
温湿度胁迫对两种榕属植物种子萌发和幼苗生长的影响
2023-09-24 07:27:08官云芳彭艳琼
种子 2023年7期
官云芳, 彭艳琼, 古 旭, 张 颖, 张 媛
(1.西南林业大学生物多样性保护学院, 昆明 650224;2.中国科学院西双版纳热带植物园, 云南 西双版纳 666303)
气候变化引起的高温、干旱、土地盐碱化等极端气候事件越来越频繁,显著地影响植物的生长和更新,耐多种逆境交互胁迫的植物群落在生态恢复中发挥着重要的作用[1]。种子萌发是植物生命周期的关键过程,是幼苗生长、植物群落建成与延续成功的基础,也是非生物胁迫最敏感的阶段[2]。温度与干旱是这一时期主要的影响因子。高温可通过改变种子的膜透性、酶促反应等过程影响种子萌发,进而影响植物幼苗增补和种群更新[3-4],故高温是影响林木生长发育的重要逆境因子之一,尤其对林木幼苗生长影响极大,幼苗能否忍耐高温逆境成苗直接关系到林木育苗的成败。湿度胁迫会使种子发芽减少或延迟,叶片气孔关闭,阻碍酶合成反应或终止光合作用,甚至导致植物死亡[5-6]。在全球变化的大背景下[7],高温往往伴随着干旱同时发生,已有的研究主要考虑高温单一因子的影响[8-9],而对温湿度交互影响的研究较为缺乏。探索种子萌发、幼苗生长对外界多种环境因子交互胁迫的响应,对于物种保护以及评估植物群落和生态系统的稳定性至关重要[10]。
桑科(Moraceae)榕属(Ficus)植物是国际上公认的热带雨林中的一类关键植物类群[11]。在热带雨林中,一年四季均可挂果的榕属植物是鸟类、兽类、昆虫等动物重要的食物来源,同时也为多种寄生和附生植物提供了良好的栖息与生存空间,使得榕树在维持热带雨林生态系统稳定性中发挥着不可替代的作用[12-13]。此外,一些榕属植物可在受到破坏的林段、林窗等区域萌发、生长,成为先锋种;一些榕属植物可通过在寄主植物上萌发生长,产生“绞杀”现象并实现林分的更新。因此,榕树种子萌发和幼苗生长定居的过程在热带森林演替的动态过程中具有重要作用[14]。此外,榕属植物也是重要的园林绿化树种和食药用植物,具有较高的应用价值[11,13]。近年来,榕属植物的生理生态特征以及经济利用价值越来越受到关注[15]。郭健华等[16]研究表明,榕树具有一定的耐热性,但极端天气的频繁出现对榕属植物的生长和存活带来了较大影响。