
西藏色季拉山暗针叶林苔藓多样性及林窗干扰的影响
2023-08-05 03:51:56马和平石玉龙赵文茵慕金勇李金航
热带亚热带植物学报 2023年4期
马和平, 石玉龙, 赵文茵, 慕金勇, 李金航
西藏色季拉山暗针叶林苔藓多样性及林窗干扰的影响
马和平1, 石玉龙2, 赵文茵1, 慕金勇1, 李金航1
(1. 西藏农牧学院高原生态研究所,西藏林芝高山森林生态系统国家野外科学观测研究站,西藏高原森林生态教育部重点实验室,西藏自治区高寒植被生态安全重点实验室,西藏 林芝 860000;2. 四川省雅安市林业局,四川 雅安 625015)
为了解西藏色季拉山暗针叶林苔藓多样性及林窗干扰的影响,在过去工作积累的基础上研究了色季拉山西坡5块样地内不同林内环境(林窗、林缘和林下)地面、腐木和树附生苔藓生物量特征。结果表明, 地面生单位面积苔藓植物生物量储量最高,平均910.10 g/m2,其次为腐木生(221.90 g/m2),树附生的最低(53.59 g/m2)。林窗地面生苔藓单位面积生物量最高,均值为360.47 g/m2,其次为林下(305.51 g/m2),最小为林缘(244.11 g/m2);林窗、林缘和林下间的地面生苔藓单位面积生物量差异显著(<0.05)。沿海拔梯度,林窗苔藓单位面积生物量表现出先增加后减少的趋势。因此,在西藏色季拉山暗针叶林内,林窗对地面生苔藓单位面积生物量有显著影响。
苔藓植物;林窗;生物量;西藏色季拉山
干扰是森林空间格局变化的主要驱动力之一,是生态学家关注的焦点[1]。干扰生态已成为当代生态学研究的活跃领域。一般认为,干扰并非由林分内部的变化产生,因而是一种异生现象。实际上林分本身对于特定类型的干扰具有诱发性,如林分内枯枝落叶等易燃物的积累可诱发火干扰。因此可认为一些干扰是异生现象,另一些干扰是自源现象[2]。各种干扰因子或单独作用于森林,或多个因子相互组合形成复合干扰,对森林群落的更新和持续发展产生重要影响。
在森林动态中,干扰是指破坏林分结构,使物理环境因子或资源有效性发生改变的相对独立的事件[3–4]。林窗是介于全光环境和林冠环境之间的结构,有利于先锋物种的更新,对于维持群落中的物种多样性十分重要[5–7]。林窗也是森林中最普遍的干扰形式,是森林群落演替的主要驱动力,对森林生态系统的恢复和保护具有不可或缺的作用[8]。它能显著影响森林的空气温度、水分以及诸多环境因子,有利于森林小气候的形成[9]。因此,研究林窗对物种丰富度的影响,有助于揭示群落物种多样性的维持机制。
植物物种多样性与地上生物量及环境因子的相互关系是环境生态系统的基础[10]。森林中苔藓植物是生物多样性的重要组成部分,也是增加森林结构复杂性的重要生物因素[11–12]。苔藓植物在森林生态系统中广泛存在,在涵养水分、养分循环、积累生物量及促进生物多样性方面具有重要作用[13–14]。长期以来,苔藓植物的生物量和多样性的变化用于监测环境变化和森林管理,揭示森林生态系统的干扰特征[15]。因此,了解高山苔藓生物量特征对我们认识高山森林生态系统在维持生物多样性以及研究森林干扰对植物的影响具有重要意义。
