
北美洲当归属(伞形科)植物花粉形态特征研究
2023-09-03 09:38:14杨明乐廖晨阳
西北植物学报 2023年7期
杨明乐,廖晨阳
(四川大学 建筑与环境学院,成都 610065)
当归属(AngelicaL.)为伞形科(Apiaceae)芹亚科(Apioideae)中最具代表性的复杂类群之一。该属在全世界约有110种,广布于北温带的高山亚高山地区及部分高纬度地区[1-3];其中,北美洲为该属的重要分布地和多样性中心之一,据美国农业部植物数据统计有24种在美国和加拿大分布,另有墨西哥特有种Angelicanelsonii与新并入的Angelicacapitellata(原SphenosciadiumcapitellatumA. Gray)[4],由此北美洲当归属物种总数为26种[5]。
由于当归属植物的形态特征丰富,变异幅度较大,与近缘类群的关系也较为复杂,自该属建立以来其分类和系统问题备受学界关注[6-7]。近年来,基于分子系统发育和果实形态特征的研究表明,广义当归属(Angelicas.l.)成员主要分布在3个进化枝:当归支系(Angelicagroup),山芹支系(Ostericumclade)和滇芹支系(Sinodielsiaclade),前者又包括5个主要次级支系,即东亚分支(Eurasian clade),古当归分支(Archangelicaclade),高山芹分支(Coelopleurumclade),滨海当归分支(LittoralAngelicaclade)和北美当归分支(North AmericanAngelicaclade,简称NAAngelicaclade)[7]。
2022年Liao等的研究指出,北美洲当归属物种大部分归属于NAAngelicaclade,3种出现在古当归分支,2种隶属于东亚分支,而A.dawsonii应归属于“Perennial endemic North American(PENA)clade”;对NAAngelicaclade的进一步分析表明,该分支包括了高山芹分支在内的21个物种,其中14个物种呈单系,2个并系群分别组成了新组合A.lineariloba与A.venenose[5]。然而,分子系统学与果实形态学证据之间仍存在不少分歧,尚待寻找更充足的分类依据。
伞形科植物的花粉材料较成熟果实更易采集和保存,受外界条件的影响较小,且花粉的大小、形状、萌发孔类型以及外壁纹饰等特征具有较丰富的多样性,花粉形状包括近圆形、近菱形、椭圆形、近长方形、超长方形及赤道收缩形6种,极轴/赤道轴(P/E)1.5~3.0,体积指数(SI)20~40,表面纹饰具有或疏或密的长条纹状、短棒状脑纹,片状脑纹、皱波网状及颗粒网纹等多种类型[8]。茴芹属(PimpinellaL.)、滇芎属(PhysospermopsisWolff)、棱子芹属(PleurospermumHoffm.)、独活属(HeracleumL.)、莳萝属(AnethumL.)、胀果芹属(PhlojodicarpusTurcz. ex Bess.)和前胡属(PeucedanumL.)等属的花粉形态特征都为这些类群的分类处理和演化关系提供了重要依据[9-13],尤其是在种间水平具有较高的参考价值。张诗怡等报道了狭义当归属卫星类群山芹属(OstericumHoffm.)8种17个居群的花粉特征,并指出该属花粉形状与多种纹饰特征可为该属不同层次的分类鉴定与亲缘关系研究提供支持[14]。
迄今为止,涉及北美洲当归属植物的孢粉学研究极少,仅20世纪90年代,佘孟兰等[15]曾对东亚与北美的当归属植物花粉形态特征进行了对比,但由于当时条件和技术的限制,该研究的样本材料较单一、定量数据不足,因而未能全面对北美洲当归属植物的花粉进行研究。
本研究广泛收集了北美洲当归属植物的花粉材料[仅林当归(A.sylvestris)的材料采自欧洲],共计26种44个居群,运用光学显微镜与扫描电子显微镜进行了观察拍照,并利用SketchUp软件对花粉形状、纹饰密度、极轴及赤道轴长度等特征进行测量分析和定量定义,以期为上述类群的系统发育和分类修订提供更多依据。