同时,一些城市种植的行道榕树,在经历了较为极端的气候波动后,出现了植株生长受到损害甚至死亡的现象,造成了城市景观的破坏和较大的经济损失[17-18]。目前,尚未见关于榕属植物对温湿度胁迫及其交互作用响应的报道。在全球环境变化大背景下,揭示榕属植物对极端环境因子响应的研究具有重要的现实意义。
斜叶榕(FicustinctoriaCorner.)和聚果榕(FicusracemosaL.)是热带雨林中的两种常见树种,同时也是常用的园林绿化树种。本研究将通过设置不同的温度和干旱条件,研究温度和湿度对斜叶榕和聚果榕种子萌发和幼苗生长的影响。通过探索两种榕属植物在温度、干旱交互胁迫下种子发芽和幼苗生长特性及耐受程度,旨在揭示两种榕树对逆境胁迫的耐受力,以期为榕属植物的保护和园林绿化树种的选择提供参考。
1 材料与方法
1.1 试验材料
试验所用斜叶榕和聚果榕种子采自中国科学院西双版纳热带植物园(21°41′N,101°25′E)及周边地区,每种选取3株树,每株树采集10个成熟榕果,将种子取出后清洗干净并阴干,从中随机挑出均匀、饱满、健康的种子备用。
1.2 试验方法
1.2.1温度对种子萌发的影响
人工气候箱(赛福 PGX-350C)设置20 ℃、25 ℃、30 ℃、35 ℃共4个温度梯度。试验采用纸皿法:将培养皿(直径12 cm)用高压蒸汽灭菌器(MLS-3751L-PC)在121 ℃下灭菌15 min,灭菌好的培养皿内垫上一薄层脱脂棉,脱脂棉上铺上消毒过的滤纸,以超纯水浸润脱脂棉和滤纸。每个处理3个重复,每个培养皿均匀放置15粒种子,每粒种子独立编号以便于跟踪观察。相对湿度设置为80%,光照条件设置为每隔12 h光暗交替1次。试验过程中每天对培养箱进行检查并及时补充蒸馏水,保持滤纸湿润、清洁,并记录种子发芽数,若发现霉变种子及时清除。
1.2.2干旱胁迫对种子萌发的影响
以PEG-4000来模拟干旱条件,设置5个浓度分别为0(对照组),5%,10%,20%,25%,对两种榕属种子分别浸泡24 h后放置于培养皿,每个处理3个重复,每皿均匀放置15粒种子,每天记录种子发芽数,人工气候箱内相对湿度和光照等设置同1.2.1。
1.2.3温度和干旱胁迫对种子萌发的交互作用
