
棕榈酸对人卵巢颗粒细胞脂质代谢和自噬的影响*
2023-09-05 01:30宁薇聂方钦张哲
医药导报 2023年9期
宁薇,聂方钦,张哲
(1.湖南省人民医院/湖南师范大学附属第一医院生殖医学中心,长沙 410005;2.中南大学湘雅三医院检验科,长沙 410013;3.中南大学湘雅三医院药学部,长沙 410013)
颗粒细胞是包绕在卵母细胞外层的多边形的细胞,通过缝隙连接和桥粒与卵母细胞连接,与卵泡发育密切相关。多囊卵巢综合征(polycystic ovary syndrome,PCOS)是一种育龄期女性最常见的排卵障碍性不孕症[1]。研究发现自噬可能是PCOS患者卵泡发育异常的重要原因[2]。自噬是细胞降解废弃蛋白质及细胞器的过程[3],维持着细胞内环境的稳定,但过度的自噬会导致细胞结构的破坏,引起细胞死亡。自噬体的积聚可诱导颗粒细胞死亡,从而影响卵母细胞的发育潜能。然而目前对卵巢颗粒细胞的自噬研究还较少。
脂质是参与代谢和内分泌调节以及生殖功能的细胞的重要组成部分[4],PCOS与脂质代谢异常有关,其中脂肪酸的急剧增加是PCOS患者的脂质代谢发生紊乱的重要原因之一[5]。棕榈酸(palmitic acid,PA)是一种长链脂肪酸,是机体内最常见的饱和脂肪酸。研究发现在PCOS患者卵泡液中PA的水平增加[6]。但是PA对颗粒细胞影响的机制尚不清楚。本研究以人卵巢颗粒细胞KGN细胞系为研究对象,观察PA对KGN细胞自噬平衡及脂质代谢的影响,并探讨该过程潜在的调控机制。
1 材料与方法
1.1材料与试剂 人卵巢颗粒细胞系KGN购自武汉普诺赛生命科技有限公司。PA(美国Sigma公司,货号:P0500,含量≥99%)、3-甲基腺嘌呤(3-methyladenine,3-MA)(美国MedChemExpress公司,货号:HY-19312,含量≥99%);油红O染色试剂盒(北京索莱宝科技有限公司,货号:G1262)、活性氧(reactive oxygen species,ROS)检测红色荧光试剂(北京普利莱基因技术公司,货号:C1300-2)、细胞增殖试剂盒(cell counting kit,CCK-8,美国APExBIO公司,货号:K1018);SREBP1鼠抗人多克隆抗体(英国Abcam公司,货号:ab28481);p62(货号:18420-1-AP)、LC3(货号:14600-1-AP)、PTEN诱导的假定激酶1(PTEN-induced putative kinase 1,PINK1)(货号:23274-1-AP)、Parkin(货号:14060-1-AP)、β-actin兔抗人多克隆抗体(货号:20536-1-AP)购自武汉三鹰生物技术公司。
1.2仪器与设备 全自动显微镜(日本 Olympus BX61),多功能酶标仪(美国Beckman DTX 880),凝胶成像分析仪(美国BIO-RAD 1708195)。
