
睡莲属植物花粉形态研究
2023-08-31 09:53:52戴忠良徐迎春刘春贵孙春青
西北植物学报 2023年7期
曹 婧,戴忠良,徐迎春,刘春贵,徐 婷,孙春青,*
(1 江苏丘陵地区镇江农业科学研究所,江苏句容 212400;2 南京农业大学,南京 210095;3 江苏里下河地区农业科学研究所,江苏扬州 225000)
睡莲科(Nymphaeaceae)有8~9属约100个种,其中睡莲属(Nymphaea)有50余种[1]。睡莲属植物根据心皮附属物的有无可分为Nymphaea亚属、Brachyceras亚属、Anecphya亚属、Lotos亚属和Hydrocallis亚属[2]。其中,Nymphaea亚属包括8种,在北半球的温带地区分布;Brachyceras亚属包括14~16个种,在热带和亚热带地区分布;Anecphya亚属包括7~10个种,主要在澳大利亚和新几内亚分布;Lotos亚属包括3~4个种,在热带地区分布;Hydrocallis亚属包括14个种,在热带地区分布[3]。尽管有些睡莲亚属的种数量少,但是睡莲属植物在生活史、心皮形态、繁育系统、传粉生物学等方面表现出显著的多样性[4-8]。
目前,从形态学[9]、解剖学[10]和分子生物学[3]已对睡莲属植物的系统分类和进化关系进行了相关研究,但仍存在互相矛盾的观点[2],因此,有必要从孢粉学角度研究睡莲属植物的系统进化关系。更重要的是,睡莲属处于被子植物系统进化的关键位置[11],并且睡莲属植物在生态和形态特征上具有多样性,因此,睡莲属植物孢粉学的特征对于研究早期被子植物生殖特征进化至关重要。但是先前的研究仅对睡莲属植物的数种花粉特征开展研究。例如,杨梅花等仅对4种睡莲的花粉形态进行研究,结果表明雪白睡莲(N.candida)的花粉形态特征在睡莲属植物中较原始[12]。
同样地,Zini等仅对Hydrocallis亚属的3个种花粉粒发育特征和超微结构进行研究,发现Hydrocallis亚属花粉的花粉壁较光滑,花粉壁无颗粒、棒状、疣状等纹饰[10]。现有睡莲属植物孢粉学研究缺乏多个亚属关键的花粉形态特征资料,因此需要对睡莲属植物未报道的花粉形态特征进行研究,并对其进行全面比较分析。此外,耐寒睡莲与热带睡莲跨亚属杂交的母本大多选择香睡莲块茎类型的耐寒睡莲[13],因此本研究对香睡莲的花粉活力进行评价,因为在睡莲杂交组合中母本自交结实率高更易获得跨亚属杂交种[14],而花粉生活力可直接影响自交结实率,以期为耐寒睡莲与热带睡莲的跨亚属杂交提供理论依据。
本研究利用扫描电镜对4个亚属的18种或品种睡莲花粉进行形态观察,测量和分析了花粉的形态特征,采用离体萌发法对‘彼得’的花粉活力进行测定,研究结果可为睡莲属植物的系统进化提供证据,也可为睡莲杂交育种亲本的选择提供依据。
1 材料和方法
1.1 供试材料
供试材料均保存在江苏丘陵地区镇江农业科学研究所水生植物资源保存圃。本试验选择4个亚属的18种或品种睡莲的具体性状特征见表1。2021年9月,待睡莲陆续开放,收集供试睡莲的成熟新鲜花粉。
1.2 方 法
1.2.1 睡莲花粉形态学研究将睡莲花药分别置于2.5%戊二醛(用pH 7.2的0.1 mol/L的磷酸缓冲液配制)溶液中4 ℃固定4 h固定8 h以上。
用生理盐水进行3次冲洗,每次10 min;再经50%乙醇→60%乙醇→70%乙醇→80%乙醇→90%乙醇→100%乙醇梯度脱水,每个处理15~20 min,将样品置于4 ℃过夜。