1 材料和方法
1.1 材 料
试验材料取自University of Alaska Herbarium (ALA)、Herbarium of Brigham Young University (BRY)、Herbarium of California Academy of Sciences (CAS)、Herbarium of the Missouri Botanical Garden (MO)、Herbarium of the University of Illinois at Urbana-Champaign (ILL)、Herbarium of the Illinois State Natural History Survey (ILLS)、Herbarium of Illinois State University (ISU)、Robert F. Hoover Herbarium (OBI)、Herbarium of Oregon State University (OSC)和Rocky Mountain Herbarium (RM)等提供的馆藏腊叶标本,共计26种44个居群,试验样本及其凭证标本信息见表1。林当归的自然分布地仅达到北美洲的格陵兰岛,19世纪被引种到北美大陆东北部栽培;本研究未获得格陵兰岛的样本,因此,选用采自欧洲瑞士和冰岛的花粉材料。

表1 北美洲当归属植物花粉材料来源信息

1.2 方 法
1.2.1 光学显微镜观察采用Erdtman醋酸酐分解法对44个居群的北美洲当归属植物花粉进行处理。在BDM320数码显微镜下观察花粉形状、萌发沟孔等形态特征,并与电镜观察结果核对。
1.2.2 扫描电子显微镜观察经醋酸酐分解后的花粉,经过乙醇和纯净水清洗,取少量固定在硅片上,干燥后置于样品台上喷金镀膜,再用SU3500型扫描电镜观察并在500,3 000(3 500)与12 000的倍数下拍照。
1.2.3 SketchUp测量花粉数据在SketchUp(2020 pro)中导入电镜花粉照片,利用标尺工具对花粉的极轴以及赤道轴长度进行测量,每份样品选取形态完整规则的10~20粒花粉进行观测,计算平均值,并记录其最大值和最小值。

2 观察结果
2.1 花粉形态特征
通过光学及电子显微镜观察,供试北美洲当归属植物花粉观测结果见表2。供试花粉的赤道面观分为椭圆形、近长方形、超长方形和超长方形(赤道缢缩)4种形态;花粉大小为(21.1~32.0) μm×(10.6~17.1) μm,极轴与赤道轴的比值为1.5~2.8,体积指数为15.8~23.3;其中极轴长度变化幅度较大,最长者是A.callii达32 μm,最短者是A.dawsonii和A.lineariloba均为21.1 μm;赤道轴最长者是林当归(35)居群为17.1 μm,最短者是毛珠当归(A.genuflexa)(17)仅10.6 μm;P/E值最大者是毛珠当归(17)与A.capitellata高达2.8,最小者是A.dawsonii仅1.5;SI值最大者是林当归(36)为23.3,最小者是A.lineariloba为15.8。

表2 北美洲当归属26种44个居群植物花粉形态特征
2.2 花粉形状
根据赤道面形状以及极轴与赤道轴的比值,供试的北美洲当归属花粉主要为近长方形(P/E<2)、超长方形(P/E≥2)、赤道缢缩的超长方形(P/E≥2,且赤道缢缩)和椭圆形4种类型,其中多数样本为超长方形。
总体来说,有21个物种的花粉形状在种内稳定,但有5个物种具有两种不同形状的花粉。
2.2.1 椭圆形花粉属于椭圆形类型的花粉仅有3种,包括林当归、A.dawsonii以及A.venenosa,极轴与赤道轴的比值P/E依次为1.5,1.5与1.8。
2.2.2 近长方形花粉供试北美洲当归属植物花粉属于近长方形的花粉仅有3种,包括A.canbyi(13)、A.lineariloba、A.wheeleri(44),极轴与赤道轴的比值P/E依次为2.0,1.8,2.0。
