
喷施铜和硼肥对忍冬生理生化及其茎藤指标成分的影响
2023-09-03 09:41:18彭雅萍陈德奥陆莹颖孙莉琼刘仙华唐晓清王康才
西北植物学报 2023年7期
彭雅萍,陈德奥,陆莹颖,孙莉琼,刘仙华,唐晓清*,王康才
(1 南京农业大学 园艺学院,南京 210095; 2 江西峡江南农园 农业发展有限公司,江西峡江 331400)
忍冬藤为忍冬科植物忍冬(LonicerajaponicaThunb.)的干燥茎枝,清热解毒,疏风通络,常用于温病发热,热毒血痢,痈肿疮疡,风湿热痹,关节红肿热痛[1],还具有抗氧化、抗炎、抗菌作用[2-4]。目前对忍冬的研究主要集中在不同品种、产地、采收期及氮、磷、钾配方施肥对金银花产量及质量的影响等方面[5],而不同微肥配施对其影响的研究甚少。
微量元素在药用植物次生代谢产物合成与积累过程中起着重要作用,可作为某些有机合成反应的催化剂或参与植物有效成分的结构功能而影响其化学成分的形成和积累,从而最终影响药材的药理活性及药材内在质量[6]。其中,施用铜和锌元素可使忍冬碳水化合物代谢、蛋白质代谢等生理活动更加活跃,也可大大增加其指标性成分的合成量[7];叶面喷施微量元素可有效提高水稻对微量元素的吸收利用[8];铜在植物中不仅有稳定植物体内有关化合物的作用,还可作为电子传递体参与氧化还原反应[9];铜元素也常参与植物糖代谢的调节,并能影响糖在植物体内的转运[10]。硼是植物不可或缺的元素,它参与了植物碳水化合物的运转;植物糖分运输在缺硼时受到抑制,碳水化合物不能及时运输到根部;硼肥也对合理调控药用植物体内物质的运输与分配从而改善药用植物品质具有重要作用[11]。
目前,关于微肥铜与硼对忍冬生理生化指标和茎藤品质影响的相关报道较少,本试验采用叶面喷施铜、硼肥的方法,研究忍冬叶片光合气体交换参数、光合色素含量和忍冬藤的营养、药用成分等生理生化指标,以及茎藤品质的变化,探索忍冬铜、硼肥配施的最佳施用组合,以期为优化忍冬栽培技术提供理论参考。
1 材料和方法
1.1 材料及仪器
试验材料中忍冬种苗选自山东临沂基地,经南京农业大学王康才教授鉴定为忍冬科植物忍冬(LonicerajaponicaThunb.)4年生苗,试验地点为南京农业大学的设施大棚;试剂硫酸铜(CuSO4·5H2O)、四硼酸钠(Na2B4O7·10H2O)(纯度≥99%)等试剂均为分析纯,乙腈为色谱纯。试验仪器包括LI-6800型光合仪(美国LI-COR公司)、UV-1800型紫外可见分光光度计、Waters UPLC超高效液相色谱仪、eppendorf Centrifuge 5810R离心机、微波消解仪MARS6、等离子体发射光谱仪ICAP 7000等。
1.2 试验设计
选用长势一致的4年生忍冬植株,于2020年10月5日移栽到塑料盆(内径24 cm,高度27 cm)内,每盆栽种忍冬1株。供试土壤的基本理化性质为:pH 6.80,总氮1.554 g/kg,有机质27.40 g/kg,有效磷25.64 mg/kg,速效钾323.60 mg/kg。试验中铜肥(硫酸铜溶液)设置10,30,50 mg/L 3个质量浓度水平(分别以Cu10、Cu30、Cu50表示),硼肥(四硼酸钠溶液)设置为20,40,60 mg/L 3个质量浓度水平(分别以B20、B40、B60表示),以喷施清水为共同对照(CK),共组成铜、硼肥单施处理6个(3+3)、配施处理9个(3×3)以及对照1个,共计16个处理,每个处理重复3次。铜、硼肥采用叶面喷施方式,在15:00进行处理液喷施,以叶片正反两面全部湿润且无滴下为宜。试验于11月9日开始处理,每隔7 d喷施1次,每次喷施处理溶液体积为50 mL,共持续喷施3个月,合计12次。处理期间其他管理措施保持一致。
