
蝴蝶兰Phal.‘B’和Phalaenopsis.SogoYukidian‘V3’正反交后代花部性状分离表现
2023-09-06 11:15:44杨书才王世尧蒋拴丽赵玉安王瑞华杨录军
安徽农业科学 2023年16期
张 果,杨书才,王世尧,王 俊,蒋拴丽,冯 建,赵玉安,王瑞华,杨录军
(郑州市农林科学研究所,河南郑州 450005)
蝴蝶兰为兰科(Orchidaceae)蝴蝶兰属植物,花形似蝶,花色绚丽,赏花期长,被誉为“兰花皇后”,是国际上销量最高的兰科花卉之一[1]。杂交育种是选育蝴蝶兰新品种最常见且重要的一种手段,截至2021年3月在英国皇家园艺学会(RHS)上登录的兰科蝴蝶兰属杂交种达37 211个[2],由于国内蝴蝶兰育种工作起步较晚,且种质资源多依赖于引进,自育品种较少,与我国蝴蝶兰市场规模极不匹配[3]。随着杂交育种工作的开展,国内蝴蝶兰新品种培育也步入正途,关于蝴蝶兰观赏性状分离表现研究也随之增多[4-6]。研究表明蝴蝶兰杂交F1代出现亲本没有的花部纹案,存在较深花斑色覆盖遮挡住较浅花底色的表现,推测花部纹案受多基因调控结果,可能与易变基因的体细胞突变或转座子的位置效应有关[7-9]。
在蝴蝶兰杂交育种过程中,花色、花斑、唇部性状等观赏性状是衡量一个新品种优良的重要指标,探究花被片底色、花斑类别、唇部肉突和中裂片颜色等花部性状遗传表现,有助于更好地选择适宜的亲本,提高目标性状的育种效率。目前关于蝴蝶兰唇部肉突和中裂片颜色的研究尚未见报道,因此,笔者以Phal.‘B’和PhalaenopsisSogoYukidian‘V3’及其正反交后代(F1)为材料,探讨蝴蝶兰正反交F1代花色和唇部性状的遗传规律,以期为杂交育种选育亲本和目标性状定向育种提供理论依据。
1 材料与方法
1.1 试验材料供试亲本材料为Phal.‘B’和PhalaenopsisSogoYukidian ‘V3’,试验亲本及其杂交后代均栽培于郑州市农林科学研究所新品种试验中心连栋温室内,植株长势良好。
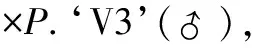
1.3 数据分析采用Microsoft Office Excel 2003、SPSS Statistics 26对试验数据进行统计分析。
2 结果与分析
2.1 亲本花部性状表现P.‘B’为白色花,花心有紫红色斑块,且该斑块有粗糙颗粒触感;P.‘V3’为大花型白色花,具体的花部性状见表1、图1A。

表1 亲本性状

注:A.亲本及杂交F1代花色;B.F1代肉突颜色;C.F1代中裂片颜色。Note:A.Parental and hybrid F1 generation flower color;B.F1 generation callus color;C.F1 generation mid-lobe color.