2.2.3 超长方形花粉根据观察结果显示,本次试验花粉中有19种为超长方形,包括古当归(A.archangelica)、毛珠当归(18)、A.ampla、A.atropurpurea、A.breweri(7)、A.callii、A.dentata、A.grayi、A.hendersonii、A.kingii、A.lucida、A.nelsonii、A.pinnata、A.scabrida、A.tomentosa、A.triquinata、A.venenosa以及A.wheeleri(43),极轴与赤道轴的比值P/E为2.1~2.7。
2.2.4 超长方形(赤道缢缩)花粉观察发现,供试花粉中共有6种花粉形状为超长方形(赤道缢缩),包括毛珠当归(17)、A.arguta、A.breweri(8)、A.canbyi(12)、A.capitellata以及A.roseana,极轴与赤道轴的比值为2.4~2.8,花粉大小为(27.1~29.8) μm×(10.6~11.7) μm,体积指数为17.2~18.6。
2.3 萌发沟
根据观察结果,供试花粉萌发沟形状细长且均匀。对比之下,沟较长近达极较多的有20种27个居群,包括古当归、A.ampla、A.atropurpurea等;较短不达极的有毛珠当归(18)、A.arguta、A.canbyi(13)、A.kingii(24)、A.lucida(27)等9种12个居群;萌发沟几达极的有A.callii、A.capitellata、A.dawsonii、A.lineariloba、A.lucida(28)共5种。
根据上述结果,北美洲当归属花粉萌发沟的长度具有较强随机性,没有表现出显著的种间或种内特性。
2.4 花粉外壁纹饰
在扫描电镜下拍照并观察,供试的北美洲当归属花粉赤道区外壁纹饰有3种类型(表3),包括片状脑纹、长条状脑纹以及短棒状脑纹,3种条纹的蜡质条纹密集、凸起与清晰度递增。10种13个居群为短棒状脑纹;16种25种个居群为长条状脑纹;5种6居群为片状脑纹。少数居群具有附着物,包括A.ampla、A.californica(9)、A.callii、A.canbyi颗粒状以及其他形状。观察结果显示,北美洲当归属花粉表面纹饰抵达极区但是集中在赤道区,部分花粉极区纹饰趋于平滑,如A.venenosa,A.dentata和A.lineariloba三者的花粉尤为明显,大致呈现为由赤道区深刻密集的纹饰向两级逐渐变浅变疏的过渡特征。

表3 北美洲当归属26种44个居群植物花粉外壁纹饰特征

2.5 花粉的分类
根据电镜观测记录的数据,本研究按照花粉形状、P/E值、SI值、纹饰类型等逐级进行分类,最终将26种44个居群的花粉划分为4级3大类15小类(表4)。

表4 北美洲当归属植物花粉分类
3 讨 论
3.1 北美洲当归属花粉形态特征的多样性
据对北美洲当归属26种44个居群花粉的观察结果,该属花粉在形状大小、极轴与赤道轴长度比、萌发孔沟长度与外壁纹饰等特征上呈现一定多样性,但整体上低于东亚当归属植物。首先,供试花粉包含了椭圆形、近长方形、超长方形及超长方形(赤道缢缩)4种形状,而东亚当归属植物具有5种;整体上花粉形状在种内较为稳定,但毛珠当归、A.canbyi及A.wheeleri3个物种的花粉形状在居群间有较大差异,可能是因为地理隔离导致的种内分化;极轴与赤道轴长度比P/E和体积指数具有较大变化幅度,远高于该属东亚成员。其次,供试花粉萌发孔沟具有较短不达极、较长近达极以及长几达极3种类型,东亚成员同样具有丰富的萌发孔沟类型[15,18]。此外,外壁纹饰上均呈现典型脑状纹饰,分为短棒状脑纹、长条状脑纹和片状脑纹3种,种内纹饰的密集度、突起程度及附属物等特征在种间和居群间呈现出不同程度变异,东亚物种也具有类似现象。
3.2 观察结果的差异性