1.3 测定项目及方法
于2021年2月1日选取忍冬新生茎枝第4节处的对生叶片测定光合气体交换参数、叶绿素含量,采集的忍冬茎枝105 ℃杀青5 min,置于40 ℃烘箱中烘干至恒重,粉碎过80目筛,用于测定营养物质与药效成分的含量。
1.3.1 叶片光合生理指标采用光合仪测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),以忍冬新生茎枝第4节处的对生叶片为测定对象。同时,以95%乙醇提取法测定忍冬茎枝新鲜叶片的叶绿素a、b含量[12]。
1.3.2 茎枝营养物质含量茎枝样品中可溶性糖含量采用蒽酮比色法测定[12],并作适当修改;样品中淀粉、可溶性蛋白以及还原糖含量均采用南京建成生物研究所试剂盒测定。以芦丁(LOT:T27F10Z81699)为标准品,采用硝酸铝-亚硝酸钠比色法测定样品中总黄酮的含量[12]。
1.3.3 茎枝绿原酸含量以绿原酸(LOT:Y228K36544)为标准品,采用UPLC法测定样品中绿原酸含量[1]。色谱条件:C18柱(5 μm,250×4.6 mm),流动相为乙腈-0.4%磷酸溶液(10∶90),进样体积为5 μL,检测波长为327 nm,流速为0.7 mL/min,柱温30 ℃。分别以绿原酸的色谱峰面积(Y)与其对应的含量(X,mg/L)作标准曲线,计算回归方程,得到Y=2 235.7X+9 863.9,R2=0.999 8(n=3),线性范围是0~70 μg/mL。
1.3.4 马钱苷含量以马钱苷(LOT:P24O11F128801)为标准品,用UPLC法测定各样品中马钱苷含量[1]。色谱条件:C18柱(5 μm,250×4.6 mm),流动相为乙腈-0.4%磷酸溶液(12∶88),进样体积为5 μL,检测波长为236 nm,流速为0.7 mL/min,柱温30 ℃。分别以马前苷的色谱峰面积(Y)与其对应的含量(X,mg/L)作标准曲线,计算回归方程,Y=9 914.9X+8 992.8,R2=0.999 1(n=3),线性范围是0~70 μg/mL。
1.3.5 矿质离子含量样品消解:采用微波消解法。所有器皿均在10%硝酸中浸泡24 h后用去离子水洗净,烘干备用。精密称取0.3 g置于内插管中,加入5 mL HNO3,轻摇是混合均匀,盖好内插管盖后放入微波消解罐中消解30 min,冷却后开罐,用去离子水将消解液定容至50 mL,混匀后备用。样品测定:ICAP 7000等离子体发射光谱仪。射频功率1 150 W,辅助器流量0.5 L/min,等离子体气流量12 L/min,蠕动泵转速50 r/min。
1.4 数据处理
所有数据用SPSS 24.0进行单向方差分析,数据结果用“Mean±SD”表示。
2 结果与分析
2.1 铜硼肥配施对忍冬光合生理指标的影响