色季拉山西坡的苔藓植物多样性与森林小气候相关,它们在一定程度上对苔藓植物的多样性产生影响,如海拔升高,空气温度逐渐降低,苔藓植物有特殊的适应低温的机制,所以在一定温度下, 苔藓植物多样性会随着温度的降低而有所增加,但是在极端气温下,苔藓植物的多样性则会受到抑制。随着海拔的升高,大气降水量增加,导致土壤湿度增加,土壤中的水分向外挥发使苔藓植物周围的小生境中空气湿度增加,苔藓植物主要从周围的空气中吸收水分和营养物质,从而能够在更多的环境中生长,表现出生物多样性增加。
苔藓植物是西藏色季拉山西坡林内物种多样性的重要组成部分,在过去工作积累的基础上,在色季拉山西坡不同海拔梯度的急尖长苞冷杉(var.)林内选择5块样地,研究了林窗、林缘和林下的地面、腐木和树附生苔藓生物量,拟探讨苔藓生物量如何响应其多样性和林窗干扰,为认识该区域森林干扰对林内苔藓植被的影响具有理论意义,也为认识色季拉山森林生态过程提供原始资料。
1 材料和方法
1.1 研究区概况
色季拉山位于藏东南雅鲁藏布江大拐弯西北侧,属念青唐古拉余脉,位于29°35~29°57′ N、94° 25′~94°45′ E。山体海拔2 800 (3 000)~4 000 (4 300) m,为亚高山(高原)寒温带半湿润区,森林分布线4 300~ 4 500 m以上为高山(高原)寒带。受印度洋暖湿气流影响,该区年均降水量1 134 mm,蒸发量544.0 mm,雨季为6月—9月,占全年降水的90%。年均温6.5 ℃,最冷月气温0 ℃~2.8 ℃, 最热月气温11.5 ℃~18.2 ℃,无霜期180 d,相对湿度60%~80%。色季拉山区是藏东南主要林区之一,天然林绵亘。其森林和植被类型及其分布范围、生长状况随坡向、海拔构成的综合生态环境变化而有明显差异[16]。色季拉山西坡主要森林植被类型为山地温带暗针叶林,以急尖长苞冷杉为建群种,高海拔区域伴生树种还有薄毛海绵杜鹃常绿灌木,低海拔区域伴生有林芝云杉(var.)、川滇高山栎()混交林等,且急尖长苞冷杉为西坡树线群落的优势树种。在海拔3 880 m的急尖长苞冷杉样地,由于上世纪70年代发生过一次较大的风灾,形成了大面积林窗。苔藓植物也是重要的物种组成部分,在高海拔地区,由于温度常年处于低温状态,植物生长周期短,苔藓植物数量少,且种类少,盖度低,主要有硬叶拟白发藓()、错那曲尾藓()、泛生丝瓜藓()、青毛藓()、曲背藓()等。
1.2 苔藓样品采集
参照汪沁等[17]的方法,在海拔3 540~4 305 m内以苔藓植物为研究对象,选取林分、坡向、坡度和地势组成等相似的5块100 m×100 m的大样地, 每块大样地以林窗为中心, 在其东、南、西、北4个方向的林窗、林缘和林下3个生境中各设置样方(表1)。地面生苔藓设置50 cm×50 cm样方,腐木和树附生苔藓样方均为20 cm×20 cm。2020年8月—9月进行苔藓调查采集,植株地上部分连同假根一起采集,同时记录样地的基本信息,如海拔、经纬度;调查样方内所有的苔藓物种,测量并记录苔藓在样方中的盖度和高度,用划分为100个5 cm边长小方格的样方框覆盖调查样方,观察每种苔藓植物所占方格数,以此作为该种苔藓在所调查样方中的盖度,如无苔藓生长,则盖度记为0。