1.3细胞培养和干预 KGN细胞系采用含有10%胎牛血清(fetal bovine serum,FBS)的达尔伯克必需基本培养液/F12培养基于 37 ℃、5%二氧化碳(CO2)的培养箱中培养,当细胞融合至80%时进行传代。根据前期细胞活性测定预实验结果确定PA及3-MA刺激的浓度和时间,将处于生长对数期的KGN细胞分为正常对照组(无处理)、含有不同浓度PA处理组(100、200、400 μmol·L-1)、3-MA(3 mmol·L-1)处理组、3-MA(3 mmol·L-1)+PA(400 μmol·L-1)处理组。各组加药处理 24 h 后收集细胞进行后续测定。
1.4油红O染色试验 取对数生长期KGN细胞,以每孔5×105个细胞的密度接种于放爬片的6孔板上,细胞培养至密度60%~70%时,进行PA干预,正常对照组加不含PA的完全培基。放入孵箱培养24 h后,取出培养板,磷酸盐缓冲液洗3次,用4%多聚甲醛固定30 min,磷酸盐缓冲液洗3次,油红O染液室温染色30 min,纯化水洗30 s至背景透明,再用苏木精复染室温染色3 min,纯化水洗30~60 s,用水性封片剂封片,显微镜下观察细胞内的脂滴。
1.5Western blotting检测SREBP1、p62、LC3、PINK1、Parkin表达水平 使用RIPA细胞裂解液处理各组KGN细胞;制备十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electropho-resis,SDS-PAGE)凝胶,将蛋白样品上样到SDS-PAGE 凝胶加样孔内进行电泳;电泳结束后用湿式转移法将蛋白转印至聚偏二氟乙烯膜。室温下,经含5%脱脂奶粉的 TBST封闭1 h后,4 ℃条件下孵育一抗过夜后,37 ℃温箱中分别与相应的连接有辣根过氧化物酶的二抗孵育1 h,用TBST洗去未发生结合的二抗。利用化学发光试剂盒显影蛋白条带。胶片经扫描后,利用Image J定量对应条带的强度。
1.6CCK-8检测细胞活性 将处于对数生长期的KGN细胞制成细胞悬液,以每孔4×103个细胞的密度接种于 96 孔板中,待细胞贴壁后进行分组处理,各处理组于24 h后,每孔加入CCK-8溶液10 μL,培养箱中继续培养 2 h;用酶标仪测定其在波长 450 nm 处的吸光度(A值)。细胞活性(%)=处理组A值/正常对照组A值×100%,重复3次。
1.7细胞内ROS水平分析 将处于对数生长期的KGN接种于放有爬片的6孔板,常规培养贴壁后更换不同加药处理组的培养基继续培养4 h。吸去细胞培养液,用磷酸盐缓冲液洗涤3次,加入用无血清培养基稀释的DCFH-DA 荧光探针和细胞一起在37 ℃温箱孵育30 min,再用磷酸盐缓冲液洗涤细胞3次,将爬片置于荧光显微镜下进行观察。用 Image J 图像分析系统分析各实验组红色荧光强度。
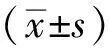
2 结果
2.1不同浓度PA对KGN细胞脂滴油红O染色结果 见图1。结果显示正常KGN细胞内未见明显橘红色脂滴。随PA刺激浓度增加,与正常对照组比较,细胞体积增大,细胞内脂滴形成增多,细胞质可见大量大泡样橘红色脂滴。且随着PA浓度的增加胞内脂滴数量增加更为明显。