2.2 正反交F1代花部性状的分离表现
2.2.1正反交F1代花色花斑分离表现。根据花色、花斑的性状特征,P.‘B’ 和P.‘V3’的正反交后代(F1)均表现出一致的性状分离规律,故将其后代分为5个组群,编号为Group 1~Group 5(图1A),其中Group 1~Group 3组群特征:花被片上有紫红色斑块,且斑块的面积逐渐减少;Group 4~Group 5特征:花被片上无斑块,其中Group 4的侧萼片分布均匀红点,而Group 5则为纯白花。
在P.‘B’和P.‘V3’的正反交后代中,不同组群所占总F1代比例的分布趋势一致,均为Group 5> Group 4> Group 2> Group 1> Group 3;同一组群在正反交后代中所占比例有所差别,表现为反交组合中Group 1~Group 3组群所占比例均大于正交组合,而反交组合中Group 4和Group 5组群所占比例则小于正交组合(表2)。表明P.‘B’ 为杂交亲本时其紫红色花斑可以遗传给后代,且后代会分离出面积及分布规律不同的紫红色花斑,此外,P.‘B’ 作为父本更容易将紫红色花斑这一性状遗传给子代。
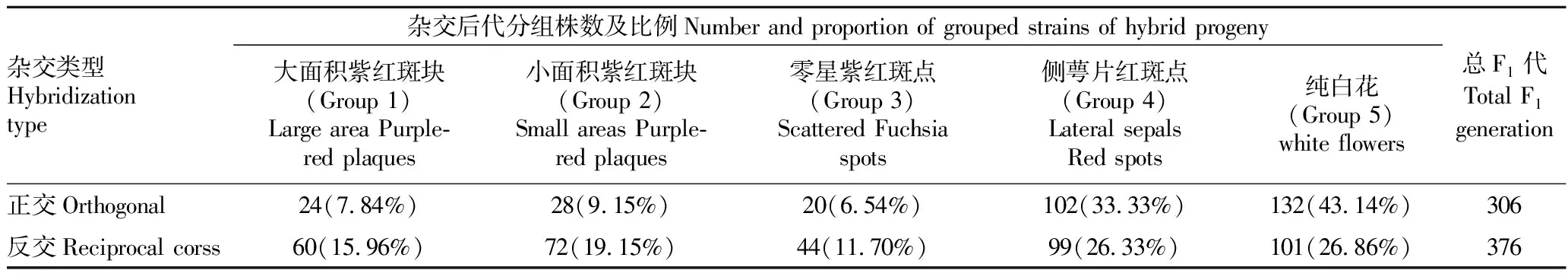
表2 蝴蝶兰正反交杂交F1代花斑分离比率
2.2.2正反交F1代唇部肉突颜色分离表现。供试亲本正反交后代(F1)中均分离出3种肉突颜色,分别为紫红色(图1B1)、黄底带紫红斑块(图1B2)及黄底分布均匀红点(图1B3)。在5个组群中,正反交组合的肉突颜色分布及所占比例规律基本一致,具体为Group 1~Group 3仅表现出紫红色和黄底带紫红斑块2种颜色,而Group 4和Group 5均表现黄底分布均匀红点,且随着紫红色花斑面积的增加,紫红色肉突所占正反交比例随之增加,均为Group 1> Group 2> Group 3;在总F1代中,正反交组合的肉突颜色趋势分布一致,但所占比例有所不同,黄底分布均匀红点最多,黄底带紫红斑块次之,紫红色最少,同时反交组合中紫红色和黄底带紫红斑块所占比例均比正交组合高(表3)。运用Spearman等级相关系数检验花瓣花斑面积与紫色肉突之间的相关性,得到的相关系数为0.975,说明花瓣面积越大,紫红色肉突出现的可能越大,显著性水平0.005<0.05,表明两变量之间的正向关系很显著。表明紫红色肉突与紫红色花斑的分布有一定的正向相关性,当花瓣上有紫红色花斑的,肉突颜色会显现紫红色,而花瓣上没有紫红色花斑的,肉突颜色则不会呈现紫红色,且会随着紫红色花斑面积的增加,紫红色肉突所占比例随之增加。 此外,P.‘B’为父本能提高杂交后代的肉突出现紫红色和黄底带紫红斑块的概率。