之后将小样方内的所有苔藓全部采集,将其放置于信封中,标注采集地点、海拔、经纬度、采集人、采集时间及样方编号。考虑到野外工作的可操作性,本文只采集样地内直径≥30 cm急尖长苞冷杉树体上的苔藓;对于地面生苔藓,采集50 cm× 50 cm小样方内的全部苔藓并带回实验室;对于腐木生苔藓,从其末端分别选取3个20 cm×20 cm的小样方并进行苔藓的采集;对树附生苔藓而言,从树体基部开始,选择距地面0、80、120和150 cm部位,采集东、南、西、北4个方向上20 cm×20 cm的苔藓。地面、腐木和树附苔藓都存在于3个林内环境中。
将带回的样品进行解剖观察,结合《中国苔藓志》[18–24]和《西藏苔藓志》[25]进行植物鉴定。之后将苔藓样品在滤网中用清水清洗、过滤,直至苔藓样品不含杂质(草根、树叶、土壤颗粒及其他),置于80 ℃烘箱中烘干,用电子天平(精度0.001)称质量。所有样地内林窗、林缘和林下的地面生苔藓单位面积生物量相加取其平均值,得出海拔梯度对地面生苔藓单位面积生物量的影响。
1.3 数据分析
利用Excel 10和SPSS 20软件进行数据分析, 采用单因素方差分析进行多重比较(<0.05), 分析林窗-林下和地面生-腐木生单位面积苔藓生物量在不同海拔间的差异性。
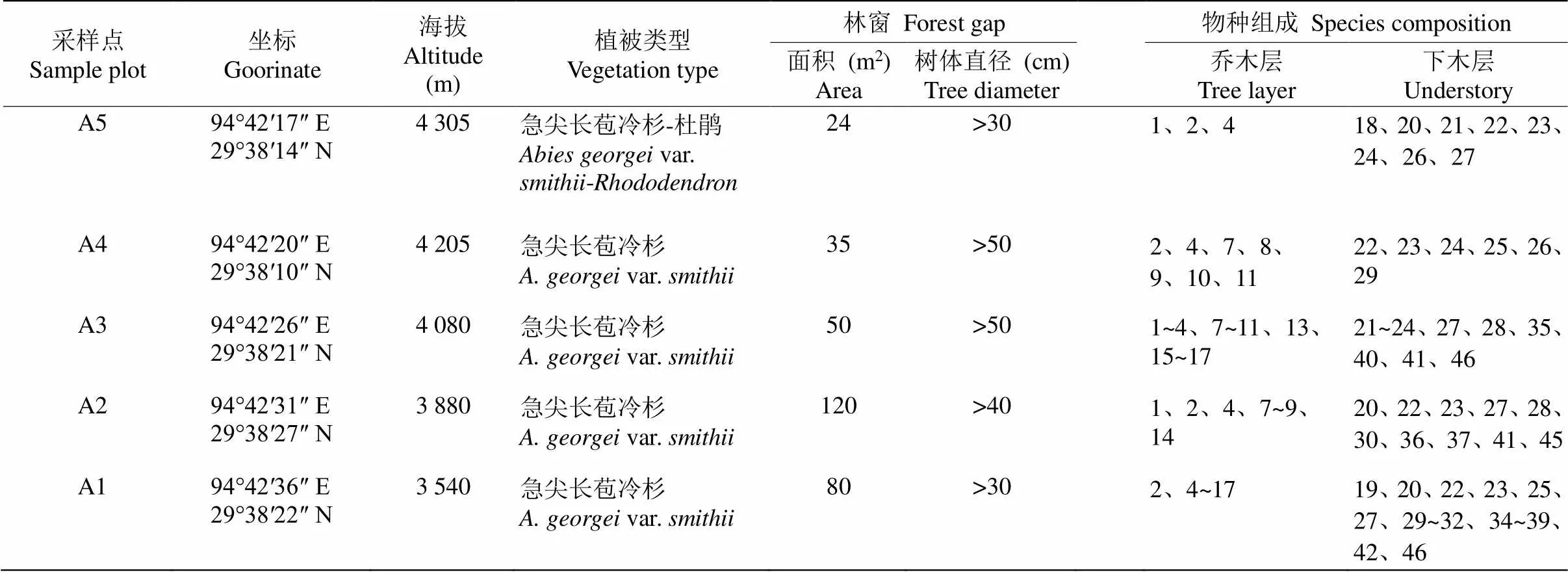
表1 调查样地基本情况
1: 红点杜鹃; 2: 薄毛海绵杜鹃; 3: 雪层杜鹃; 4: 急尖长苞冷杉; 5: 林芝云杉; 6: 糙皮桦; 7: 长尾槭; 8: 硬毛杜鹃; 9: 黄杯杜鹃; 10: 西南花楸; 11: 锈毛西南花楸; 12: 川滇高山栎; 13: 冰川茶藨子; 14: 陇塞忍冬; 15: 杯萼忍冬; 16: 柳叶忍冬; 17: 峨眉蔷薇;18: 长鞭红景天; 19: 长芽绣线菊; 20: 凉山悬钩子; 21: 扫帚岩须; 22: 宽叶苔草; 23: 直立悬钩子; 24: 岩白菜; 25: 长柱鹿药; 26: 西藏对叶兰; 27: 贡山肋毛蕨; 28: 小舌紫菀; 29: 香根芹; 30: 翅柄蓼; 31: 水栒子; 32: 无翅兔耳风; 33: 大叶山芥碎米荠; 34: 六叶箻; 35: 卷叶黄精; 36: 腋花扭柄花; 37: 总梗委陵菜; 38: 苇叶獐芽菜; 39: 假百合; 40: 小蓼; 41: 尾巴草; 42: 沼兰; 43: 西藏五福花; 44: 绒楚卫矛; 45: 荨麻叶凤仙花; 46: 五裂蟹甲草。
1:; 2:; 3:; 4:var.; 5:s var.; 6:; 7:; 8:; 9:; 10:; 11:var.; 12:; 13:; 14:; 15:; 16:; 17:; 18:; 19:; 20:; 21:; 22:; 23:; 24:; 25:; 26:; 27:; 28:; 29:; 30:; 31:; 32:; 33:var.; 34:; 35:; 36:; 37:; 38:; 39:; 40:; 41:; 42:; 43:; 44:var.; 45:; 46:.