图1 不同浓度PA对KGN细胞脂滴油红O染色的影响(n=3)
2.2不同浓度PA对KGN细胞SREBP1蛋白表达结果 与正常对照组比较,随PA刺激浓度增加,KGN细胞固醇调节元件结合蛋白SREBP1蛋白表达水平呈上调趋势,其中PA为400 μmol·L-1时对SREBP1表达上调最为明显(t=4.52,P<0.05),见图2。
①与正常对照组比较,t=4.52,P<0.05。
2.3PA抑制卵巢颗粒细胞活性 正常对照组、PA(100、200、400 μmol·L-1)处理组的KGN细胞活性分别为(100.00±0.00)%、(78.74±2.87)%、(57.48±6.50)%、(46.58±4.26)%。PA(100、200、400 μmol·L-1)处理组与正常对照组比较,差异均具有统计学意义(t=12.10、11.06、21.15,P<0.05)。随着PA浓度增加,KGN的活性呈下降趋势,其中PA浓度为400 μmol·L-1时对细胞活性的影响最为明显,表现出明显的抑制作用,故后续实验以400 μmol·L-1PA进行处理。
2.4自噬抑制剂3-MA对KGN细胞活性的影响 正常对照组、400 μmol·L-1PA处理组、3-MA组、PA和3-MA联合处理组的KGN细胞活性分别为(100.00±0.00)%、(28.11±1.24)%、(81.66±5.12)%、(37.26±1.28)%。400 μmol·L-1的PA 处理后,细胞的增殖受到明显抑制(t=78.11,P<0.01),加入3-MA预处理后,可逆转PA对细胞活性的抑制作用(t=8.87,P<0.05)。而3-MA单独作用细胞不引起细胞活性改变(P>0.05)。
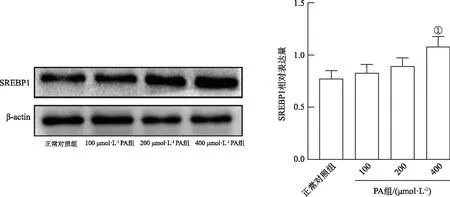
2.53-MA对PA诱导的KGN细胞ROS含量的影响 ROS荧光检测结果见图3。与正常对照组的KGN细胞比较,400 μmol·L-1PA处理4 h后的KGN细胞红色荧光信号强度增加,表明ROS水平升高(t=15.54,P<0.01),而使用3-MA预处理后ROS生成减少(t=5.04,P<0.05)。此外,3-MA单独作用细胞与对照组相比不引起ROS水平的改变(P>0.05)。
A.正常对照组;B.400 μmol·L-1PA组;C.3 mmol·L-13-MA组;D.3 mmol·L-13-MA+400 μmol·L-1PA组。①与正常对照组比较,t=15.54,P<0.01;②与PA组比较,t=5.04,P<0.05。
2.63-MA对PA诱导的p62、LC3-Ⅱ/LC3-Ⅰ表达的影响 Western blotting检测结果见图4,各实验组中均可见p62及LC3蛋白表达。3-MA单独处理组与对照组比较,LC3比值和p62相对表达水平均差异无统计学意义(均P>0.05)。而400 μmol·L-1PA处理组与对照组比较,p62相对表达水平与LC3-Ⅱ/LC3-Ⅰ比值均差异有统计学意义(t=14.58、10.75,P<0.05)。加入3-MA预处理后与PA组比较,p62表达下调(t=10.28,P<0.05),LC3-Ⅱ/LC3-Ⅰ比值减小(t=8.02,P<0.01)。

