预测未来气候改变的背景下牧草的发展趋势提供理论依据[7-8]。
对有性繁殖植物而言,种子萌发、幼苗生长、繁殖生长是实现生活史的重要阶段。但是由于不同阶段植物形态的差异,受温度影响的程度各不相同。研究者发现,在植物生活史的前期阶段对环境变化的敏锐性更强,种子需要适宜的温度打破休眠状态,温度的升高会使有些种子萌发时间提前,种子出苗率提高[9],但对于需要低温环境来打破休眠的种子来说,增温反而会缩短低温时间,导致种子无法萌发[10]。植物成功出苗后,随即而来的是脆弱的幼苗生长阶段,地上部分和地下部分的生长反映出植物对资源的分配策略,面对温度的变化,植物会表现出最适生长策略,包括等速生长和异速生长等模式,目的是吸收更多养分以保证存活[11]。研究结果显示,温度增加能够同时促进幼苗地上和地下部分的生长[12],但也有研究发现,该环境下幼苗更倾向于将资源投入到地上部分[13]。植物繁殖生长阶段是生活史中较为稳定的时期,对环境因素的响应主要体现在营养生长和生殖生长权衡策略方面。增温在一定程度上提高了环境对植物热量的供给,直接或间接地影响植物在繁殖阶段中对各构件资源的分配[14]。在这类研究中得出的结论不尽相同,有的研究者发现增温会提前植物结实期的物候,为种子生长提供更多的物质积累,从而提高植物对生殖生长的投入[15];而有些研究者却得出在幼苗稳定建植后植物利用增温环境有效积累地上地下能量,从而对营养生长投入的资源大于生殖生长[16],甚至还有研究认为增温对禾本科植物的生长不具有影响[8]。
无芒雀麦(Bromus inermis)作为新疆广泛分布的优良野生牧草,是培育措施匮乏的天然草地中不可或缺的植物,其种质资源也可收集起来作为栽培牧草的来源。在前人的研究中发现,无芒雀麦可以通过改变表型性状和生物量分配来达到定居、扩张等一系列生命活动,而在混播当中也能占据合适的生态位[17-18]。在刈割和切根的人为胁迫过后能通过对影响因素的快速响应,积极调整自身结构使其避免生长受阻[19]。由此可见,无芒雀麦具有良好的表型可塑性,这种能力在自然情况和人为参与情况中都有所表现。然而,在全球气候变暖的影响下,无芒雀麦如何通过调节自身资源分配以及各构件如何协调生长来适应环境因素的变化还尚未明确。