2 结果和分析
2.1 苔藓植物多样性
对采集于色季拉山西坡的450份地面、腐木和树附生苔藓植物标本进行鉴定,共有14科45属78种。以种数≥6的科视为优势科,分别是真藓科(Bryaceae)、灰藓科(Hypnaceae)、丛藓科(Pottiaceae)、曲尾藓科(Dicranaceae)和青藓科(Brachytheciaceae)。地面生苔藓植物45属,共有4优势属,分别是丝瓜藓属()、青毛藓属()、青藓属()和棉藓属()。界定能在5个海拔样地中都出现的苔藓植物为该区域的优势种,分别是卷叶毛口藓()、明角长灰藓()、曲叶小锦藓()、泛生丝瓜藓()、错那曲尾藓()、赤茎小锦藓()、长蒴丝瓜藓()、曲背藓()、赤茎藓()、大灰藓(),隶属于6科8属。
2.2 苔藓生物量储量特征
西藏色季拉山西坡不同海拔梯度基质单位面积苔藓植物生物量储量见表2。地面生单位面积苔藓植物生物量储量最高,平均为910.10 g/m2,其次为腐木生(221.90 g/m2),树附生的最低,其平均值为53.59 g/m2。
对地面生苔藓植物而言,A2样地的单位面积生物量储量最高,达1 298.24 g/m2,A5样地最小; 对腐木生苔藓植物而言,A1样地的单位面积生物量储量最高(303.75 g/m2),A3样地最小;A1样地的树附生苔藓植物单位面积生物量储量最高(96.25 g/m2), A5样地的最小。
在本研究区,随着海拔的升高,温度降低, 苔藓盖度和高度显著降低,相应地其生物量也随之减少,尤其是在A5样地,由于地温、气温偏低,昼夜温差大,降雪早,并且半年以上覆盖有积雪,土壤长期处于冻结状态,再加上山高风大等综合因素的影响,逐渐不利于苔藓的孕育。加之该区长期处于低温,不易分解的凋落物(以杜鹃的枝和叶为主)覆盖了大部分地面,而过多的凋落物影响了苔藓的生长,致使其盖度降低,有些地方甚至不足5%。
2.3 林窗对地面生苔藓单位面积生物量的影响
从表3可见,林窗对地面生苔藓植物单位面积生物量储量有很大影响。A1样地内地面生苔藓植物单位面积生物量储量为林下>林缘>林窗,A2林下>林窗>林缘,A3和A4林窗>林下>林缘,A5林窗>林缘>林下,可见,不同海拔梯度下,林窗、林缘和林下苔藓植物单位面积生物量储量表现出无序性。
从表3可见,A1的变幅为林下>林缘>林窗, A2和A3林窗>林下>林缘,A4和A5林窗>林缘>林下,A2的变幅以林窗最大,为232.11 g/m2,林缘最小。A1的变异系数为林下>林缘>林窗,A2、A4为林窗>林缘>林下,A3为林下>林窗>林缘,A5为林窗>林下>林缘。
不同海拔均以林窗的地面生苔藓单位面积生物量最高,平均360.47 g/m2,其次为林下(305.51 g/m2),最小为林缘(244.11 g/m2)。林窗、林缘和林下间的地面生苔藓单位面积生物量差异显著(<0.05)。
2.4 海拔和林窗对苔藓单位面积生物量的影响
从图1可见,不同样地中林窗环境对苔藓单位面积生物量的影响不一致,沿海拔梯度林窗苔藓单位面积生物量呈先增加后减少的趋势。样地A1、A2、A4和A5间的林窗单位面积苔藓生物量储量差异不显著(>0.05),但A2和A4, A1和A5间的差异显著(<0.05)。A3与其他样地间均差异显著(<0.05)。A3和A4样地中,林窗与林缘和林下的差异显著(<0.05),而林缘和林下间的差异不显著(> 0.05);A5的林下与林窗和林缘差异显著(<0.05), 而林窗和林缘无显著差异(>0.05)。