2.1.1 光合气体交换参数由表1可知,忍冬叶片的净光合速率(Pn)在喷施适宜质量浓度的铜硼肥(B20、Cu30、B20Cu50、B40Cu50以及B60Cu10)后比对照均有一定程度提高,尤以B40Cu50配施处理最高,同时也有部分处理比对照不同程度下降,但除Cu50、B20Cu10、B40Cu10和B40Cu30处理显著低于对照外,其余处理与对照差异均未达到显著水平;与Cu10、Cu30处理相比,当喷施高质量浓度的铜肥(Cu50)时,忍冬叶片的Pn明显降低,这可能是由于喷施铜肥质量浓度过高对植物叶片产生了毒害作用。同时,在配施铜硼肥处理下,忍冬叶片的气孔导度(Gs)与蒸腾速率(Tr)呈正相关,即气孔导度越大,蒸腾速率越高,尤以B60Cu10处理表现最显著;而喷施铜硼肥后,忍冬叶片的Pn与胞间CO2浓度(Ci)呈负相关,即胞间CO2浓度越高,净光合速率越低,尤以B40Cu50处理表现最显著。综合分析以上可知,B40Cu50处理最有利于忍冬叶片光合作用,B60Cu10处理次之。

表1 不同浓度铜硼肥处理下忍冬叶片光合气体交换参数和叶绿素含量
2.1.2 叶绿素含量叶绿素a、b是植物进行光合作用的主要色素,并且在光合作用的光吸收阶段起核心作用。表1显示,忍冬叶片叶绿素a含量在单施铜肥、单施硼肥和铜硼肥配施条件下分别以Cu30、B40和B40Cu50处理最高,均不同程度高于对照但未达到显著水平;所有施肥处理中仅Cu10处理叶绿素a含量与CK差异显著(P<0.05),且具有最低值。叶绿素b含量在单施铜肥、单施硼肥和铜硼肥配施条件下分别以Cu10、B40和B60Cu30处理最高,也均不同程度高于对照,且B60Cu30处理增幅达到显著水平;铜肥质量浓度越高,叶绿素b含量就越低,而单独施加铜肥或单独施加硼肥处理的叶绿素b含量普遍高于铜硼肥配施处理,但仅B60Cu30处理与对照差异显著。
总叶绿素含量在单施铜肥、单施硼肥和铜硼肥配施条件下分别以Cu30、B40和B60Cu30处理最高,但所有喷施处理均与对照无显著差异(P>0.05)。
2.2 铜硼肥配施对忍冬藤营养物质含量的影响
植物的营养物质不仅为植物的生长发育提供能量,还可以作为渗透调节物质维持细胞生长,有利于植物生理生化过程正常进行,从而维持植物的正常生长。表2结果表明,忍冬藤中可溶性蛋白含量在B60、Cu10、Cu50、B40Cu30、B40Cu50和B60Cu50处理下均显著高于CK,但这些施肥处理间均无显著差异,其中B60Cu50处理可溶性蛋白含量最高(5.43 mg/g),较CK显著增加了44.80%;其余施肥处理可溶性蛋白含量与对照均无显著差异。同时,忍冬藤中可溶性糖含量在所有铜硼施肥处理下大多较CK普遍增加,在单施铜肥、单施硼肥和铜硼肥配施条件下分别以Cu30、B60和B20Cu10处理最高,并以B20Cu10处理最高(11.70 mg/g),较CK增加了32.50%。忍冬藤中淀粉含量在铜硼肥配施处理下均较CK普遍增加,在单施铜、硼肥处理下大多较CK降低,其中最低的B40处理(149.73 mg/g)较CK下降了38.07%,最高的B40Cu50处理(392.46 mg/g)较CK增加了62.31%。但是,所有铜硼肥处理忍冬藤的可溶性糖和淀粉含量均与CK不存在显著差异。另外,忍冬藤中还原糖含量在各铜硼施肥处理下大多较CK显著增加,其中B20、B40Cu50处理的还原糖含量最高(均为1.09 mg/g),较CK显著增加了47.30%。以上结果说明适宜质量浓度的铜硼肥处理可显著提高忍冬藤中可溶性蛋白和还原糖含量,对可溶性糖和淀粉的积累也有一定促进作用,且铜硼肥配施处理的效果更明显;铜硼肥对忍冬藤中还原糖含量的影响最大,对可溶性蛋白含量影响次之,对可溶性糖和淀粉影响较小。