因此,本研究通过野外模拟增温试验,探究无芒雀麦个体在发芽期、幼苗期和繁殖期的生长特性,明确其种子发芽、幼苗生长、繁殖分配的响应模式,再通过综合评价反映无芒雀麦自种子发芽至繁殖个体成熟阶段对增温的适应性表现,研究结果可为野生禾本科牧草的合理利用和有效保护提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于乌鲁木齐南山谢家沟试验站(87°02′24″ E,43°30′07″ N),海拔1670 m;位于天山北坡中部,低山带的山溪坡阶地,属于草甸草原,年平均降水量388.7~535.9 mm,年平均蒸发量1141.7~1564.9 mm;年平均气温2.1~3.3 ℃,无霜期100~113 d;土壤为山地栗钙土和山地黑钙土;气候类型属于半干旱草原气候。该区域是无芒雀麦集中分布的典型区域,且无芒雀麦已成为该群落的优势种。
1.2 试验处理
增温装置选择开顶式增温箱(open-top chambers,OTC),其上端开口,箱体为正六边形,由透明亚克力板材料(透光率可达90%)制成。OTC 装置通过改变箱体高度和底部面积,形成不同增温效果。
试验共设置3 个处理(图1)。对照处理(CK):不设置OTC 增温装置,选择1 m×1 m 样方,保持植被自然状态;W1增温处理:自然状态下利用OTC1装置进行增温;W2增温处理:自然状态下利用OTC2装置进行增温。每种增温处理3 个重复,间距2 m,对照处理与增温装置距离≥5 m。于试验开展期间定期对CK 样方、W1处理、W2处理内5、10 cm 土壤温度进行测量(9:00、13:00、17:00、21:00),计算增温效果,增温装置规格及增温效果见表1。

表1 增温设置规格及增温效果Table 1 Temperature increase specifications and effect
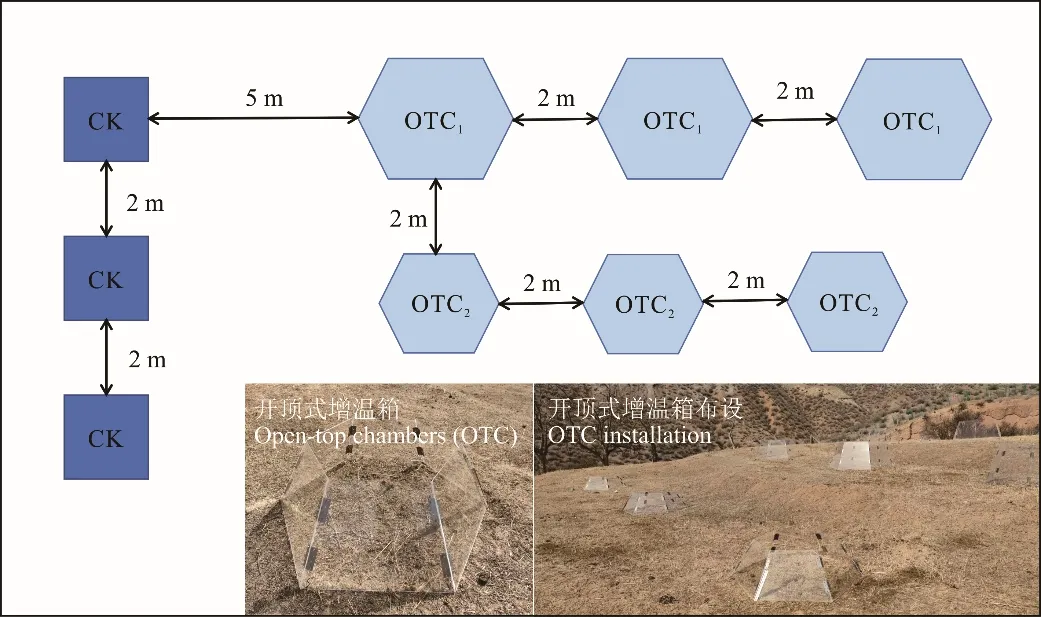
图1 增温装置布设Fig.1 Layout of the heating device
1.3 试验设计
1.3.1 种子发芽试验 挑选籽粒饱满、健康的无芒雀麦种子,播种于装有试验土的育苗杯(直径11 cm;高18 cm)内,底部制排水孔。试验土为原生境表层10 cm 土壤和花土1∶1 混匀,为了避免土壤种子库对试验结果造成影响,将土壤过1 mm 筛后放入100 ℃烘箱中48 h,每杯装入试验土400 g。按50 粒·杯-1标准均匀播种,播种深度为2 cm。
于2021 年4 月5 日将育苗杯放入增温装置,各重复的装置内埋置3 个育苗杯,即每个处理共9 个育苗杯,育苗杯的埋深与杯内土壤持平。根据监测站提供的1990-2019 年月降水数据,计算出4 月月均降水量为39.73 mm,按6 次·月-1的降水频率,每次浇水6.62 mm。降水时将装置开口处进行遮挡。共观察30 d,每天记录发芽数。