3 结论和讨论
森林中苔藓植物的分布首先受到地理及气候等大尺度环境因子影响[26–27],而森林结构所决定的森林内部光、温度及湿度等小尺度环境因子对苔藓植物的分布也具有极大的影响,导致苔藓植物生物量在森林内的分布不同[28–31]。汪沁等[17]研究表明川西高山森林木生苔藓植物生物量随林窗位置的变化存在显著差异。马文章等[32]对热带、亚热带木生苔藓研究发现,边缘效应对苔藓植物的生长具有制约效应,从林内到林缘木生植物的生物量为降低趋势,这与我们的研究结果一致。

表3 色季拉山西坡地面生苔藓植物单位面积生物量特征值

图1 不同样地的地面生苔藓单位面积生物量。柱上不同大、小写字母分别表示相同海拔不同位置、不同海拔间的差异显著(P<0.05)。
本研究结果表明,在色季拉山的高海拔区域,如A5样地,地温、气温低,昼夜温差大,降雪早, 并且大半年有大量的积雪覆盖,土壤长期处于冻结状态,再加山高风大等综合因素的影响,严寒的胁迫使苔藓植物种类减少,环境逐渐不利于藓类植物的生长,除腐木生苔藓外,地面生和树附生苔藓单位面积生物量均最低,究其原因,可能是海拔的增加,温度逐渐降低,乔木分布受到抑制,海拔越高,植被越稀疏,紫外线照射越强烈,强烈的紫外线抑制了苔藓的生长,再加上这些区域长期处于低温状态,导致单位面积苔藓植物生物量较小。
林窗显著影响着苔藓植物生物量。如色季拉山西坡3 540~4 080 m区域,几乎是原始冷杉林,尤其是A2样地,林窗面积最大,在自然干扰下,林窗内灌木和草本稀少而地面生苔藓植物十分发达, 其盖度达到95%~100%,高度为20~40 cm,该处地面生苔藓单位面积生物量达到了最大值。A1样地林分郁闭度达75%,林内阴湿,腐木生和树附生苔藓植物丰富,故而以腐木生树附生苔藓单位面积生物量最大。
在色季拉山西坡,地面生单位面积苔藓植物生物量储量最高,其次为腐木生,树附生最低,生物量均呈先增加后降低的趋势。林窗对地面生苔藓单位面积生物量有显著影响。
[1] LIU H M, YANG Q S, FANG X F, et al. Influences on gap species richness in a subtropical evergreen broad-leaved forest [J]. Biodiv Sci, 2015, 23(2): 149–156. [刘何铭, 杨庆松, 方晓峰, 等. 亚热带常绿阔叶林林窗物种丰富度的影响因素 [J]. 生物多样性, 2015, 23(2): 149–156. doi: 10.17520/biods.2014137.]
[2] OLIVER C D, LARSON B C. Forest Stand Dynamics [M]. New York: McGraw-Hill, 1990: 1–140.
[3] WU Y G, LI H B. Theoretical development in landscape ecology [M]// LIU J G. Beijing: Science and Technology of China Press, 1992: 30–39.[伍业钢, 李哈滨. 景观生态学的理论发展 [M]// 刘建国. 当代生态学博论. 北京: 中国科学技术出版社, 1992: 30–39.]