上述每个温度梯度均设置5个PEG浓度(0,5%,10%,20%,25%),共20个组合,每个组合3次重复,每种榕果测量60个培养皿,每皿15粒种子,共计900粒种子,其他方法同1.2.1。
1.3 指标测定及数据处理
1.3.1指标测定
试验以胚根露出种皮2 mm作为种子萌发的标准,每天观察种子萌发数直至所有种子萌发结束,发芽最高峰时间为当日种子萌发数最多的时间。每周使用电子游标卡尺(宝工PD-151)测定每株幼苗的胚根和胚芽长,若一周内胚根和胚芽长不再生长视为生长结束。实验结束后计算发芽率、发芽势、发芽指数。
发芽率/%=(发芽种子数/供试种子总数)×100%;
发芽势/%=(第9天发芽种子数/供试种子总数)×100%;
发芽指数=∑(Gt/Dt),
式中:Gt为在时间t日的发芽数,Dt为发芽日数。
1.3.2响应面分析试验
以温度胁迫(T)、PEG胁迫(P)、萌发时间(C)为变量,根据 Box-Behnken 试验设计原理[19],设计3因素3水平的中心组合,以种子发芽率为响应值,并采用Design-Expert13.0.1 Trial 软件进行设计与分析。
1.4 数据处理与分析
运用Microsoft Excel2010软件整理数据,IBM SPSS Statistics26.0软件进行多因素方差分析,分析温度和PEG溶液模拟干旱胁迫及其交互作用对种子萌发指标和幼苗生长参数的影响,根据Duncan检验分析不同处理间的差异显著性,运用OriginPro2019软件和Design Expert13.0软件作图。所有数据用平均值±标准误表示。
2 结果与分析
2.1 温度与干旱胁迫对两种榕属植物种子生长进程的影响
低温显著延长了榕树种子萌发进程,高温与PEG溶液浓度形成的共同胁迫可以减缓种子萌发速度,降低种子萌发率。从图1(A)可知,25 ℃和30 ℃温度水平下,斜叶榕种子在第7天开始萌发,且最终萌发率可达90%以上,说明适宜温度下斜叶榕种子对干旱有一定抗性,高低温则显著延长了种子萌发时间,在35 ℃无干旱胁迫下,萌发率可达80%,但同样温度加上干旱胁迫后,萌发率降至20%~60%;与斜叶榕类似,由图1(B)可知,聚果榕在第7天开始萌发,但聚果榕种子发芽进程与斜叶榕存在一定差异,主要表现在35 ℃下的萌发进程较斜叶榕快,但低温状态下萌发进程较斜叶榕慢,且PEG溶液浓度对种子萌发进程的影响在低温状态下更显著。