1.3.2 幼苗生长试验 采用盆栽法进行育种,试验土规格同种子发芽试验土,盆直径25 cm,底部具排水孔,按50 粒·盆-1播种。于种子发芽试验同期播种,根据月均降水量,每2 d·次-1进行浇水,直至种子萌发。种子萌发后长至二叶期,定苗至10 株·盆-1,缓苗5 d 后即2021 年5 月1 日进行温度处理,每个重复内埋置3 盆,各处理共9盆。增温处理时间为30 d,同样计算出5 月月均降水量为40.97 mm,单次浇水6.83 mm,雨天进行遮挡。试验结束后随机选取10 株健康完整的幼苗用于指标测定。
1.3.3 繁殖生长试验 于2021 年3 月在研究区内无芒雀麦群落布设OTC 装置,在距离增温装置>2 m 的位置选择3 个1 m×1 m 的样方作为对照组(CK)。持续观察至2021 年8 月,于无芒雀麦繁殖期进行取样。
1.4 测量指标
1.4.1 种子发芽测量指标 测定指标及计算方法如下:
式中:N1,N2,N3,…,Nn分别指第1 天、第2 天,第3 天,…,第n天的种子发芽率。
1.4.2 幼苗生长测量指标 每个处理随机选取10 株健康完整的幼苗,测量根长、叶长、根生物量、叶生物量,并计算根苗比、根冠比、地上生物量分配、地下生物量分配。
1.4.3 繁殖生长测量指标 每个处理挑选10 株健康完整的无芒雀麦单株,测量指标分为生殖指标(千粒重、生殖投入、生殖生长比率)、营养指标(株高、根冠比、营养投入、茎粗、分蘖数、叶长、根长)。
1.5 数据处理
使用Excel 2019 进行数据整理,运用SPSS 24.0 进行有重复观测值的单因素方差分析(one-way ANOVA),检验不同处理下萌发指标、生长指标的差异显著性,通过最小显著差异法(least significant difference,LSD)进行多重比较,所有结果由平均值±标准误(means±standard error)表示。使用SigmaPlot 14.0 制图。
2 结果与分析
2.1 模拟增温对无芒雀麦种子发芽的影响