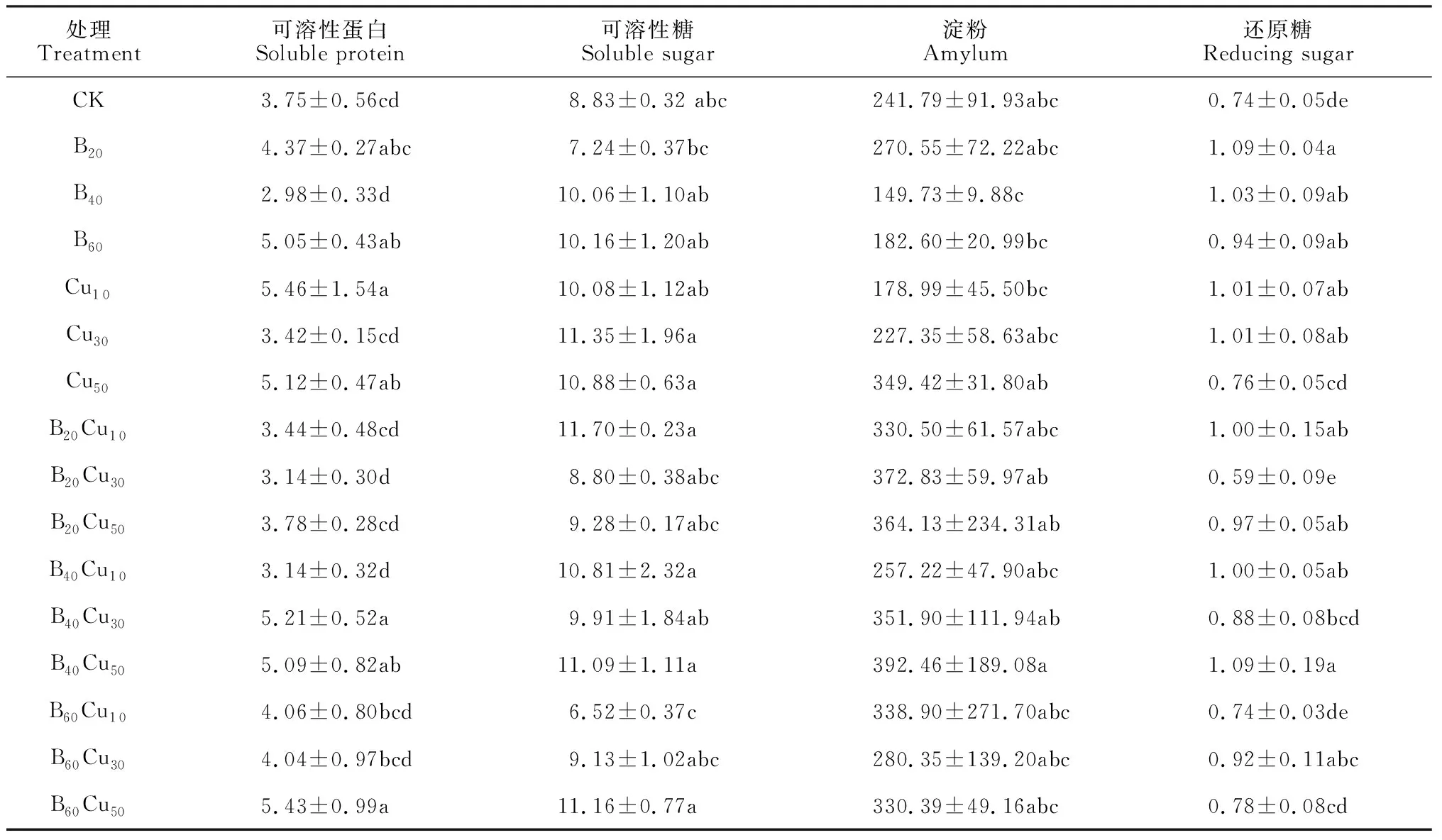
表2 不同质量浓度铜硼肥下忍冬藤营养物质含量的变化
2.3 铜硼肥配施对忍冬藤药效成分含量的影响
忍冬藤中的绿原酸、马钱苷、总黄酮为次生代谢产物,大量的次生代谢产物可增强植物自身的免疫力和抵抗力,同时也是作为评价忍冬藤药用质量的重要指标。
表3显示,叶面喷施铜硼肥对忍冬藤中绿原酸、马钱苷、总黄酮含量均有显著性影响(P<0.05)。其中,忍冬藤中绿原酸含量在各施肥处理下均比对照不同程度提高,除B40、Cu30和B20Cu50处理外增幅均达到显著水平,且铜硼肥配施处理的增幅普遍更大,并以B60Cu50处理绿原酸含量最高,增幅最大(453.85%);忍冬藤中马钱苷含量除B60、B20Cu50和B60Cu10处理外均比CK显著增加,其中B20Cu30处理的马钱苷含量最高,增幅最大(48.61%);忍冬藤中总黄酮含量除B20Cu30处理外均比CK显著增加,并以B60处理的总黄酮含量最高(5.64 mg/g),增幅最大(195.29%)。这说明铜硼肥处理对忍冬藤中绿原酸积累促进作用最大,且铜硼肥配施处理的效果更明显,而对总黄酮含量影响较大,其中单施铜肥或硼肥的影响更大。

表3 不同质量浓度铜硼肥处理下忍冬藤指标成分的变化
2.4 铜硼肥配施对忍冬藤矿质离子含量的影响
植物中的矿质元素为植物生长所必需,不仅可以提高植物体内酶的活性,还可促进植物中营养物质的积累。表4显示,喷施不同质量浓度铜硼肥对忍冬藤中Fe3+、Cu2+、Mn2+、Zn2+和B3+含量均有显著影响,各处理均较对照有不同程度提高,且增幅大多达到显著水平(P<0.05)。

表4 不同质量浓度铜硼肥处理下矿质离子含量的变化
其中,在单施硼肥处理时,忍冬藤中Fe3+、Mn2+和B3+含量随着喷施B3+质量浓度的增加呈现先上升后下降的趋势,均在B40处理下最高,分别是对照的8.31倍、14.58倍和6.73倍;Cu2+含量却呈现持续上升趋势,并在B60处理时最高(0.058 mg/kg),较CK显著增加了544.44%;而Zn2+含量呈现下降趋势,但均显著高于对照,最高的B20处理(0.576 mg/kg)较CK显著增加了333.08%。
在单施铜肥处理时,忍冬藤中Mn2+和Zn2+含量随着喷施Cu2+质量浓度的增加均呈现先上升后下降的趋势,最高的Cu30处理分别较CK分别显著增加了1 144.81%和362.41%;而其Fe3+和B3+含量却呈现下降趋势,但Fe3+含量在各Cu2+质量浓度处理间无显著差异,B3+含量却均差异显著;Cu2+含量呈现上升趋势,处理间有显著差异。在铜硼肥配施时,Fe3+含量在B20Cu10、B20Cu30和B20Cu50处理,Cu2+含量在B20Cu50和B40Cu50处理,Mn2+含量在B20Cu30处理,Zn2+含量在B20Cu10、B20Cu30和B40Cu10处理,B3+含量在B20Cu10和B20Cu30处理下具有较高值,并显著高于其余配施处理;B20Cu30和B20Cu50配施处理最有利于提高忍冬藤中矿质离子Fe3+、Zn2+、B3+、Cu2+和Mn2+含量。