注:T1、T2、T3、T4分别表示4 个温度水平(20 ℃、25 ℃、30 ℃、35 ℃);P1、P2、P3、P4、P5分别表示5 个PEG溶液浓度(0,5%,10%,20%,25%)。图1 温度、PEG胁迫下两种榕属植物种子萌发的进程Fig.1 Seed germination progress of 2 Ficus plants under temperature and PEG stress
2.2 温度与干旱胁迫对两种榕属植物种子发芽率、发芽势和发芽指数的影响
如图2所示,在25 ℃和30 ℃温度梯度下,不同干旱胁迫下斜叶榕种子都具有较高的发芽率(97.30%和99.56%),且保持较为一致的发芽速度。在低温(20 ℃)和高温(35 ℃)下,发芽率则受到影响,分别降低至61.33%和47.61%。聚果榕种子在4 种温度处理下的平均发芽率达70%以上,在30 ℃时萌发率最高,为88.89%,而在25 ℃时发芽整齐度最高,发芽势为63.37%,发芽速率也最快。在适宜温度下,榕属植物种子的发芽率、发芽势、发芽指数均较高,说明此时斜叶榕和聚果榕种子不但具有较高的发芽率和种子活力,同时也有更高的萌发整齐度。同时,比较两种榕属植物在25℃和30℃条件下,种子的发芽指标在不同PEG溶液浓度下变化幅度较小,而在高温(35 ℃)和低温(20 ℃)状态下变化较大,说明在极端温度条件下,种子对干旱更为敏感,两种榕属种子均呈现相似规律。

注:误差线表示标准偏差。不同小、大写字母分别表示处理间差异显著(p<0.05)。图2 温度、PEG胁迫下两种榕属植物种子的发芽率、发芽势、发芽指数Fig.2 Seed germination rate, germination potential and germination index of 2 Ficus plants under temperature and PEG stress
2.3 温度与干旱胁迫对两种榕属植物幼苗生长的影响