不同模拟增温处理对种子发芽特性均具有影响(图2),W1增温处理的发芽率相较于CK 显著降低41.5%,对发芽势和发芽速率不具有显著影响(P>0.05);W2增温处理显著提高了发芽势和发芽速率(P<0.05),分别为CK 的1.38 和1.49 倍。从发芽率、发芽势和发芽速率的关系可以看出(图3),和CK 相比,W1和W2增温处理下发芽势和发芽率的关系发生变化,表现为种子发芽数量变多,但时间不集中;发芽速率和发芽率的关系在3 种处理下趋势一致且关系更为密切,表明增温可有效提高发芽速率,表现为发芽又多又快的趋势。
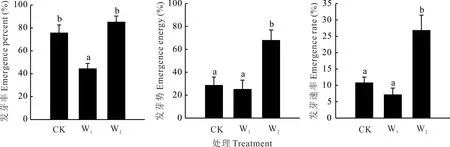
图2 野外模拟增温对种子发芽特征的影响Fig.2 Effects of field simulated warming on seed emergence characteristics
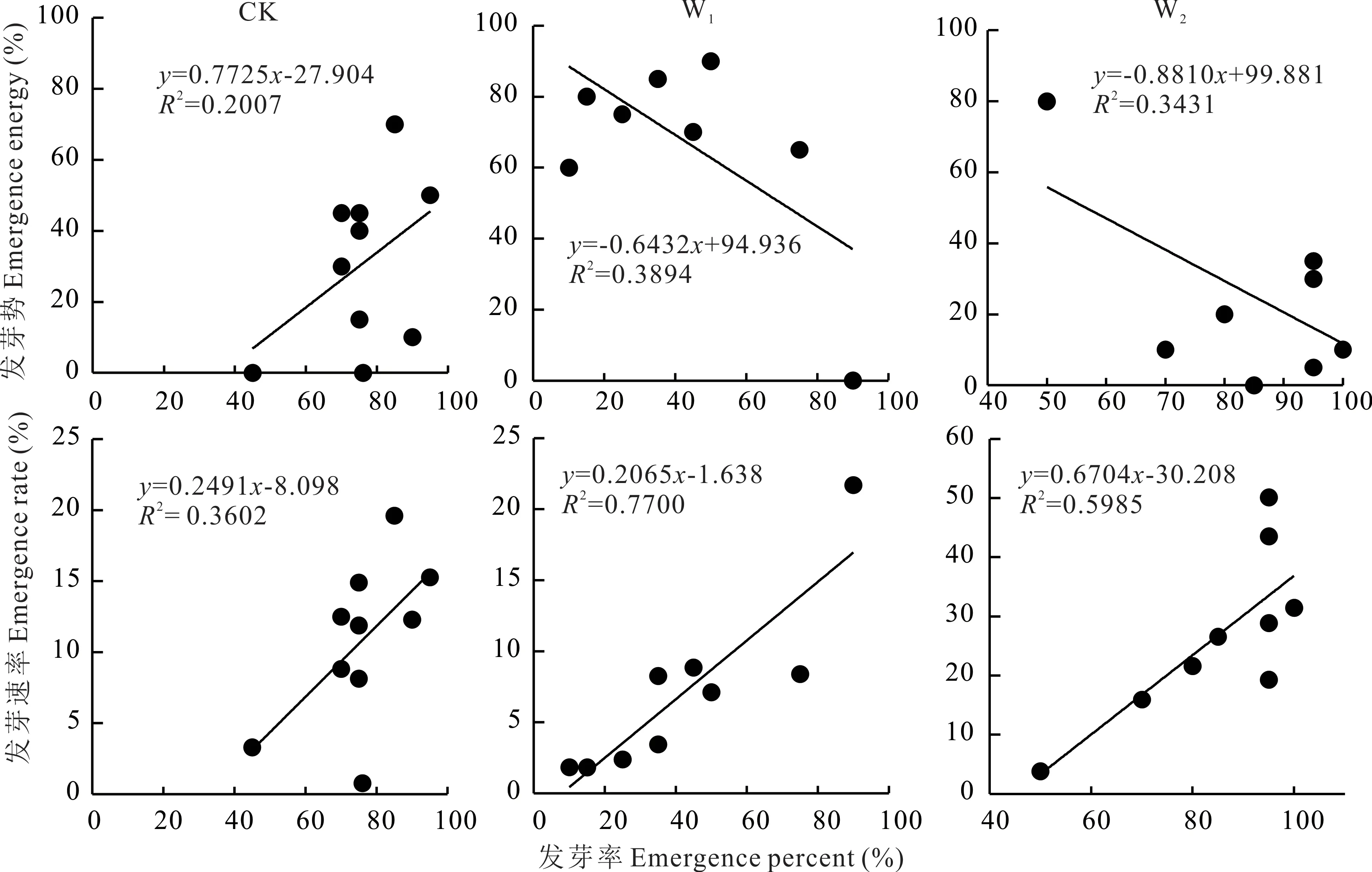
图3 野外模拟增温对发芽率与发芽势、发芽速率关系的影响Fig.3 Effect of field simulated warming on the relationship between emergence percent, emergence energy and emergence rate
2.2 模拟增温对无芒雀麦幼苗生长特性的影响
模拟增温处理对幼苗的生长起到抑制作用(图4),与CK 相比,叶长和根长平均减小9.9%和43.3%,导致根苗比表现为W1增温处理显著低于CK(P<0.05),W2增温处理和CK 差异不显著(P>0.05)。相对而言,生物量的分配在CK 和增温处理间差异较小,根冠比在增温处理和CK 间差异不显著(P>0.05)。
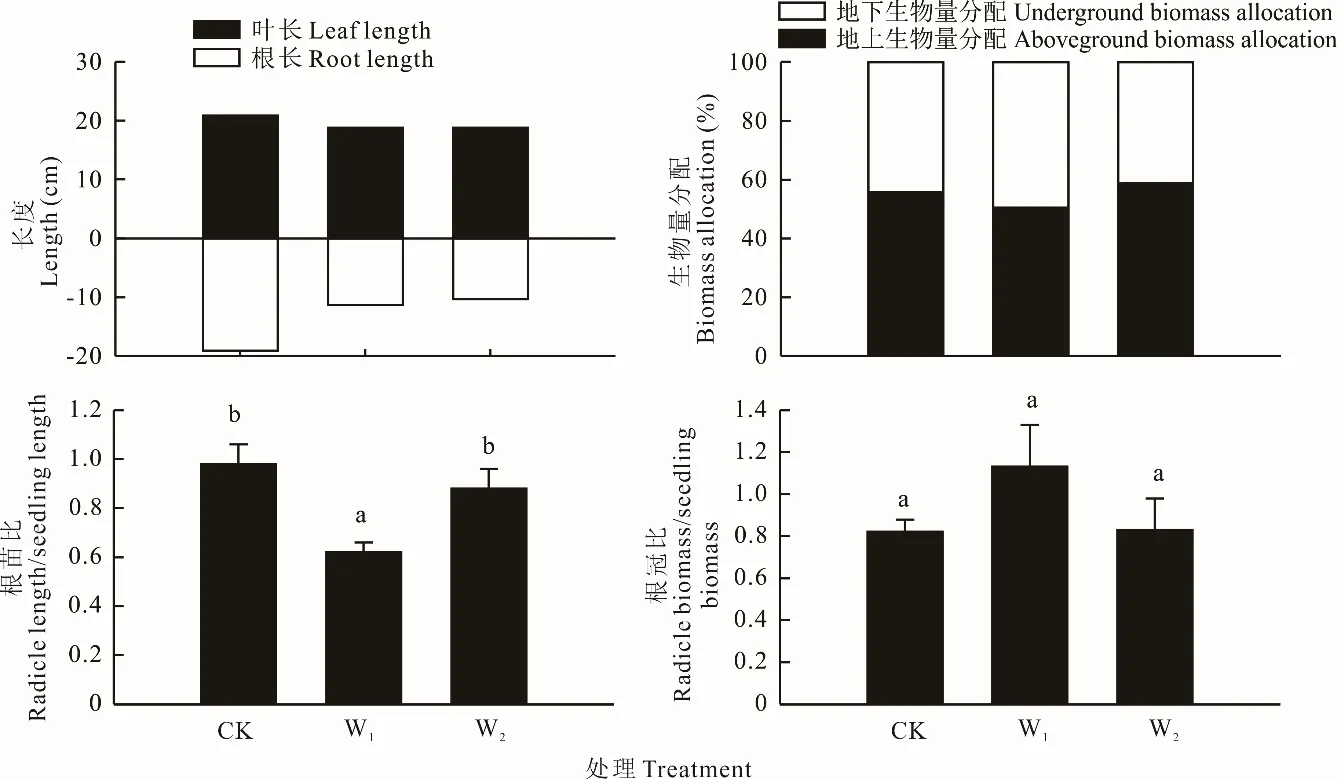
图4 野外模拟增温对幼苗生长特征的影响Fig.4 Effects of simulated warming on seedling growth characteristics in field
2.3 模拟增温对无芒雀麦繁殖生长特性的影响
当无芒雀麦生长至繁殖期时,个体生长受到增温处理的影响,对生殖生长和营养生长的分配表现出不同的适应性(表2)。结果表明,与CK 相比,W1处理下无芒雀麦植株高度增加8.91%;但其生殖生长比率却降低43.22%,根冠比降低41.67%。W2处理则对生殖生长影响较大,提高了无芒雀麦的生殖生长比率,对生殖投入和千粒重也起到一定的正向影响。
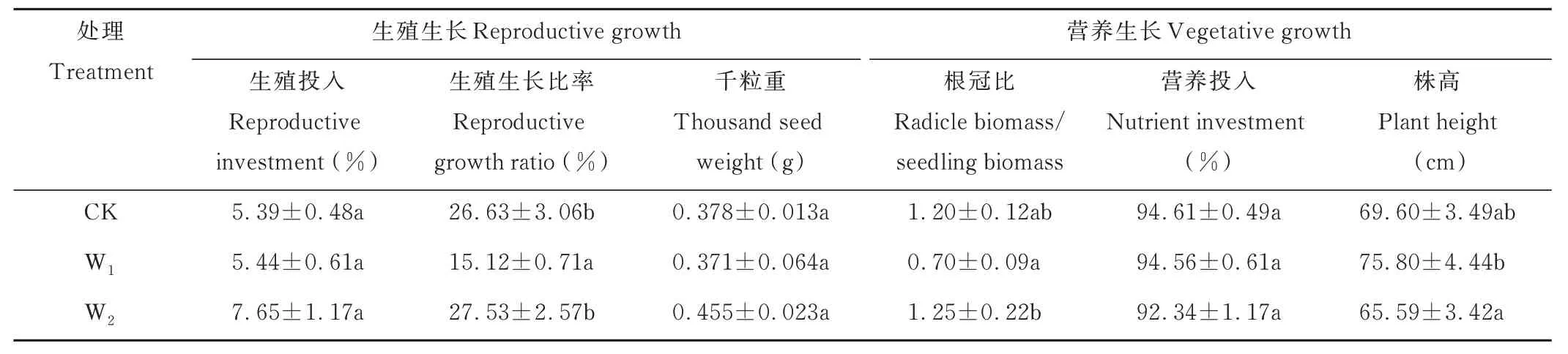