相较于佘孟兰等[15]的研究,本研究扩充了北美洲当归属花粉研究材料(包括该地区全部26个物种的44个居群),并采用SketchUp软件对花粉进行测量并计算了P/E值及SI值,同时对表面纹饰类型和密度进行了观测和定量定义。就花粉形态来看,首先,花粉赤道面观形状以超长方形以及赤道收缩形为主,约占84%,其他为椭圆形和近长方形,而前人认为以椭圆形花粉为主,可能是因对花粉形状的主观判断有偏差。第二,本次观察结果P/E值1.5~2.8,体积指数15.8~23.3,而在前人的研究结果中花粉P/E值相对偏小(1.3~2.3),体积指数相对偏大(17.0~26.9),与本次观察结果差异较大,原因可能是由影像清晰度不足或测量方法误差所导致。第三,本研究亦注意到居群间的变异幅度,尽管大多数物种的花粉形态特征比较稳定,但林当归、毛珠当归、A.canbyi、A.lucida、A.scabrida、A.venenosa等存在不同程度的种内分化(表2、表3)。
3.3 北美洲当归属种间亲缘关系分析
3.3.1AngelicadawsoniiAngelicadawsonii是落基山脉北段特有的多年生中小型草本,可见于加拿大艾伯塔省西南部和不列颠哥伦比亚省东南部,以及美国蒙大拿州西部和爱达荷州中北部的山区。该物种具有较小的复伞形花序(直径3~5 cm),发达的总苞片和小总苞片,亮黄色的花瓣,以及易分离的外果皮和截面呈五边形的胚乳,这些形态特征与其他当归属植物有显著不同。分子系统发育显示该物种位于PENA clade,可能与PENA clade成员Lomatiumbrdegeei(J.M. Coult. &Rose)和Podisterayukonensis(Mathias &Constance)有密切亲缘关系,发达的苞片和亮黄色花瓣亦是PENA clade的常见特征之一,因此提出该物种应被移出当归属。在本研究中,A.dawsonii的花粉椭圆形,P/E值1.5为观测样本中最小,体积指数亦较小,外壁纹饰较为特殊呈密集且较平滑的短棒状脑纹(图版Ⅱ,15),均与其他当归属椭圆形花粉呈现出显著的不同,支持上述系统发育的研究结论。
3.3.2Angelicavenenosa与A.dentataAngelicavenenosa是美国分布最广的当归属物种,其分布范围从中部密苏里州至东部纽约州,而A.dentata仅生长在佛罗里达州北部和佐治亚州南部的低洼林地。尽管两者的植株和叶片形态具有明显不同,但它们具有同类型的果实,且在ITS与cpDNA系统发育树中都呈现并系关系,因此被Liao等修订为1个新组合[5]。从本研究来看,A.venenosa和A.dentata的花粉特征极为相似,A.venenosa具有超长方形和椭圆形2种形状的花粉,外壁纹饰均呈稀疏且粗糙的片状脑纹、极区纹饰较浅,其中A.venenosa2个居群花粉的P/E=2.2、SI=17.6~18.2(图版Ⅰ,41、42),与A.dentata(图版Ⅰ,16)尤为相似,均属于I类型;而伊利诺伊州南部Johnson County (40)居群则为较特殊的椭圆形(图版Ⅰ,40)。上述结果与分子系统结果高度吻合,总体上支持该新组合的建立。
3.3.3Angelicalineariloba与A.scabridaAngelicalineariloba生长在加利福尼亚州和内华达州的内华达山脉中部和南部的高海拔地区,而A.scabrida仅生长于内华达山脉东南部的Spring Mountains一带。尽管二者叶形差异极大,但它们具有V型Ⅱ亚型果实,且在分子系统中A.scabrida嵌入了A.lineariloba居群间,因此将其合并为新组合[5]。本研究对A.lineariloba与A.scabrida花粉进行观察,结果显示,A.lineariloba花粉赤道面观为近长方形(图版Ⅰ,25),外壁纹饰为稀疏且平滑长条状脑纹,且极区纹饰趋于平滑,为Ⅱ类型(图版Ⅱ,25),A.scabrida花粉赤道面观为超长方形(图版Ⅰ,33、34),外壁纹饰为稀疏与密集且较粗糙的长条状脑纹,极区纹饰深刻,为I类型(图版Ⅱ,33、34),可见二者花粉特征存在较大差异,是其种间分化的有利证据之一。