除10% PEG处理时聚果榕的胚芽长在各温度水平差异不显著外,其他各PEG处理条件下,两种榕属植物的胚芽和胚根长在4种温度水平下均有显著差异(表1)。且随着温度的升高,幼苗生长均呈现先促进后抑制的趋势。斜叶榕在所有温度梯度下,PEG处理对胚芽长的影响显著。而聚果榕在除35 ℃外的其他温度梯度下,PEG处理对其胚芽长的影响均显著。斜叶榕在除30℃外的其他温度梯度下,PEG处理对其胚根长的影响均显著,而聚果榕在所有温度梯度下,PEG处理对其胚根长的影响均显著。同时,斜叶榕幼苗的生长速度整体趋势上比聚果榕快,尤其是在25 ℃和30 ℃下斜叶榕胚根获得最适生长。
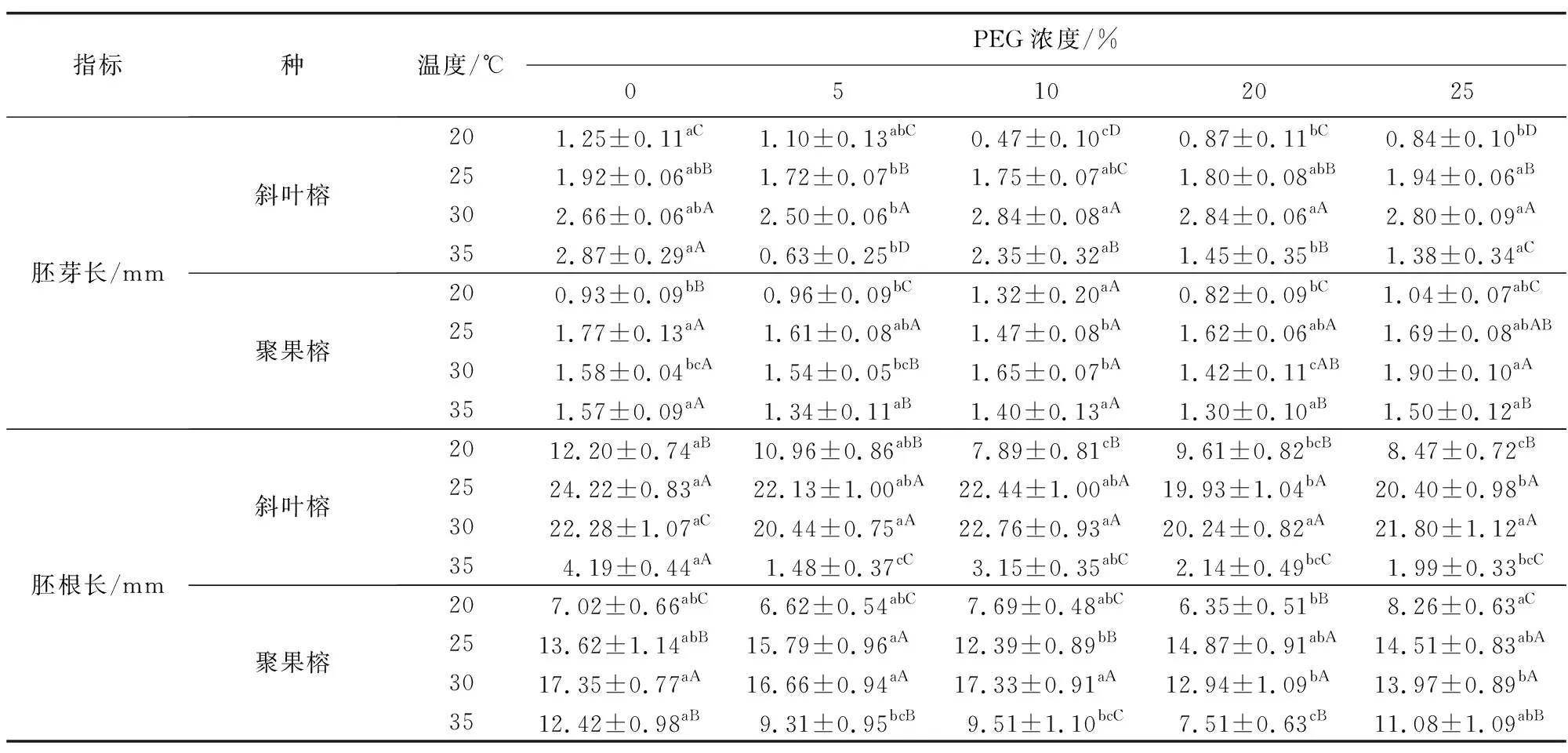
表1 温度、PEG胁迫下两种榕属植物幼苗的胚根和胚芽长Table 1 Root and shoot lengths of 2 Ficus plant seedlings under temperature and PEG stress
2.4 温度与干旱胁迫对两种榕属植物种子萌发和幼苗生长的交互作用分析
由表2可知,当温度单独作用时,对两种榕属种子萌发及幼苗生长的所有指标有极显著影响,当模拟干旱单独作用时,对斜叶榕所有指标均有显著影响,而对聚果榕的发芽势、胚根和胚芽影响显著,对发芽率和发芽指数影响不显著。温度、干旱共同作用时,对斜叶榕的发芽率、胚根长和胚芽长具有交互作用,对聚果榕的胚根长和胚芽长表现出交互作用。
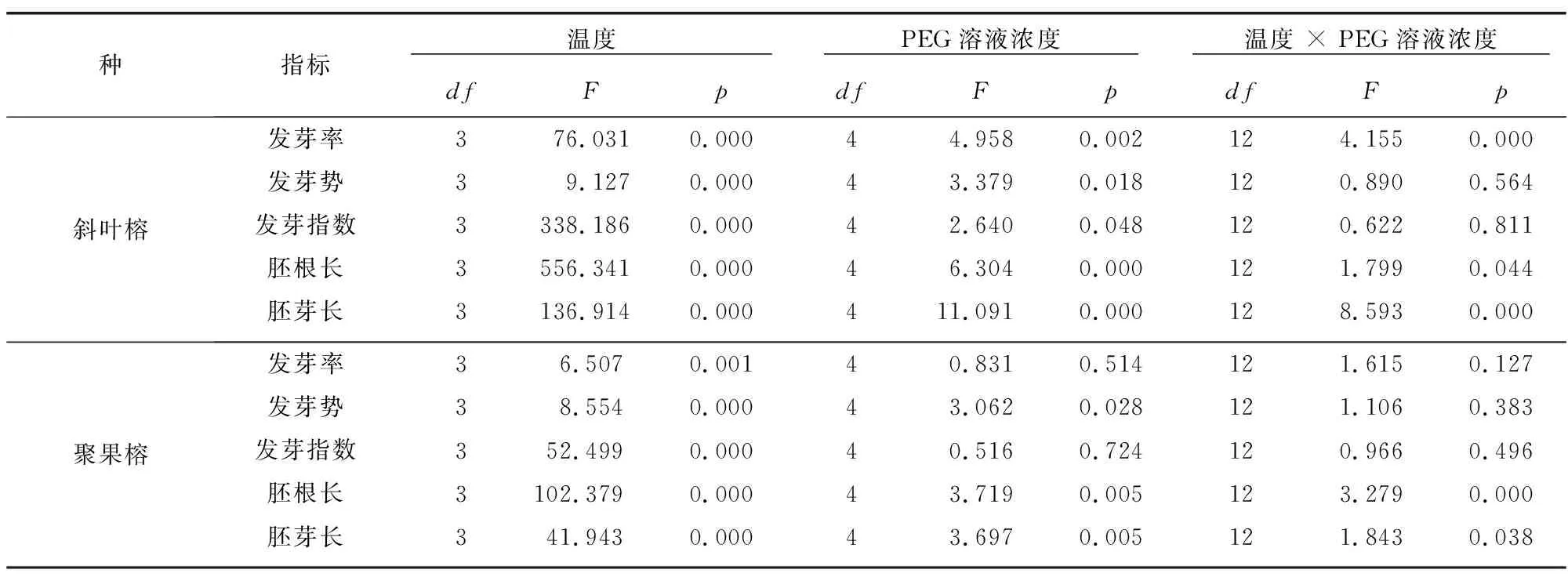
表2 温湿度及其交互作用对两种榕属植物种子发芽率、发芽势、发芽指数、胚根长和胚芽长的影响Table 2 Effects of temperature,drought and its interaction on germination rate,germination potential,germination index,root length and shoot length in two Ficus plants
2.5 温度、干旱交互胁迫对两种榕属植物种子萌发率的拟合分析
斜叶榕种子发芽率的回归方程:Y1=98.67-34.03A+2.89B+6.54C+4.85AB+0.86AC-2.22BC-38.92A2+3.55B2-8.89C2;聚果榕种子发芽率的回归方程:Y2=92.06-4.17A-1.67B+0.83C+1.67AC+1.11BC-6.8 7A2-0.75B2-5.76C2。由表3可知,斜叶榕试验模型达到极显著水平,模型决定系数R2=0.995 3,说明该模型的拟合度很好,失拟项差异不显著;聚果榕试验模型达到显著水平,模型决定系数R2=0.826 9,说明该模型的拟合度较好,失拟项差异不显著,综上所述,两个模型均可用于斜叶榕、聚果榕发芽率的条件分析和预测。