表2 繁殖生长指标在模拟增温处理下的差异Table 2 Differences of reproductive growth indexes under simulated warming treatment
2.4 无芒雀麦幼苗期和繁殖期地上、地下生物量拟合分析
为进一步探究无芒雀麦幼苗期和繁殖期地上生物量和地下生物量的相关关系,对二者进行回归分析(图5)。幼苗期的无芒雀麦在W1处理下,生长模式仍和CK 处理相同,均为异速生长,但W1处理下的斜率为2.21,小于CK 的斜率,说明地上生物量和地下生物量异速生长的关系减弱。W2增温处理时,两者呈线性关系,符合等速生长理论。从不同处理无芒雀麦幼苗期和繁殖期的表现来看,CK 处理下幼苗期地上生物量和地下生物量从显著(P<0.01)的异速生长转变为等速生长,在W1处理下两者的生长模式也遵循这一变化规律,但相关性明显减弱。在W2处理下,幼苗期和繁殖期的无芒雀麦均表现为极显著线性相关关系,但在繁殖期时,两者等速生长关系更为密切。
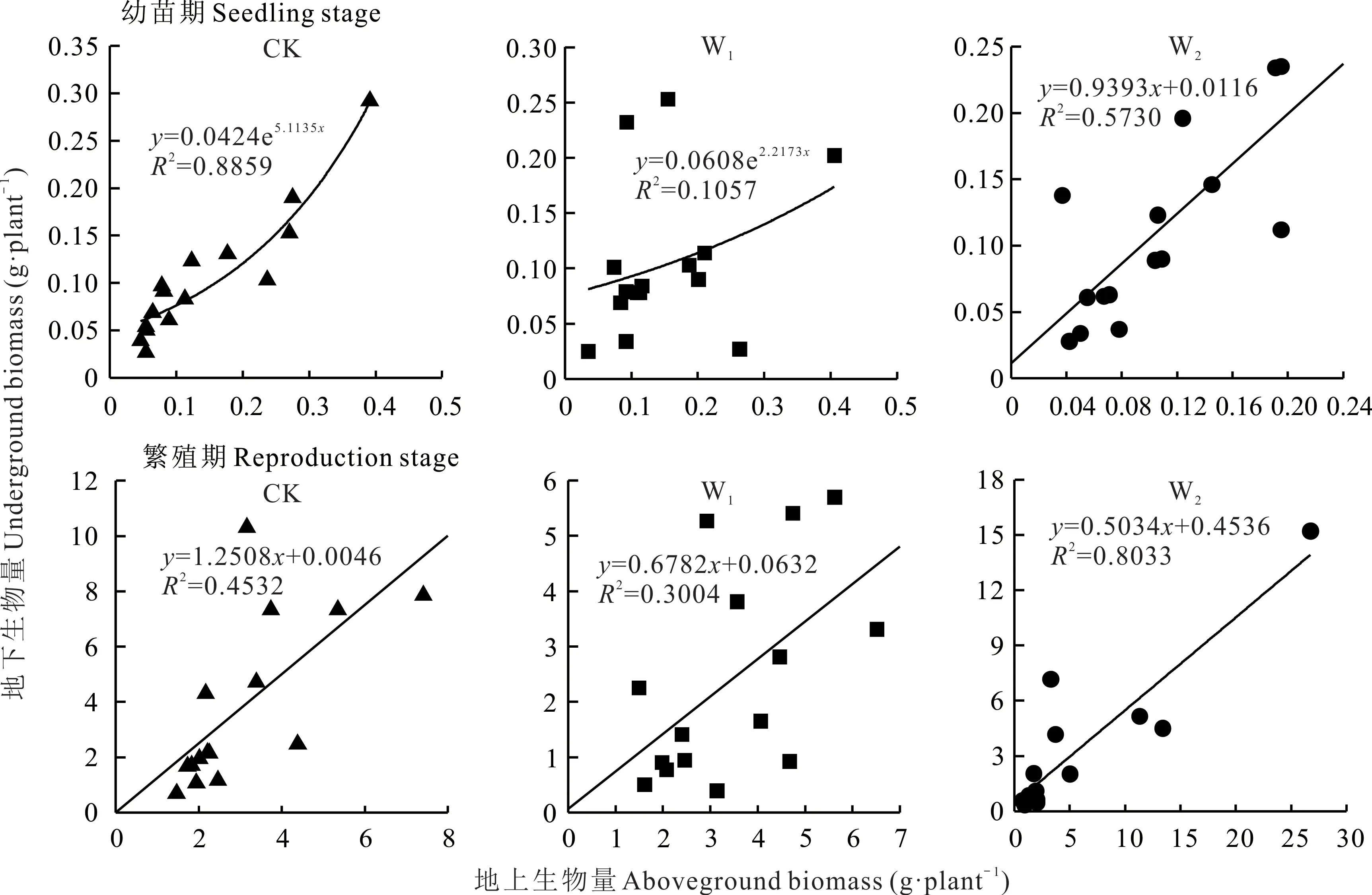
图5 幼苗期和繁殖期地上生物量与地下生物量的拟合Fig.5 Fitting of aboveground biomass and underground biomass at seedling and reproduction stages
2.5 无芒雀麦不同阶段生长特性在温度变化下的通径分析
无芒雀麦不同阶段应对环境增温的适应情况可以通过生长特征直接表现出来,利用通径分析明确生长特征指标之间相关影响的程度(图6)。在增温环境下无芒雀麦会降低发芽率,减少幼苗生物量的积累,生长至繁殖期则会抑制植株个体增高而提高分蘖能力,最终通过减少繁殖器官生物量提高营养器官生物量,减少地上生物量而提高地下生物量来规避增温胁迫的风险。
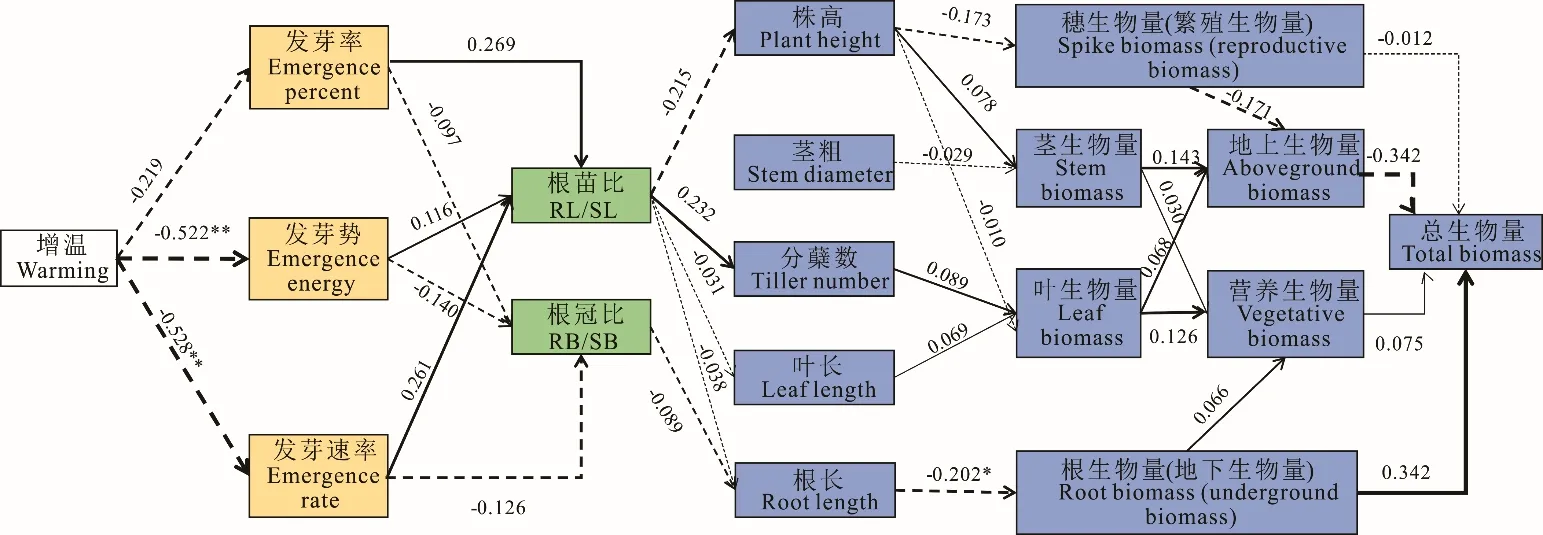
图6 增温影响无芒雀麦不同生长阶段生长指标的通径分析Fig.6 Path analysis of the influence of warming on growth indexes of B. inermis at different growth stages
3 讨论
3.1 种子发芽对增温的适应性