A.正常对照组;B.400 μmol·L-1 PA组;C.3 mmol·L-1 3-MA组;D.3 mmol·L-1 3-MA+400 μmol·L-1 PA组;①与正常对照组比较,P<0.01;②与PA组比较,P<0.05;③ 与PA组比较,P<0.01。
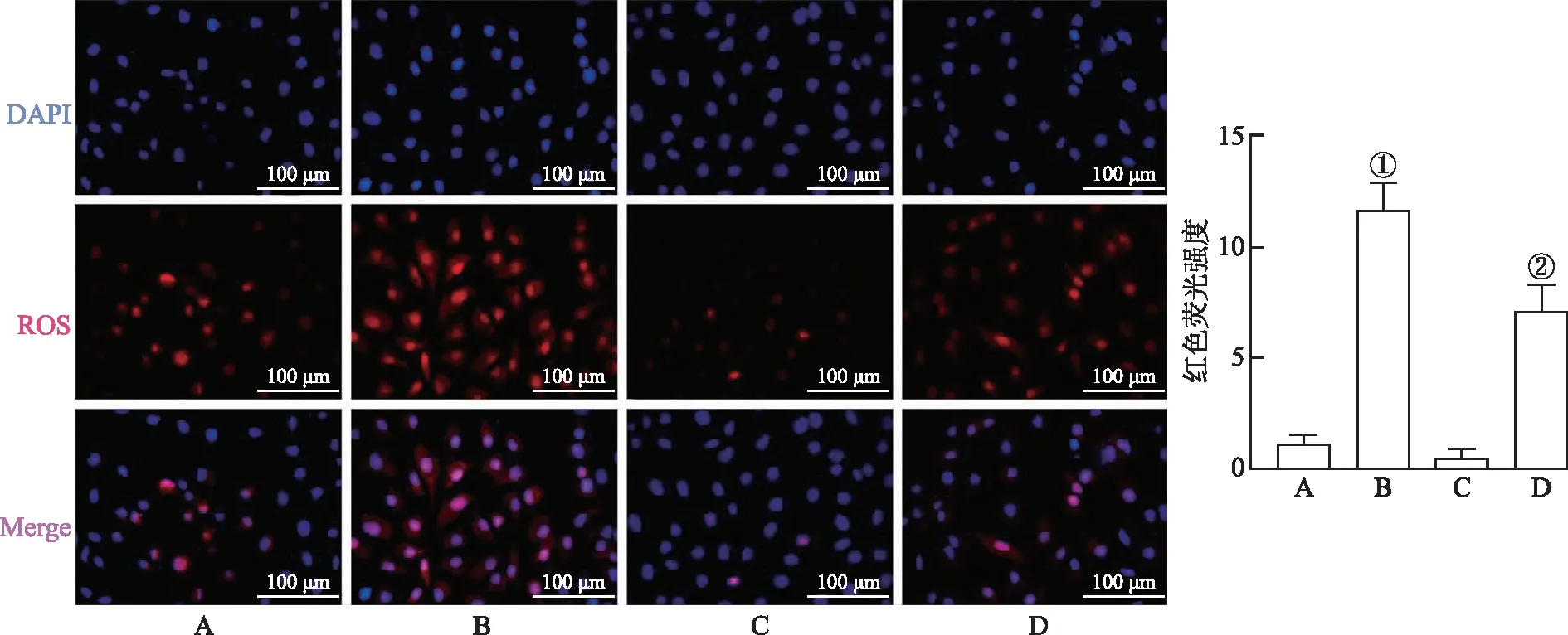
2.73-MA对PA诱导的PINK1、Parkin表达的影响 Western blotting检测结果见图5,与正常对照组比较,400 μmol·L-1PA处理组KGN细胞PINK1和Parkin蛋白表达水平升高(t=6.43、5.97,P<0.05)。使用3-MA预处理细胞后,与PA处理组比较,KGN细胞PINK1和Parkin蛋白表达水平降低(t=4.65、6.50,P<0.05)。
A.正常对照组;B.400 μmol·L-1PA组;C.3 mmol·L-13-MA组;D.3 mmol·L-13-MA+400 μmol·L-1PA组;①与正常对照组比较,P<0.05;②与PA组比较,P<0.05。
3 讨论
本研究表明PA对卵巢颗粒细胞具有脂毒性,导致颗粒细胞内脂质蓄积和细胞活性降低,ROS含量增加。细胞表型改变与PA诱导颗粒细胞氧化应激,激活线粒体自噬有关。
体内游离脂肪酸(free fatty acid,FFA)会引起心脏及外周器官细胞脂质含量增加,并转化为三酰甘油[7]。过量的FFA对颗粒细胞和卵母细胞发育是有害的。在以往的研究中证明了饱和游离脂肪酸尤其是PA,可以引发胰岛素抵抗,对卵泡发育产生负性影响[8]。本研究显示,随着PA浓度增加引起了颗粒细胞内脂滴数量的显著增加,脂肪酸合成途径的主调控因子SREBP1蛋白表达水平增加,伴随颗粒细胞活性下降,提示PA促进颗粒细胞内脂质蓄积,影响颗粒细胞脂质代谢,从而引起细胞损伤。
卵母细胞是人体内最大的细胞,其发育需要充足能量供应。除自身线粒体产生的能量外,还需要其周围的颗粒细胞线粒体氧化呼吸提供能量。颗粒细胞中含有丰富的线粒体,通过糖酵解途径将胞质中的葡萄糖代谢为丙酮酸,并转运到卵母细胞中用以产生三磷酸腺苷,维持卵母细胞的发育[9]。本研究发现PA引起颗粒细胞ROS含量增加,说明在PA刺激下颗粒细胞氧化应激加重,细胞损伤的效应细胞器主要在线粒体。3-MA为经典自噬抑制剂,可抑制自噬标记蛋白LC3-Ⅰ向LC3-Ⅱ转化,从而抑制自噬活性。为了进一步佐证PA引起的颗粒细胞线粒体损伤与自噬活性增加有关,笔者以3-MA为工具药,观察颗粒细胞在PA刺激下自噬的变化。结果显示3-M可逆转PA引起的氧化应激现象,证明了 PA引起颗粒细胞线粒体损伤与自噬活性相关。