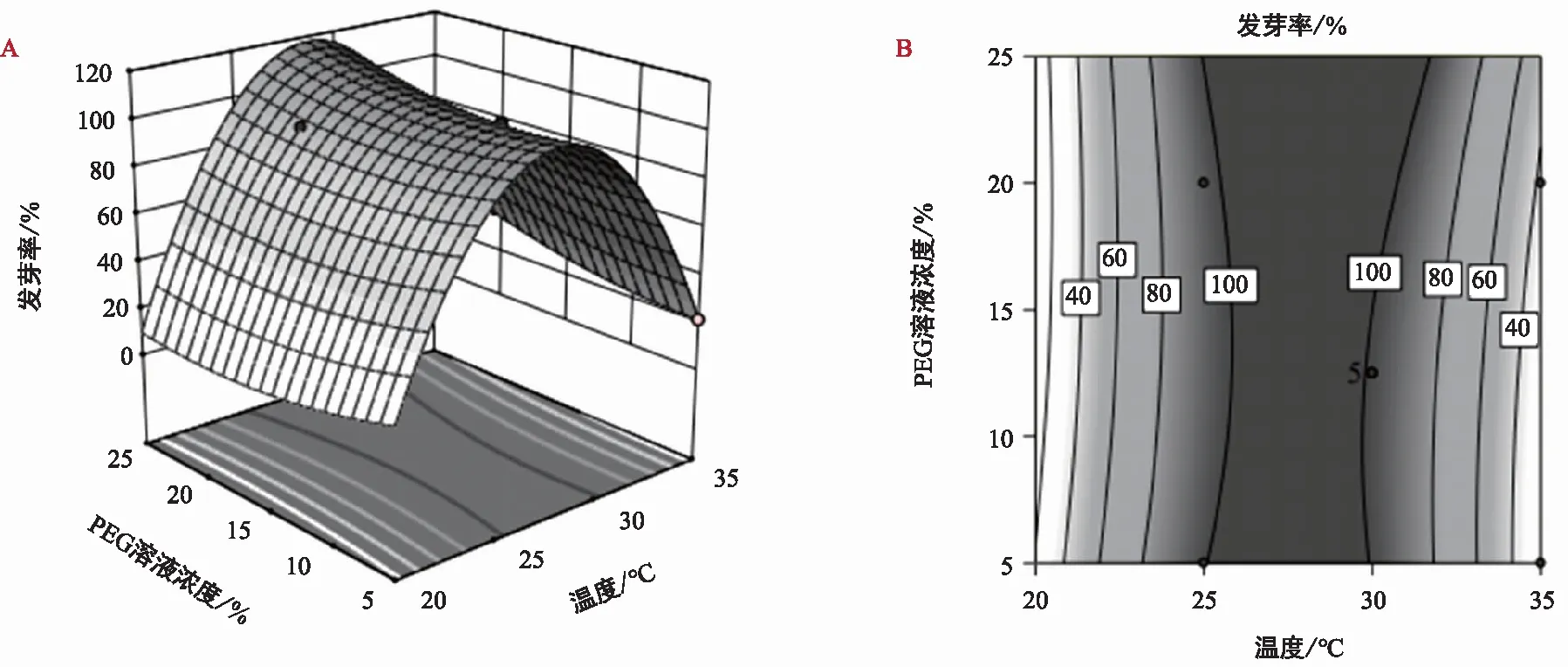
图3 温度和PEG交互作用对斜叶榕种子发芽率的影响 Fig.3 Effect of temperature and PEG interaction on seed germination rate of Ficus tinctoria Corner.
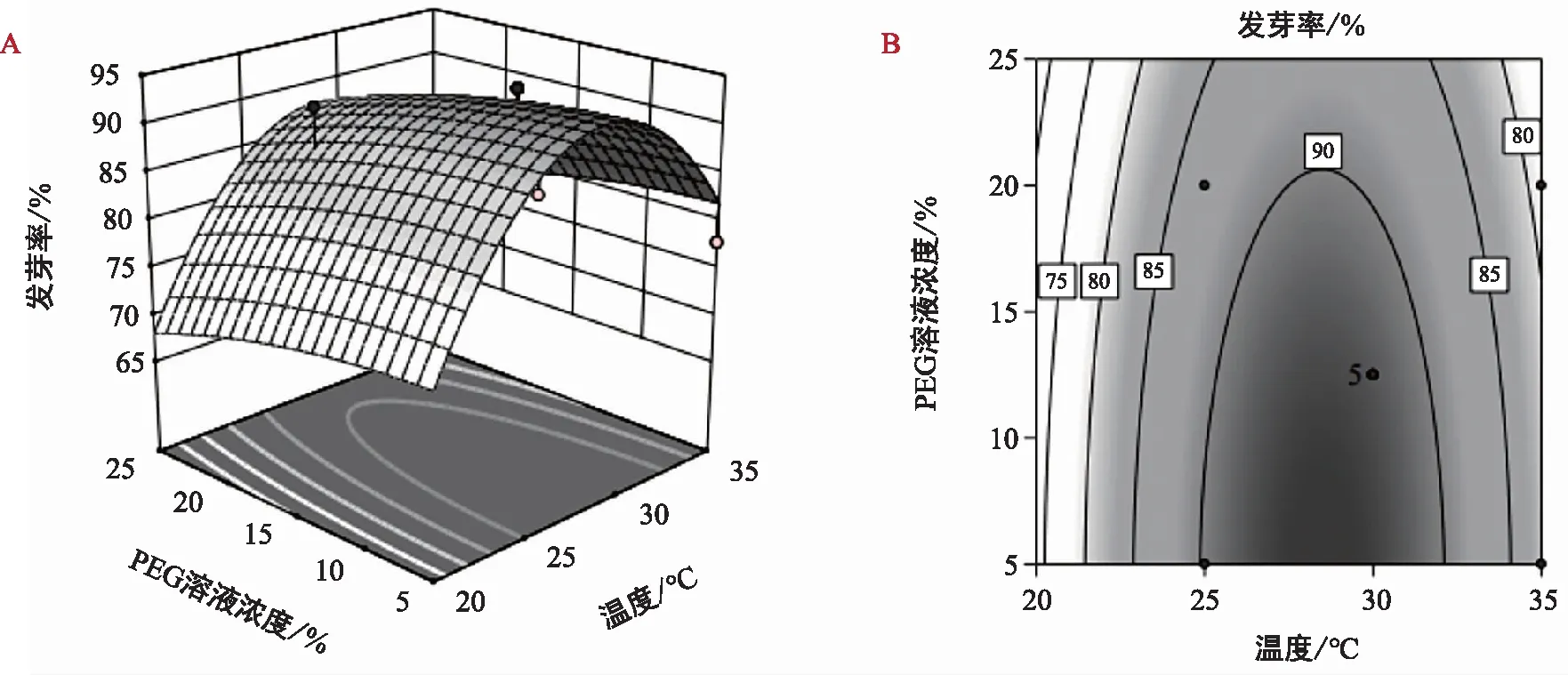
图4 温度和PEG交互作用对聚果榕种子发芽率的影响 Fig.4 Effect of temperature and PEG interaction on seed germination rate of Ficus racemosa L.

表3 回归模型方差分析Table 3 Regression model analysis of variance
通过比较两种榕属植物种子萌发的响应面曲线,模拟结果如图3和图4所示。密集或陡峭程度越高表明对结果响应值的影响越大,温度和PEG溶液浓度在斜叶榕种子萌发上体现出比聚果榕显著的交互作用,这与方差分析结果一致。
3 讨 论
种子萌发是植物生活史的重要阶段,易受自身和外界胁迫环境的影响,探索种子萌发、幼苗生长应对温度、干旱及其交互胁迫下的响应策略,可以更好地了解植物生存适应和种群建植的过程[4,20]。研究表明,在不同环境和外界胁迫条件下,不同植物种类生长发育的能力也不同[21-22]。本研究结果表明,种子萌发状态受温度影响的程度因物种而异。25~30 ℃更有利于斜叶榕和聚果榕的种子萌发及幼苗生长。在20 ℃和35 ℃时,斜叶榕种子萌发和幼苗生长受到较大影响,而与斜叶榕相比,聚果榕种子对高低温表现出更强的抗性,在本试验设置的温度范围内,聚果榕种子发芽率变化较斜叶榕更小,说明不同种榕树对高低温的耐受性存在差异。徐迪等[23],洪岚等[24]研究表明,一些植物的种子,如三叶鬼针草、沙棘种子的发芽率对温度的响应并不明显。