温度是决定种子萌发的重要条件之一,适宜温度为种子奠定良好的萌发环境,但在如今全球气候变化的背景下,温度的变化将会影响种子萌发的进程[20]。增温对种子出苗多为正向影响,即增温会提高种子的发芽率[21]。本研究中,无芒雀麦种子发芽率在W1增温处理下显著低于对照处理(CK),说明小幅度增温对无芒雀麦种子发芽并未起到促进作用,甚至在试验期间还抑制了种子发芽。前人研究证明,田间变温条件可以刺激野生菟丝子(Cuscuta chinensis)种子的萌发而提高发芽率[22],但有些研究中也发现如果增温幅度较小,对发芽率的影响可以忽略不计,甚至会对发芽率起到抑制作用,这也与种子质量以及种子对温度的耐受程度有关[9,23]。但是当增温幅度增大时,对种子发芽的促进作用还是显而易见的,本研究中W2增温处理有效提高发芽势和发芽速率。说明当积温持续增加至某一温度时,增温对种子发芽起到了促进作用,这也表明无芒雀麦种子对增温具有一定的适应调节能力,增温2.63 ℃还是在发芽适应的温度范围内,因此达到其原有的生长状态,并且表现出更高的水平。在适应的温度下快速萌发,能够在原生境中提前获取生长资源,提高该植物健康生长的几率[24]。
3.2 幼苗生长对增温的适应性
温度是影响植物生物量分配模式发生改变的重要因素之一,温度通过影响植物对水分、养分的吸收,进而间接影响植物的生长和生物量的累积[25]。本研究中,增温阻碍了幼苗叶长和根长的生长。在其他同类研究中,不同物种对增温的响应情况表现并不一致,矮嵩草在增温0.56~2.48 ℃范围内,叶片生物量随温度增加呈先减少后增加的趋势,黑褐苔草(Carex alrofusca)为负相关关系但影响不显著,垂穗披碱草和草地早熟禾(Poa pratensis)则与温度呈正相关关系[26],所以由于增温幅度、物种不同等原因会造成试验结果的差异;还有学者发现植物幼苗在适应温度范围内可以起到促进生长的作用,但超出耐受能力以外,幼苗的生长则会受到一定的抑制[27]。从本研究生物量分配上来看,增温对其的影响不显著,并没有通过调节生物量来促进幼苗的生长,这也说明无芒雀麦幼苗能在W1、W2温度下较为稳定地存活。根苗比、根冠比等是植物通过适应、调节后所表现出的综合指标,本研究结果显示在W1增温下根苗比显著低于对照组,说明在这一温度下根系的生长受到抑制,无法增加长度获取更多的水分和养分,即使能获得略多的生物量分配,也无法实现地下部分的良好生长。
3.3 繁殖生长对增温的适应性