3.3.4Angelicabreweri、A.californica、A.hendersonii、A.tomentosa、A.arguta、A.scabrida与A.lineariloba加利福尼亚州及邻近地区表现为北美洲当归属的多样性中心,种类多、分布集中且在分子水平上分化程度不彻底,例如A.breweri、A.californica、A.hendersonii、A.tomentosa、A.scabrida、A.lineariloba以及A.arguta,上述种在ITS系统树构成分支subclade IV,而在cpDNA系统树上分别形成2个并系类群[5]。在本研究中,A.breweri、A.californica、A.hendersonii与A.tomentosa的花粉赤道面观均为超长方形赤道略有缢缩(图版Ⅰ,7~10、Ⅰ,21~22、37),P/E值相近为2.3~2.5,体积指数相近为18.4~20.3,外壁纹饰为长条状脑纹或短棒状脑纹(图版Ⅱ,7~10、Ⅱ,21~22、37),表明其亲缘关系可能较近;尽管这4个物种的花粉外壁纹饰存在细微差异,但本研究结果仍支持上述4个物种近缘的结论(本研究未收集到A.arguta加州北部居群花粉)。然而,另外3个物种A.arguta、A.scabrida与A.lineariloba三者的花粉形态差异较大,分别为超长方形、超长方形(赤道缢缩)以及近长方形,P/E值分别为2.5~2.6,1.8以及2.6,外壁纹饰也具有一定差异,故花粉特征可为它们提供鉴别的依据。
3.3.5Angelicalucida的种内分化Angelicalucida分布广阔,可见于北美洲西部阿拉斯加州至加利福尼亚州北部地区,以及东北美洲东北部大西洋沿岸地区,亦有报道该物种也出现于东北亚西北太平洋海岸和岛屿(异名Coelopleurumgmelinii(DC.) Ledeb.)。Angelicalucida与A.saxatilisTurcz.、A.nakaiana(Kitag.) Pimenov曾组成独立的高山芹分支,但在最近的研究中该分支被置于NAAngelicaclade的基部。一般认为高山芹分支的物种在形态特征和分子水系的分化程度较低,然而从孢粉学证据来看,尽管三者花粉的体积指数较接近(SI=20~21),但形状具有较显著差异。例如,A.nakaiana的花粉为椭圆形(P/E=1.8),A.saxatilis与A.lucida的花粉为超长方形(P/E=2.1)。此外,A.lucida花粉具有一定程度的种下分化,本研究对A.lucida的北美洲东西海岸的3个居群的观测发现:东海岸居群的体积指数(SI=18.8)(图版Ⅰ,28)显著小于西海岸居群花粉(SI=20.7和21.0)(图版Ⅰ,26、27),花粉外壁纹饰为稀疏且粗糙的短棒状脑纹(类型Ⅰ-1-1-1)(图版Ⅱ,28),不同于西海岸居群稀疏且粗糙的长条状脑纹(类型Ⅰ-1-2-3,图版Ⅱ,26、27)。这些差异可能是因该物种在地理上的间断分布导致的。
3.3.6 毛珠当归毛珠当归主要分布在从加利福尼亚州北部沿海、俄勒冈州西部、华盛顿州的滨海地区到加拿大西部和阿拉斯加南部,甚至可达亚洲东北部沿海地区,A.polymorphaMaxim.分布于中国东北各地以及日本朝鲜,A.tianmuensisZ.H. Pan &T.D. Zhuang分布于中国浙江北部。三者分布区域较为分散,但叶轴和小叶柄膝状反曲是它们显著的共同特征,分子系统研究亦显示它们同属于东亚分支(Eurasian clade)且亲缘关系密切。本研究对毛珠当归的2个居群进行观察,结果显示,毛珠当归花粉的赤道面观为超长方形(类型Ⅰ-2)(图版Ⅰ,17、18),其主要居群间差异体现在P/E值与外壁纹饰类型(图版Ⅱ,17、18),这可能是由于岛屿效应导致的种内分化。同时也注意到,毛珠当归(18)与A.polymorpha和A.tianmuensis花粉赤道面观都为超长方形(图版Ⅰ,18),P/E值相近分别为2.3~2.4,体积指数也相近(分别为18.8,17.11与18.2),外壁纹饰均为密集短棒状脑纹[19](图版Ⅱ,18),表明其亲缘关系应十分接近,支持以上3个类群近缘的结论。