3 讨 论
3.1 铜硼肥对忍冬叶片光合生理特征的影响
微肥为植物生长发育提供必需的营养元素,对植物生长有重要影响。本研究发现喷施一定质量浓度的铜硼肥有助于提高叶片的净光合速率,尤以B40Cu50配施处理最佳。在一定范围内,单施铜肥或硼肥对植物的光合作用有促进作用,但过高的铜肥或硼肥则起抑制作用,这与刘海琴[13]对芍药的研究结论相似。单施铜肥或硼肥处理总叶绿素含量都是随铜硼肥质量浓度先升高再降低,硼、铜过量时叶绿体类囊体基粒片层明显减少,类囊体膜受到破坏而解体,叶绿素含量减少,活性降低,叶肉组织坏死,光合叶面积降低等[14-15],从而导致叶片中总叶绿素含量降低,这与本试验的研究结果一致,但总体上与对照并无显著差异,这可能与选取的4年生忍冬属于小型灌木,其生长发育并不容易发生显著变化有关。
3.2 铜硼肥对忍冬茎藤中营养物质和药效成分含量的影响
碳、氮代谢是植物体内重要的代谢途径,调控不同时期的养分供应水平,决定植物生长发育[16];两者存在着代谢和能量的竞争关系,因此调节二者之间的关系,使其协调分配,这对于植物体内碳、氮物质代谢平衡以及药材产量和品质的形成有着十分重要的意义。本研究中,忍冬藤中可溶性蛋白含量随着硼质量浓度增大而增大,在硼质量浓度达60 mg/L时得到显著提高。一定质量浓度的铜肥对忍冬的生理代谢有促进作用,是植物生长发育过程中不可或缺的微量元素,但过量的铜会对植物产生毒害。当铜肥质量浓度达到50 mg/L时,忍冬藤中的可溶性蛋白含量明显提高。有研究[17]表明,小麦等遇到铜胁迫时籽粒中可溶性蛋白含量升高,这与本研究结果一致。同时,铜硼配施后忍冬藤中可溶性糖含量较CK普遍增加,说明铜硼肥处理对忍冬体内可溶性糖积累有促进作用。另外,配施铜硼肥处理后忍冬藤中淀粉含量和还原糖含量较CK有不同程度的增加。充足的B和Cu对维持生菜、茼蒿叶片功能起着重要作用,缺B和Cu会使植物体内叶绿素含量下降,叶绿体结构遭到破坏,气孔关闭,导致光合作用产物减少,影响碳水化合物的合成[18],这与本试验的研究结果一致。但相较于可溶性糖和淀粉,忍冬藤中还原糖含量的增加较显著,这与崔志伟等的试验结果[7]一致,这可能是由于适宜铜硼不仅有利于可溶性糖和淀粉的运输,还有利于将多糖水解成单糖或二糖造成的。
绿原酸、马钱苷与总黄酮是忍冬藤的主要次生代谢产物,三者含量的高低是评价忍冬藤药用质量的重要标准。Campa等[19]认为咖啡中的绿原酸主要有2条合成途径,即莽草酸和咖啡酸途径。在绿原酸合成途径中涉及到的关键酶苯丙氨酸解氨酶(PAL),其活性在烟草植株中缺B元素时则会受到影响,进而影响体内绿原酸的合成[20]。因此喷施硼肥必然会对绿原酸以及黄酮类物质合成产生影响。研究发现适宜的铜硼配施质量浓度可以有效提高忍冬藤中绿原酸、马钱苷和总黄酮的含量。相较于营养物质,忍冬藤中的药效成分含量增加较显著,这可能是由于营养物质为忍冬藤中药效成分的合成提供了丰富的物质基础造成的。 关于铜、硼肥对忍冬藤中马钱苷、绿原酸和黄酮合成代谢的影响机制,可能与铜硼肥参与调控碳、氮代谢有关,铜硼肥不仅促进营养物质的合成,为药效成分的增加提供了丰富的物质基础,而且铜硼肥可能通过调节药效成分合成代谢过程中关键酶如裂环马钱苷合成酶(SLS)、PAL和黄酮合酶(FNS)等酶活性来调控马钱苷、绿原酸和黄酮的代谢途径以及含量,但其具体的作用机制还需进一步研究。