OTC 装置通过改变增温范围内的水热等微气候状况,对植物的物候、群落结构、生物量等的生态学反应奠定基础[28],研究者通过OTC 装置探究植物群落结构和植物生长,增温能有效促进植物能量的积累[29-30],但如果温度增加超过最适温度后,反而起到抑制作用[31]。随着植物响应气候变化的研究逐渐被重视,观察植物形态结构在环境中的变化成为主要研究内容,对将来气候变化的适应需要许多不同的性状同时进化,因此这些性状之间存在着相关性,相互牵制。
Berkowitz[32]发现,植株高度和光竞争能力之间显著正相关。本研究中小幅度增温(W1)使无芒雀麦株高增加,利于光竞争能力的提高,同时促进叶片等营养器官的生长,以此输出更多的光合产物,前人研究也表明温度的增加能延长植物生长季节,从而提高叶片在生长过程中的功能作用[33]。禾本科功能群植物具有从生理上调控资源分配模式的能力,其叶片较大并分布于不同的高度层,所以在增温条件下,有利于其光合速率增加和分蘖能力、种子繁殖能力增强[34]。
Reekie[35]研究认为植物株高的增加将会牺牲生殖分配,说明在这两者间的权衡也是植物应对环境变化的策略之一。本研究中,大幅度增温(W2)使无芒雀麦株高降低,提高生殖生长的投入,增加了生殖生物量。这是植物个体随生殖生物量增加表现出的最直接的代价,增大生殖分配投入的同时意味着植物需要减少对个体大小结构的分配。以上表现都说明模拟增温提高了无芒雀麦的同化产物投入到生殖生长中的比例,这有利于实现植物的繁殖更新。
植物从营养生长转变为生殖生长主要依靠温度的控制[36],植物各组分生物量的分配随着生长阶段的变化,也会提供适时的分配策略,本研究中增温能提高无芒雀麦的生物量积累,营养生长的减少使投入到生殖生长的资源增加。有研究表明,气候变暖会促进草地生态系统地下生物量的增加[37],这可能是因为植物碳水化合物生物量的积累速度比消耗速度快[38]。也有研究表明,夏季变暖显著降低了羊草(Leymus chinensis)的地下部分生物量,增加其地上生物量[39]。本研究中大幅度增温(W2)相较于小幅度增温(W1)来说,无芒雀麦将更多的资源偏向于投入到地下部分,首先考虑根部的生长,保证植物的存活,这符合植物在生存和繁殖二者间两面下注的理论[40]。
3.4 植物适应性生长对增温的响应
本研究中幼苗期的无芒雀麦受到增温影响时,地上和地下生物量的生长关系从异速生长转变为等速生长。异速生长被认为是一些物种自然选择的结果[41],但在温度升高环境背景下,幼苗逐渐将生物量更多集中在地上部分,是为了在生长初期获取更多的光资源,产出更多的光合产物,再利用有限的资源优先分配给地上[42],随后在增温幅度更大的情况下,地上和地下生物量的关系更加密切,说明在增温过程中无芒雀麦幼苗发挥了自身的调节机制来适应环境变化。和繁殖期无芒雀麦相比,幼苗期这种关系的转变可以确定这一时期更易受到增温的影响。本研究中繁殖期的无芒雀麦在增温处理下地上和地下生物量之间的关系,一直保持着等速生长关系,并且随着增温幅度的增大拟合程度更高,说明无芒雀麦对适应大幅度增温具备更良好的应对能力。