3.3.7 古当归、Angelicaampla与A.atropurpurea组成古当归分支的6个物种在中亚、欧洲和北美洲广泛分布,其中古当归出现在欧洲和北大西洋的岛屿上,而A.ampla和A.atropurpurea则是北美洲特有物种。在本研究中,古当归、A.ampla与A.atropurpurea的花粉都属于第I大类,花粉形态均为超长方形,P/E值极为相似分别为2.2~2.5,体积指数分别为17.9~19.6(图版Ⅰ,1、2、5、6),比较接近,前两者的外壁纹饰为密集且较粗糙的短棒状脑纹,后者为密集且粗糙的长条状脑纹(图版Ⅱ,1、2、5、6)。

图版 Ⅰ 电镜下北美洲当归属花粉形态(赤道面观)PlateⅠ Pollen morphologies of Angelica species from North America under SEM (the equatorial view)1. A. ampla (×3 000); 2. 古当归 A. archangelica (×3 000); 3. A. arguta (×3 000); 4. A. arguta (×3 000); 5. A. atropurpurea (×3 000); 6. A. atropurpurea (×3 500); 7. A. breweri (×3 500); 8. A. breweri (×3 000); 9. A. californica (×3 000); 10. A. californica (×3 000); 11. A. callii (×3 500); 12. A. canbyi (×3 000); 13. A. canbyi (×3 000); 14. A. capitellata (×3 000); 15. A. dawsonii (×3 000); 16. A. dentata (×3 000); 17. 毛珠当归 A. genuflexa (×3 500); 18. 毛珠当归 A. genuflexa (×3 000); 19. A. grayi (×3 000); 20. A. grayi (×3 000); 21. A. hendersonii (×3 000); 22. A. hendersonii (×3 000); 23. A. kingii (×3 000); 24. A. kingii (×3 500); 25. A. lineariloba (×3 000); 26. A. lucida (×3 000); 27. A. lucida (×3 000); 28. A. lucida (×3 000); 29. A. nelsonii (×3 000); 30. A. pinnata (×3 000); 31. A. pinnata (×3 000); 32. A. roseana (×3 000); 33. A. scabrida (×3 000); 34. A. scabrida (×3 000); 35. 林当归 A. sylvestris (×3 000); 36. 林当归 A. sylvestris (×3 500); 37. A. tomentosa (×3 000); 38. A. triquinata (×3 000); 39. A. triquinata (×3 000); 40. A. venenosa (×3 000); 41. A. venenosa (×3 000); 42. A. venenosa (×3 000); 43. A. wheeleri (×3 500); 44. A. wheeleri (×3 000).