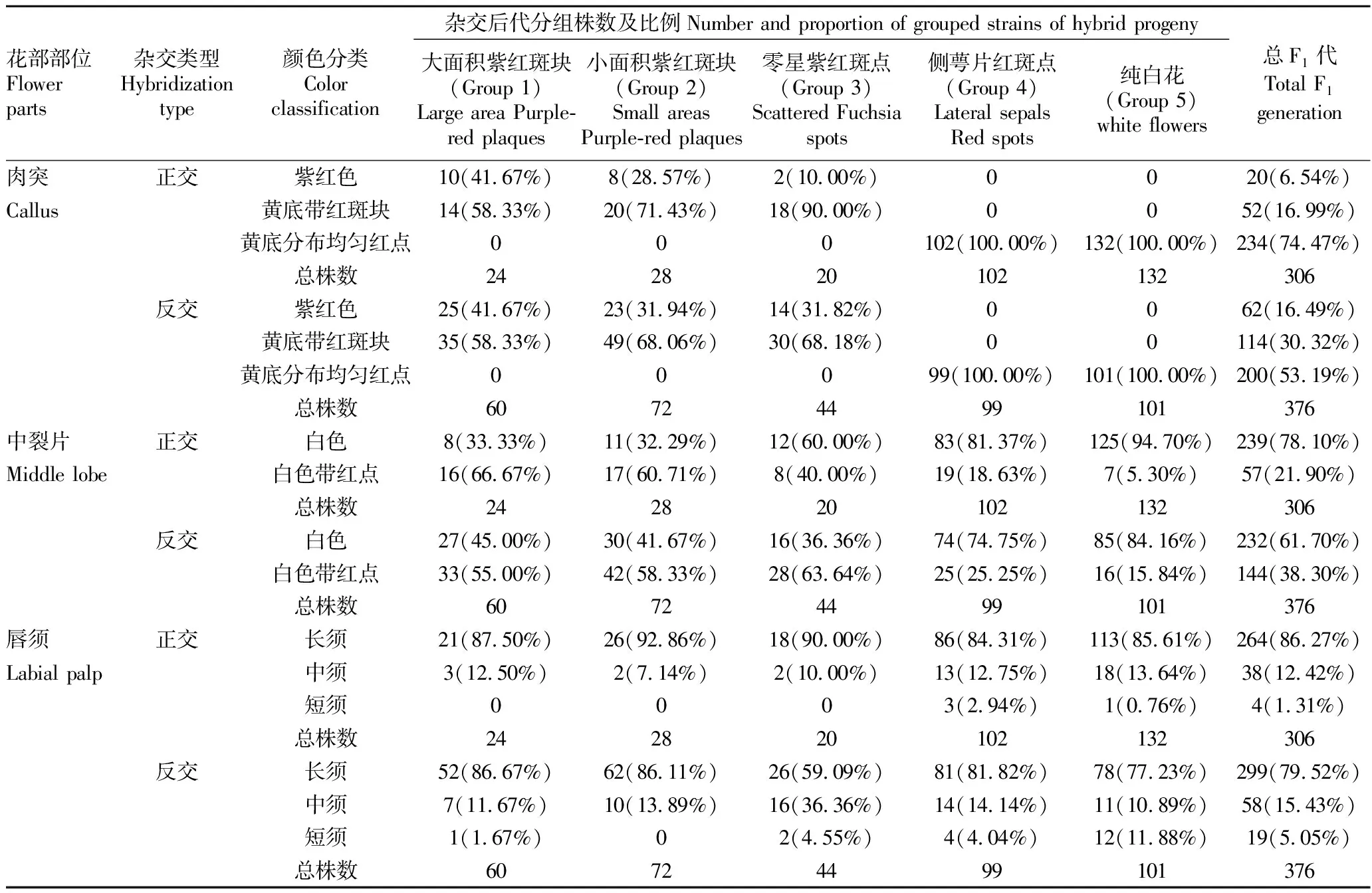
表3 正反交杂交F1代唇部性状分离比率
2.2.3正反交F1代唇部中裂片颜色分离表现。在P.‘B’ 和P.‘V3’ 杂交后代(F1)中,均分离出2种中裂片颜色,分别为纯白色黄唇、白色黄唇带些许红色点或红纹。在Group 1~Group 3中,正反交后代中裂片颜色的分布趋势和比例均存在一定差异,其中正交后代的中裂片为白色带些许红色所占比例大小为Group 1> Group 2> Group 3,而反交组合则刚好相反为Group 3> Group 2> Group 1;在Group 4和Group 5中,白色带些许红色的分布和比例基本一致,除Group5反交外,均为Group 4> Group 5,且反交组合中白色带些许红色的比例都比正交组合大;在总F1代中,正反交组合均表现一致,具体为白色带些许红色所占比例都小于纯白色中裂片,同时反交组合中白色带些许红色中裂片所占比例要高于正交组合(表3)。表明紫红色花斑分布对中裂片颜色有一定的影响,当花瓣上有紫红色花斑时,中裂片更易呈现红色,且P.‘B’为父本时杂交后代中这种遗传影响尤甚。
2.2.4正反交F1代唇须分离表现。在P.‘B’ 和P.‘V3’ 的正反交后代(F1)中,唇须类型均分离出3种,分别为长须、中须和短须,除Group 5外,正反交后代中的其他4个组群与总F1代的唇须比例规律基本一致,均为长须>中须>短须,不同组群的唇须分布差异不大(表3)。父母本均为长须,正反交后代结果表明长须和长须杂交更容易出现长须性状。
2.2.5正反交F1代花型及花瓣排列方式分离表现。在杂交后代(F1)中,正交组合后代花型分离出内弯和平整2种类型,分别占3.59 %和96.41 %,而反交组合则分离出内弯、平整及外弯3种类型,分别占比11.70%、86.70%和1.60%。结果表明杂交后代中出现了亲本没有的内弯和外弯,推测可能是受环境和遗传双重因素的影响,有待进一步的研究。杂交F1代花瓣均表现为分开,但分开的幅度有所不同,幅度最小花瓣几乎相接,幅度最大的花瓣接近父本。
3 讨论
蝴蝶兰的花瓣颜色、花斑分布、唇色、花型及花瓣排列方式等花部性状是主要的观赏部位,研究其后代的遗传分离规律,可为杂交育种中亲本的选择和新品种的选育提供理论参考。
目前已对蝴蝶兰花部性状在杂交F1代中性状分离规律开展了大量研究[12-15]。关于花斑的形成机理比较复杂,形成花部彩斑的原因主要有嵌合体和病毒病质体、位置效应、叶绿体的分离和缺失、易变基因的体细胞突变和各种类型的染色体畸变等[16]。该研究以P.‘B’ 和P.‘V3’ 为亲本进行正反交,通过亲本性状及其F1代的性状分离,发现正反交后代性状分离的类别、分布趋势及所占比例基本保持一致,这与李佐等[7-9]研究结果一致。表明P.‘B’ 为杂交亲本时其紫红色花斑可以遗传给后代,且后代会分离出面积及分布规律不同的紫红色花斑,此外,P.‘B’ 作为父本更容易将紫红色花斑这一性状遗传给子代。