再将脱水过夜处理的花粉进行乙酸异戊酯置换、二氧化碳临界点干燥[15-17]。脱水和干燥后用双面胶将花粉均匀地粘在金属台上,真空喷金、镀膜后用Hitachi S-3000N型扫描电镜(日本Hitachi公司)进行观察。
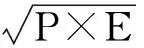

运用系统聚类组间联接的分析方法,根据平方欧氏距离测量,最终获得18种或品种睡莲属植物花粉聚类分析图,从而推断其系统进化规律以及它们的亲缘关系[20]。
1.2.3 睡莲花粉生活力研究采用离体培养法对睡莲品种‘彼得’进行花粉生活力测定。用吸管吸取2~3滴柱头液于载玻片作为培养液,选择第2天开放的耐寒睡莲花朵,于上午9:00用镊子沾取花粉播种于载玻片的柱头液上,后将载玻片放入垫有湿润滤纸的培养皿中,置于恒温光照培养箱,28 ℃培养4~6 h。
显微镜下观察花粉萌发情况,以花粉管长度大于或者等于花粉粒长度作为花粉管成功萌发的标准[21]。
随机选择3个视野,统计花粉粒数量及萌发数量,每个视野随机抽取3粒萌发的花粉粒,测量花粉管长度。
1.3 数据统计分析
数据采用Excel 2003进行统计和分析,图片处理采用Adobe Photoshop CS5。
2 结果与分析
2.1 花粉粒形态特征
利用扫描电镜对18种或品种睡莲属植物的花粉形态进行观察,发现18种或品种睡莲属植物的花粉都是单粒花粉。
如图版Ⅰ所示,18种或品种睡莲属植物花粉粒极轴/赤道轴比值范围为:0.62~0.89,形态均为近球体或扁球体,极面观为圆形或近圆形,赤道面观为圆形、长圆形或近圆形;其中,花粉粒极轴长度范围:15.1~27.9 μm,赤道轴长度范围:24.5~32.8 μm,巨花睡莲花粉粒最大,体积指数为30.3;埃及白花粉粒最小,体积指数为19.2;其余花粉粒体积指数在26左右(表2)。此外,埃及白睡莲、喀麦隆睡莲、印度红睡莲、墨西哥黄睡莲的花粉形态具有异极性。例如:墨西哥黄睡莲花粉的远极面光滑,而近极面为瘤状纹饰。
埃及蓝睡莲的萌发沟为单槽萌发沟,而其他17种睡莲属植物都为环槽萌发沟,且与赤道平行(图版Ⅰ和Ⅱ)。
栽培品种‘达尔文’和‘佛罗里达’为远级环槽类型,其他15个种或品种属于赤道环槽或者近赤道环槽。
香睡莲、‘达尔文’、‘火冠’、‘亚克’、‘佛罗里达’和‘征服者’的花粉萌发沟较浅。Brachyceras亚属的蓝星睡莲萌发沟内有环形嵴。
18种或品种睡莲属植物花粉外壁纹饰存在一定差异,类型也比较丰富(表3),大部分为小颗粒状、瘤状和小柱状,少数具有长条状、微波纹起伏状和网纹状(图版Ⅰ、Ⅱ、Ⅲ)。Lotos亚属喀麦隆睡莲、埃及白睡莲的花粉外壁表面均较为平滑,附属物少;而印度红睡莲的花粉外壁表面可观察到颗粒状纹饰。巨花睡莲、小花睡莲和睡莲子午莲的花粉外壁为颗粒状凸起纹饰。
蓝星睡莲和一些具有香睡莲块茎的种和品种(香睡莲、‘亚克’、‘火冠’)花粉外壁具有颗粒状凸起、棒状和刺状纹饰,只是纹饰的密度和高度有差异。
2.2 聚类分析
聚类分析结果如图1所示,18种或品种睡莲属植物可分为4个类群。第1类群有香睡莲、白睡莲、墨西哥黄睡莲和睡莲子午莲4个原生种以及‘宽瓣白睡莲’、‘达尔文’、‘火冠’、‘亚克’、‘佛罗里达’、‘征服者’和‘彼得’7个栽培种,其花粉主要为圆球形,几乎每个品种都有多个外壁纹饰类型,P/E值在0.70~0.84之间。第2类群有蓝星睡莲、小花睡莲和埃及蓝睡莲3个原生种,花粉为扁球形,其P/E值在0.70~0.73之间,体积指数相近(22.0~23.0),均有瘤状附属物。第3类群仅有巨花睡莲,花粉形态为圆球形,P/E值为0.85,外壁纹饰为微波纹起伏状,极轴、赤道轴等各项指标数据均较大。第4类群有印度红睡莲、喀麦隆睡莲和埃及白睡莲3个原生种,其P/E值在0.62~0.89之间,这3个原生种的体积指数、沟长远小于其他种(品种),表面纹饰单一,与其他3个类群又明显区别。