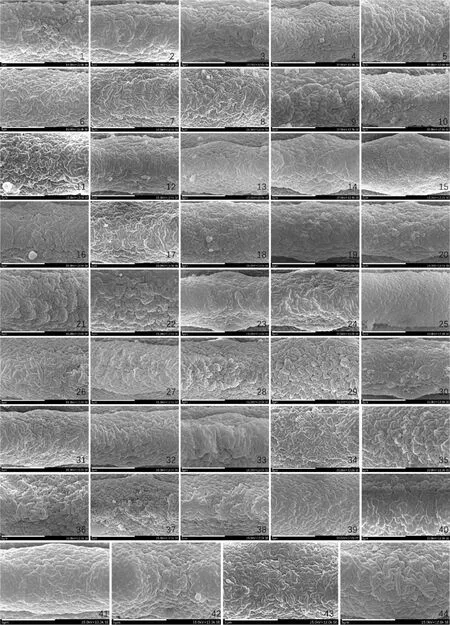
图版 Ⅱ 电镜下北美洲当归属花粉外壁赤道区纹饰(×12 000)Plate Ⅱ The equatorial ornamentation of exine walls of Angelica pollens from North America under SEM (×12 000)1. A. ampla; 2. 古当归 A. archangelica; 3. A. arguta; 4. A. arguta; 5. A. atropurpurea; 6. A. atropurpurea; 7. A. breweri; 8. A. breweri; 9. A. californica; 10. A. californica; 11. A. callii; 12. A. canbyi; 13. A. canbyi; 14. A. capitellata; 15. A. dawsonii; 16. A. dentata; 17. 毛珠当归 A. genuflexa; 18. 毛珠当归 A. genuflexa; 19. A. grayi; 20. A. grayi;21. A. hendersonii; 22. A. hendersonii; 23. A. kingii; 24. A. kingii; 25. A. lineariloba; 26. A. lucida; 27. A. lucida; 28. A. lucida; 29. A. nelsonii; 30. A. pinnata; 31. A. pinnata; 32. A. roseana; 33. A. scabrida; 34. A. scabrida; 35. 林当归 A. sylvestris; 36. 林当归 A. sylvestris; 37. A. tomentosa; 38. A. triquinata; 39. A. triquinata; 40. A. venenosa; 41. A. venenosa; 42. A. venenosa; 43. A. wheeleri; 44. A. wheeleri.
虽然上述3种的花粉外壁纹饰存在一定差异,但P/E值与体积指数相近,表明其亲缘关系可能接近。产于中亚的近缘种A.brevicaulis(Rupr.) Rchb.和A.decurrensLedeb.也都为超长方形花粉(P/E=2.4和2.3),但可能因观测方法不同,其体积指数(分别为22.4和16.4)与上述3个种花粉存在一定差异[19]。总的来说,古当归分支的花粉多样性较低,与分子系统发育和果实解剖的研究结论[5]一致。
致谢:四川大学建筑与环境学院张诗怡、高庆和四川大学分析测试中心王辉协助了试验的完成;美国University of Lllinois,Urban-Champaign的(UIUC) Dr. S. Downie和D. Katz-Downie在采集材料和研究过程中给予了重要帮助;Missouri Botanical Garden(MO)、University of Alaska Herbarium (ALA)、Herbarium of Brigham Young University(BRY)、Herbarium of California Academy of Sciences(CAS)、Herbarium of the Missouri Botanical Garden (MO)、Herbarium of the University of Illinois at Urbana-Champaign(ILL)、Herbarium of the Illinois State Natural History Survey (ILLS)、Herbarium of Illinois State University (ISU)、Robert F. Hoover Herbarium (OBI); Herbarium of Oregon State University(OSC)和Rocky Mountain Herbarium (RM)等标本馆为本研究提供了植物标本和花粉材料。
猜你喜欢
军事文摘(2024年2期)2024-01-10 01:58:44
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
天天爱科学(2022年4期)2022-05-23 12:41:40
军事文摘(2021年22期)2022-01-18 06:22:38
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
青少年科技博览(中学版)(2020年1期)2020-04-21 08:57:40
奥秘(创新大赛)(2019年4期)2019-04-15 07:13:08
草地学报(2018年5期)2018-11-07 02:25:00
小布老虎(2016年4期)2016-12-01 05:45:55
专用汽车(2016年5期)2016-03-01 04:14:40