[4] PICKETT S T A, WHITE P S. The Ecology of Natural Disturbance and Patch Dynamics [M]. New York: Academic, 1985: 472.
[5] LIANG X D, YE W H. Advances in study on forest gaps [J]. J Trop Subtrop Bot, 2001, 9(4): 355–364. [梁晓东, 叶万辉. 林窗研究进展[J]. 热带亚热带植物学报, 2001, 9(4): 355–364. doi: 10.3969/j.issn. 1005-3395.2001.4.015.]
[6] GARDNER R H, ENGELHARDT K A M. Spatial processes that maintain biodiversity in plant communities [J]. Persp Plant Ecol Evol Syst, 2008, 9(3/4): 211–228. doi: 10.1016/j.ppees.2007.08.003.
[7] BISWAS S R, MALLIK A U. Disturbance effects on species diversity and functional diversity in riparian and upland plant communities [J]. Ecology, 2010, 91(1): 28–35. doi: 10.1890/08-0887.1.
[8] LIU F, LI J J, TAN C, et al. Canopy gap characteristics and spatial patterns in subtropical broad-leaved forest [J]. Trans Chin Soc Agric Mach, 2020, 51(1): 230–236. [刘峰, 李建军, 谭畅, 等. 亚热带阔叶林林窗干扰特征与分布模式研究 [J]. 农业机械学报, 2020, 51(1): 230–236. doi: 10.6041/j.issn.1000-1298.2020.01.025.]
[9] LIU W J, LI Q J, ZHANG G M, et al. The microclimatic differences between and within canopy gaps in the dry-hot season inforest [J]. Acta Ecol Sin, 2000, 20(6): 932–937. [刘文杰, 李庆军, 张光明, 等. 西双版纳望天树林干热季不同林窗间的小气候差异 [J]. 生态学报, 2000, 20(6): 932–937. doi: 10.3321/j.issn:1000- 0933.2000.06.005.]
[10] GUO J X, YE M, YIN X K, et al. Spatial patterns of aboveground biomass and species diversity of grassland in the Two-river source region in Altay mountains [J]. J NW Agric For Univ (Nat Sci), 2022, 50(12): 87–96. [郭建兴, 叶茂, 殷锡凯, 等. 阿尔泰山两河源草地地上生物量和物种多样性的空间格局 [J]. 西北农林科技大学学报(自然科学版), 2022, 50(12): 87–96. doi: 10.13207/j.cnki.jnwafu.2022.12. 010.]
[11] LINDO Z, GONZALEZ A. The bryosphere: An integral and influential component of the earth’s biosphere [J]. Ecosystems, 2010, 13(4): 612– 627. doi: 10.1007/s10021-010-9336-3.
[12] TIAN W L, SUN S Q. Ecological functions of bryophyte: Recent research progress [J]. Chin J Ecol, 2011, 30(6): 1265–1269. [田维莉, 孙守琴. 苔藓植物生态功能研究新进展 [J]. 生态学杂志, 2011, 30(6): 1265–1269. doi: 10.13292/j.1000-4890.2011.0234.]
[13] WANG B, WU F Z, XIAO S, et al. Effect of succession gaps on the understory water-holding capacity in an over-mature alpine forest at the upper reaches of the Yangtze River [J]. Hydrol Processes, 2016, 30(5): 692–703. doi: 10.1002/hyp.10613.
[14] YANG L L, WANG Z S, ZHOU L Y, et al. Response and bioindicator of bryophyte and lichen as cryptogamae plants to environmental change [J]. J Nanjing For Univ (Nat Sci), 2012, 36(3): 137–143. doi: 10.3969/j.issn.1000-2006.2012.03.028. [杨琳璐, 王中生, 周灵燕, 等.苔藓和地衣对环境变化的响应和指示作用 [J]. 南京林业大学学报(自然科学版), 2012, 36(3): 137–143. doi: 10.3969/j.issn.1000-2006. 2012.03.028.]
[15] THOMAS S C, LIGUORI D A, HALPERN C B. Corticolous bryophytes in managed Douglas-fir forests: habitat differentiation and responses to thinning and fertilization [J]. Can J Bot, 2001, 79(8): 886– 896. doi: 10.1139/b01-066.