2.3 花粉生活力
‘彼得’花粉在28 ℃恒温光照培养箱生长6 h后,花粉萌发比例高达95%,如图2所示,睡莲花粉粒生长出花粉管,平均长度约为260 μm,花粉管生长状态好。因为睡莲授粉后花粉是落到柱头液中,经过长期进化选择,柱头液内含丰富的氨基酸、微量元素[21],为睡莲花粉发育提供了最有利的条件,使得花粉具有较好的育性。
3 讨 论
3.1 睡莲属植物花粉形态多样性及分类学意义
18种或品种睡莲属植物的花粉形态具有丰富的多样性。花粉形态的聚类分析结果可将18种或品种睡莲属植物分为4个类群,这一结果与基于核糖体DNA的ITS序列[3]和叶绿体基因[2]所建立的系统分类一致。18种或品种睡莲属植物的花粉萌发沟除了埃及蓝睡莲是单槽的,其余均为环槽形的,并且环槽形花粉沟的位置在花粉远极端和赤道间变化,而以前的研究结果都认为睡莲属植物的花粉萌发沟都是环槽类型的[22-23]。Brachyceras亚属的3个种都有明显区别于其他种的花粉形态特征,这为这些种的鉴定提供了依据。Anecphya亚属的澳大利亚巨睡莲的花粉体积最大,可能由于其所处的生态环境影响所致,这些特征也是鉴定这些种的可靠依据。Lotos亚属的花粉外壁纹饰较光滑,且花粉形态差异小,这与前人的研究结果[24]一致,可以利用花粉形态特征从睡莲属植物中将Lotos亚属区分开来。Nymphaea亚属中具有香睡莲块茎的种和品种的花粉外壁纹饰相似度较高,一般都具有棒状、刺状和颗粒状纹饰,只是纹饰的密度和高度有差异,这一特征可以区分大多数睡莲属植物中具有香睡莲血统的种和品种。根据上述特征,我们推断Nymphaea亚属的栽培品种‘亚克’、‘火冠’、‘佛罗里达’和‘征服者’都可能是香睡莲的杂交品系。此外,以Brachyceras亚属蓝星睡莲、Nymphaea亚属香睡莲为代表,其花粉粒表面小颗粒突出且数量多,且都具有棒状、刺状纹饰,这些特征可对花粉起到较好的保护作用,有利于结实和发育,因此这些种作为亲本杂交成功率大。
3.2 睡莲属植物花粉形态特征的系统进化趋势
花粉的形态特征,特别是花粉外壁纹饰、萌发沟等特征,对于了解睡莲属植物系统演化、亲缘关系及系统分类均有重要意义。花粉外壁表面纹饰特点可以反映植物进化的一般规律[25],花粉外壁表面较光滑的较为原始,其次为颗粒状凸起,再到棒状、刺状,最终演化为条纹状及网状[26]。本研究中,Lotos亚属中喀麦隆睡莲、埃及白睡莲和印度红睡莲的花粉形态相似,但三者的花粉外壁纹饰不同,其中喀麦隆睡莲、埃及白睡莲的花粉外壁表面均较为平滑,附属物少,说明它们在睡莲属植物中较为原始、进化程度较浅,而印度红睡莲的花粉外壁表面可观察到有颗粒状附属物存在,说明有进化发展的趋势;Zini等[23]发现Hydrocallis亚属的3个种花粉壁较光滑,表明Hydrocallis亚属和Lotos亚属的种在睡莲属植物中最原始,并且这2个亚属的种可能有共同的起源[24]。Anecphya亚属巨花睡莲、Brachyceras亚属的小花睡莲、Nymphaea亚属的睡莲子午莲的花粉外壁都有颗粒状凸起,Brachyceras亚属的蓝星睡莲、Nymphaea亚属的香睡莲花粉外壁的颗粒状凸起更加明显,且蓝星睡莲和香睡莲粉外壁在颗粒状凸起的基础上演变出棒状和刺状纹饰,说明睡莲属植物是不断进化的,其中Nymphaea亚属的香睡莲在睡莲属植物中进化程度较高。此外,Nymphaea亚属中的墨西哥黄睡莲花粉的远极面是光滑的,而近极面具有颗粒状凸起,说明墨西哥黄睡莲可能是睡莲属植物进化中的过渡类型。