正常情况下在一个生殖周期内,自噬是卵泡发育、生长和分化、闭锁所必需的[10-11]。据报道PCOS 患者卵巢组织中自噬活性增高,且在PCOS大鼠模型也发现自噬标记蛋白LC3-Ⅱ表达上调[12]。研究显示,p62等自噬相关蛋白会在 PCOS女性卵巢的膜细胞层中积累[13],本研究结果显示对照组也有p62和LC3-Ⅰ、LC3-Ⅱ的表达,说明体外培养的卵巢颗粒细胞内存在基础水平的自噬;加入PA处理后会明显引起卵巢颗粒细胞LC3-Ⅱ/LC3-Ⅰ比值和p62的表达增加,且可被3-MA逆转。p62为自噬接头蛋白,LC3-Ⅰ和LC3-Ⅱ是自噬体膜的结构蛋白。有研究认为当自噬被激活时自噬囊泡中p62等蛋白被溶酶体酶降解,p62水平降低,而LC3-Ⅱ/LC3-Ⅰ比值增高,但本研究显示p62与LC3-Ⅱ/LC3-Ⅰ变化趋同。p62变化存在偏差其原因可能是在正常细胞中,自噬不断发生p62蛋白不断降解以促使泛素化蛋白的清除。但在应激条件下,特别是在本研究中PA诱导的氧化应激的环境下,可以引发自噬功能缺陷进而导致p62蛋白聚集[14]。
当细胞受到氧化应激时极易对线粒体造成损伤,导致其功能障碍[15]。此时线粒体稳态平衡需要通过线粒体自噬实现。PINK1是线粒体自噬的主要调节因子,能够迅速有效地将E3泛素连接酶Parkin募集到受损的线粒体。Parkin促进线粒体膜上蛋白质泛素化,从而启动线粒体自噬以清除受损线粒体。PINK1和Parkin蛋白的过表达表明线粒体自噬增加,与线粒体损伤有关[16]。研究发现在PCOS患者的卵巢颗粒细胞中PINK1/Parkin被过度激活[17]。本研究也证实PA处理的卵巢颗粒细胞能够增加ROS含量和PINK1、Parkin蛋白的表达。且可被自噬抑制剂3-MA逆转,这说明PA可能过度激活PINK1/Parkin介导的线粒体自噬导致线粒体损伤,这可能是PA参与PCOS的发病机制。
本研究从卵巢脂质代谢切入探讨PCOS的发病机制及PA对卵巢颗粒细胞自噬平衡的影响,结果表明PA可以通过引起卵巢颗粒细胞脂质蓄积,增加ROS含量,激活PINK1/Parkin介导的线粒体自噬导致线粒体损伤损害生殖功能;这些发现将有助于从新的视角为PCOS的机制研究和治疗提供理论支持。本研究还存在不足之处,即仅在体外进行实验探究了 PA 对卵巢颗粒细胞自噬平衡的影响,自噬平衡破坏引发的 PCOS 的潜在机制尚未完全阐明。 关于 PA 对卵巢颗粒细胞自噬的调控及其相关作用机制还需要进一步研究证实。
猜你喜欢
服饰导报·鞋世界(2021年4期)2021-05-17
中国计划生育和妇产科(2021年9期)2021-04-18
中国临床医学影像杂志(2019年6期)2019-08-27
中国药理学通报(2019年5期)2019-01-11
中成药(2018年9期)2018-10-09
中成药(2018年1期)2018-02-02
中成药(2017年9期)2017-12-19
中成药(2017年4期)2017-05-17
癌变·畸变·突变(2014年1期)2014-03-01
中成药(2014年9期)2014-02-28