一般而言,温度对种子萌发的影响是一个持续积累的过程[25],低温沉积则有助于打破种子休眠[26]。本研究中,高低温处理下两种榕属种子的最终萌发率会稍有降低,尤其是低温下种子萌发进程被显著延缓,斜叶榕种子的萌发高峰期也稍晚于聚果榕种子,但在适宜的温度条件下,两种榕属种子都可以大量萌发且萌发进程较快,这表明低温下榕属植物种子可能具有一定的休眠时间,种子完全解除休眠可能需要更高的积温条件。
水分是影响种子萌发、幼苗形态建成的主要环境因子,是种子吸胀后胚轴伸长的起始因子,能够参与发芽代谢过程[27]。许多植物长期生活在干旱生境中,植物体内因水分缺失导致细胞内所需水分含量不足,危害严重的可使种子无法正常萌发或抑制幼苗生长[28]。目前气候变化加剧了干旱的严重性和持续时间,热带雨林也同样受到干旱等极端气候的影响[29]。有关研究显示,西双版纳热带雨林种子和幼苗在6个月的旱季中,林下树种幼苗的死亡比例能达26.49%[30],可见旱季是热带雨林林下幼苗存活的一个关键时期。本研究结果显示,除10% PEG处理外,两种榕属植物的胚芽和胚根长在其他PEG浓度处理下均有显著差异,表明幼苗生长对PEG胁迫较为敏感,这与闫兴富等[2]的研究结果一致。一般情况下,雨季会出现一个较短的干旱持续时间,植物对低于正常水平的短期干旱反应迅速,能够利用部分降水维持其生长[29],这一短暂的关键期不仅有助于种子从深层土壤中充分吸收水分尽早萌发,而且也能减少因降雨强度过大对幼苗根系造成的伤害,在一定程度上降低了幼苗在雨季的死亡率。因此,在生产实践中,利用雨季中短暂的干旱期播种榕属种子可能比雾凉季、干热季播种时更容易缓解干旱胁迫对种子造成的伤害。
植物在自然界常常要面临多种环境因子的共同胁迫,其中干旱与温度因子的共同作用可能会加剧对种子发芽和幼苗形态建成的影响[4]。本研究表明,温湿度交互胁迫,对斜叶榕的影响较聚果榕更大,说明在极端温度条件下,斜叶榕对干旱的抵抗能力下降,而聚果榕表现出了更强的温度和干旱的适应性。两种榕属种子的发芽率、发芽势、发芽指数在20 ℃与PEG交互作用时均有显著差异,说明低温下种子萌发对水分有更高的要求,其原因可能是在西双版纳热带森林中,极易在低温下形成浓雾,从而降低了干旱胁迫,在一定程度上缓解了对种子可能造成的冻害程度。