植物的生活史都要经历生长和繁殖的过程,而在不同生长发育阶段由于各器官所承担任务的能力不同,导致植物不得不在各阶段对资源的分配做出倾向性安排[43-44]。本研究通过2 个模拟增温处理发现,增温首先抑制了种子的发芽,在幼苗生长期间会通过增强根苗比来帮助无芒雀麦度过脆弱的幼苗期,而在营养生长和繁殖生长的协调中,无芒雀麦会更倾向于提高营养生物量的积累来维持生长。这种权衡方式可以保证野生无芒雀麦在群落中占据优势地位,从而获取更多的生长资源和生产空间[41]。但有些研究却发现在生活史后期,植物会更愿意将资源投入到繁殖生长中以确保延续后代[45]。无芒雀麦具备有性繁殖和无性繁殖两种繁殖方式,相比之下植物会更倾向于将资源分配给更需要的部分,或者是投入到更有把握的方式中[46]。
4 结论
模拟增温达到W2增温幅度才会提升无芒雀麦种子的发芽能力,加快萌发进程;但幼苗地上部分和地下部分的生长都受到增温的抑制影响;繁殖生长过程中受增温影响将更多的资源投入到生殖生长,同时加强根系的生长以保证地下部分生物量的积累。最终通过减少繁殖器官生物量来提高营养器官生物量,减少地上生物量而提高地下生物量来规避增温胁迫的风险,以此来实现植物在增温环境下存活和繁殖的协调。
猜你喜欢
Rice Science(2021年4期)2021-07-13 10:00:54
中国生殖健康(2020年8期)2021-01-18 03:05:08
中国生殖健康(2020年2期)2021-01-18 02:51:10
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
中国生殖健康(2018年1期)2018-11-06 07:14:48
中国生殖健康(2018年2期)2018-11-06 07:10:32
种子科技(2018年11期)2018-09-10 00:56:48
许昌学院学报(2018年8期)2018-09-05 02:05:18
第二课堂(小学版)(2018年6期)2018-07-09 01:06:46
快乐语文(2016年10期)2016-11-07 09:44:51