关于肉突的颜色遗传尚未见相关的报道,该研究着重探究了花斑面积分布对肉突颜色的影响,结果表明紫红色肉突与紫红色花斑的分布有一定的正向相关性,当花瓣上有紫红色花斑的,肉突颜色会显现紫红色,而花瓣上没有紫红色花斑的,肉突颜色则不会呈现紫红色,且会随着紫红色花斑面积的增加,紫红色肉突所占比例随之增加。 此外,P.‘B’为父本能提高杂交后代的肉突出现紫红色和黄底带紫红斑块的概率。
蝴蝶兰花斑色的遗传规律研究除在杂交后代性状分离方面外,还集中在调控花青素合成的转录因子方面,Hsu 等[17-18]在小兰屿蝴蝶兰中鉴定出 3 个R2R3-MYB 转录因子,即 PeMYB2、PeMYB11 和 PeMYB12,推测三者分别参与了萼片和花瓣中红色色素沉着、红色斑点和条纹性状的形成,而唇瓣中底色和斑点的着色则由PeMYB12 和PeMYB11 决定,据此可以推测,MYB转录因子家族表达的时空特异性可能与蝴蝶兰花被片复杂多样着色模式的形成密不可分。随后,进一步证明了PeMYB11 转录因子对黑色花朵上深紫色斑点的形成起调控作用[18]。随着基因组、转录组、代谢组、蛋白组等多组学时代的到来,花色相关功能基因的挖掘将是未来兰花研究的热点[19],未来将继续加深对蝴蝶兰花斑、唇色等花色形成机理的探索。
蝴蝶兰花部性状在杂交后代中遗传研究虽取得了一定的进展,但仍存在不足之处。蝴蝶兰花色受多基因调控,机理复杂,后续将继续探索不同品种杂交后代观赏性状的遗传规律,为杂交育种选择亲本提供参考,为目标性状定向育种提供依据。
猜你喜欢
天津农业科学(2024年9期)2024-12-31 00:00:00
幼儿教育·父母孩子版(2023年5期)2023-06-04 23:40:07
山东林业科技(2022年3期)2022-07-14 07:54:20
舰船科学技术(2021年1期)2021-03-09 07:19:00
第二课堂(课外活动版)(2021年6期)2021-01-18 18:55:58
山东工业技术(2019年12期)2019-05-30 11:26:28
作文周刊·小学二年级版(2019年12期)2019-04-26 12:37:56
丝绸(2018年9期)2018-09-10 19:20:10
今日文摘(2017年8期)2017-04-25 12:07:12
大灰狼(2015年5期)2015-07-25 20:57:36