3.3 铜硼肥对忍冬藤微量元素含量的影响
矿质离子是植物新陈代谢必需的营养元素,是高等绿色植物生长过程中可维系其正常生长、促进其新陈代谢的无机营养元素[21]。本研究发现,单施与配施铜硼肥,忍冬藤矿质离子含量均有显著增加,其中配施铜硼肥比单施铜硼肥增加效果显著,尤以B20Cu10、B20Cu30和B20Cu50处理最佳。公勤等[22]研究表明低质量浓度(50 mg/kg)Cu2+处理显著增加了菠菜体内的Cu2+含量,但是植物能够主动调节抗氧化系统抵御Cu2+胁迫伤害,促进了矿质元素吸收;而高质量浓度(1 000 mg/kg)Cu2+胁迫导致菠菜体内Cu2+含量平均增至对照的105倍,此时植物通过调节体内抗氧化系统已很难抵御高质量浓度Cu2+胁迫伤害,导致矿质元素吸收受到显著抑制。本试验中Cu2+处理质量浓度为低质量浓度水平,因此各处理与CK相比较均促进了忍冬藤矿质离子的吸收,与前人的试验结果相一致。
4 结 论
喷施适宜质量浓度的铜、硼肥有利于提高忍冬叶的叶绿素含量及净光合速率,从而促进其生长,其中B40处理叶绿素含量最高(2.30 mg/g)。喷施适宜质量浓度的铜、硼肥还增加了忍冬藤营养物质含量,从而提高其品质,尤以B40Cu50处理组合最佳,其可溶性糖、淀粉、还原糖含量最高,分别为11.09 mg/g、392.46 mg/g和1.09 mg/g。铜硼肥处理也可显著提高忍冬藤的药效成分含量,其中绿原酸含量以B60Cu50组合处理最高(0.289%),B60Cu30(0.256%)和B40Cu50(0.255%)处理次之;马钱苷含量以B20Cu30组合处理最高(0.321%),B40Cu50(0.290%)和B60Cu50(0.292%)次之。此外,在单施与配施铜硼肥条件下,忍冬藤矿质离子含量均有明显增加,其中配施铜硼肥比单施铜硼肥增加效果显著,尤以B20Cu10、B20Cu30和B20Cu50处理组合最佳。综合以上分析,B40Cu50配施比例的忍冬叶片生理生化指标及其茎藤营养、药效成分含量均高于其他处理,这也为合理利用铜硼肥、优化忍冬栽培技术和提高其药材品质提供了理论参考。
猜你喜欢
河南农业科学(2023年1期)2023-03-22 12:41:44
中国肿瘤外科杂志(2022年4期)2022-09-01 08:52:26
腐植酸(2021年2期)2021-12-04 04:27:17
云南农业(2021年5期)2021-06-19 03:00:14
东坡赤壁诗词(2020年5期)2020-11-06 07:33:43
浙江农业科学(2019年4期)2019-04-19 05:40:20
现代农村科技(2019年5期)2019-01-06 11:05:53
现代园艺(2017年13期)2018-01-19 02:28:12
中国中医药信息杂志(2018年11期)2018-01-05 11:12:04
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