关于睡莲属植物环槽萌发沟的起源一直有争议,现有两种假说。第一种假说认为环槽萌发沟起源于萍蓬草属(Nuphar)的单槽萌发沟[27]。本研究中埃及蓝睡莲的萌发沟为单槽萌发沟,而其他17种睡莲属植物都为环槽萌发沟,根据第一种假说,笔者推断埃及蓝睡莲的萌发沟为睡莲属植物的原始形态,而环槽萌发沟是睡莲属植物的进化形式。第二种假说认为睡莲属植物环槽萌发沟是单孔形式的,环槽是单孔结构的扩展,花粉粒的整个远极端半球可看作是单孔的修饰部分[27]。根据第二种假说,如果远极端半球可以看作是修饰盖,那远极端与近极端的超微结构不同[28]。在本研究中,Nymphaea亚属的白睡莲和墨西哥黄睡莲的远极面和近极面的超微结构不同,这一结果支持了第二种假说,但本研究其他睡莲属植物不存在这种现象。睡莲属植物萌发沟的进化历程可能为:远极单槽→远极环槽→赤道环槽[24]。根据这一进化历程,本研究中埃及蓝睡莲的萌发沟属于最原始类型,Nymphaea亚属白睡莲和睡莲子午莲属于较原始类型,其余原生种萌发沟都是较进化类型(赤道环槽)。此外,栽培品种中‘佛罗里达’和‘达尔文’属于远极环槽,可能是由于品种间杂交的原因。
3.3 睡莲属植物花粉形态特征的亲缘关系分析
花粉形态特征的稳定性强,可用于植物种间亲缘关系分析[29]。18种或品种睡莲属植物的聚类分析结果表明:Nymphaea亚属和Brachyceras亚属中的种和栽培种间的亲缘关系较近;Lotos亚属中印度红睡莲和喀麦隆睡莲亲缘关系较近,而它们与埃及白睡莲的亲缘关系较远;Nymphaea亚属和Brachyceras亚属间亲缘关系较近,许多育种者已经获得了这2个亚属的杂交种[13,30];Brachyceras亚属和Anecphya亚属亲缘关系较远,但是我们已经得到了这2个亚属的杂交种,说明利用花粉形态特征分析睡莲属植物的亲缘关系有一定的误差。Nymphaea亚属和Anecphya亚属、Lotos亚属与其他3个亚属间的亲缘关系远,至今无育种者获得这些亚属间的杂交种。
综上所述,花粉形态特征,特别是花粉萌发沟类型、外壁纹饰等特征可为睡莲属植物种间分类、亚属间以及种间亲缘关系、系统进化规律提供关键的证据。但孢粉学的研究对分析睡莲属演化、亲缘关系及系统分类具有一定的局限性,仍需通过形态学、解剖学和分子生物学等手段深入探讨和分析,以便更深层次地了解睡莲属植物系统演化过程。
猜你喜欢
儿童故事画报·自然探秘(2024年6期)2024-05-22 22:26:48
汽车零部件(2022年9期)2022-09-30 00:54:20
作物杂志(2021年6期)2021-12-17 07:20:24
制冷技术(2021年1期)2021-03-31 09:02:34
植物研究(2021年5期)2021-03-02 03:53:10
经济技术协作信息(2020年30期)2020-10-28 08:56:12
浙江农业学报(2020年1期)2020-03-07 05:07:12
浙江农业科学(2017年8期)2017-08-30 12:48:08
浙江农业科学(2016年5期)2016-12-07 07:03:27
——系统分类与检索*
寄生虫与医学昆虫学报(2015年3期)2015-11-12 00:32:38