植物性状是生态过程的体现,它们通过调整幼苗获取资源的结构达到最适合的条件,从而提高资源的吸收[30]。本研究结果显示,斜叶榕的胚根生长速度明显比聚果榕快,且对干旱的响应更加明显,表明斜叶榕分配给胚根生长的能量更多。本结果也显示,适宜的温度条件下,当给植物施与轻度干旱时,植物的胚根生长更强,极端高温下则随着干旱胁迫程度的增加胚根生长受到抑制,这可能是因为斜叶榕的生长环境及种子特性造成的。斜叶榕主要分布于低谷山地林中或旷地、水旁,在瘠薄的土壤或石缝中均可生长,根系发达,也是热带雨林中常见的半附生榕树,其种子常常在棕榈树等的缝隙中萌发形成“绞杀”现象[14],斜叶榕为了保持自身的生长,会将根系不断扩展到土壤或附生植物深处以获得水分。与此相反,聚果榕自身为阳性植物,幼树在光照条件充足地段生长很快,这可能也是聚果榕胚芽得以在中高温下保持较高生长能力的原因。两种榕属植物对温湿度交互作用的响应表现出差异,从植物生理方面对不同榕属植物响应的机制进行研究将是一个有意义的研究方向。
本研究结果暗示,在全球变化大背景下,热带雨林中榕属植物的种子萌发和幼苗形态建成可能会受到较大的影响,但影响程度在不同的榕树之间可能存在差异,有必要在更大范围内开展榕属植物与气候变化关系的研究,从而为更大尺度上开展热带雨林生态系统稳定性的研究提供科学依据。同时,在选择榕属植物作为行道树种进行繁殖栽培时,应根据不同榕树的环境耐受性,结合种植地区的气候特征,进行科学合理的选择,避免因环境因子的不适宜而影响榕树的生长发育,甚至导致行道树的死亡[18]。
猜你喜欢
广西林业科学(2021年6期)2022-01-21 07:39:28
今日农业(2021年10期)2021-11-27 09:45:24
山西农业科学(2021年8期)2021-08-17 06:20:10
山西农业科学(2020年1期)2020-02-27 17:00:54
种子科技(2018年11期)2018-09-10 00:56:48
许昌学院学报(2018年8期)2018-09-05 02:05:18
天津农林科技(2015年1期)2015-12-30 13:05:14
中国火炬(2015年12期)2015-07-31 17:38:35
中国火炬(2013年3期)2013-07-24 14:15:06
中国火炬(2011年8期)2011-07-25 10:44:25