[16] XU F X. Appraising the quality and use of environment and resouse in Shergyla forest region, Tibet [J]. J Tibet Agric Anim Husb Coll, 1995(1): 1–8. [徐凤翔. 西藏色季拉林区环境、资源及质量与利用评价 [J]. 西藏农牧学院学报, 1995(1): 1–8.]
[17] WANG Q, YANG W Q, WU F Z, et al. Effects of forest gap and coarse woody debris on biomass and diversity of epixylous moss in an alpine forest [J]. Acta Ecol Sin, 2019, 39(18): 6651–6659. [汪沁, 杨万勤, 吴福忠, 等. 高山森林林窗和粗木质残体对木生苔藓生物量和多样性的影响 [J]. 生态学报, 2019, 39(18): 6651–6659. doi: 10.5846/stxb 201807171546.]
[18] GAO Q. Flora Bryophytarum Sinicorum, Vol. 1 [M]. Beijing: Science Press, 1994: 76–200. [高谦. 中国苔藓志, 第1卷 [M]. 北京: 科学出版社, 1994: 76–200.]
[19] LI X J. Flora Bryophytarum Sinicorum, Vol. 3 [M]. Beijing: Science Press, 2000: 70–94. [黎兴江. 中国苔藓志, 第3卷[M]. 北京: 科学出版社, 2000: 70–94.]
[20] LI X J. Flora Bryophytorum Sinicorum, Vol. 4 Eubryales [M]. Beijing: Science Press, 2006: 119–150. [黎兴江. 中国苔藓志, 第4卷真藓目 [M]. 北京: 科学出版社, 2006: 119–150.]
[21] WU P C, JIA Y. Flora Bryophytorum Sinicorum, Vol. 5 [M]. Beijing: Science Press, 2011: 173. [吴鹏程, 贾渝. 中国苔藓志, 第5卷[M]. 北京: 科学出版社, 2011: 173.]
[22] WU P C. Flora Bryophytorum Sinicorum, Vol. 6 [M]. Beijing: Science Press, 2002: 137. [吴鹏程. 中国苔藓志, 第6卷 [M]. 北京: 科学出版社, 2002: 137.]
[23] HU R L, WANG Y F. Flora Bryophytorum Sinicorum, Vol. 7 [M]. Beijing: Science Press, 2005: 260. [胡人亮, 王幼芳. 中国苔藓志, 第7卷 [M]. 北京: 科学出版社, 2005: 260.]
[24] WU P C, JIA Y. Flora Bryophytorum Sinicorum, Vol. 8 [M]. Beijing: Science Press, 2004: 254. [吴鹏程, 贾渝. 中国苔藓志, 第8卷 [M]. 北京: 科学出版社, 2004: 254.]
[25] Tibetan Plateau Comprehensive Scientific Expedition Team, Chinese Academy of Sciences. Tibetan Bryophyte Sinicorum [M]. Beijing:Science Press, 1985: 33–443. [中国科学院青藏高原综合科学考察队. 西藏苔藓植物志 [M]. 北京: 科学出版社, 1985: 33–443.]
[26] SCHMITT C K, SLACK N G. Host specificity of epiphytic lichens and bryophytes: A comparison of the Adirondack Mountains (New York) and the southern Blue Ridge Mountains (North Carolina) [J]. Bryologist, 1990, 93(3): 257–274. doi: 10.2307/3243509.
[27] PIIPPO S. Epiphytic bryophytes as climatic indicators in eastern Fennoscandia [J]. Acta Bot Fenn, 1982, 119: 1–39.
[28] TRYNOSKI S E, GLIME J M. Direction and height of bryophytes on four species of northern trees [J]. Bryologist, 1982, 85(3): 281300.doi: 10.2307/3243047.
[29] GONZÁLEZ-MANCEBO J M, ROMAGUERA F, LOSADA-LIMA A, et al. Epiphytic bryophytes growing on(Seub.) Franco in three laurel Forest areas in Tenerife (Canary Islands) [J]. Acta Oecol, 2004, 25(3): 159–167. doi: 10.1016/j.actao.2004.01.005.
[30] GUO S L, CAO T. Studies on community distributive patterns of epiphytic bryophytes in forest ecosystems in Changbai Mountain [J]. Acta Phytoecol Sin, 2000, 24(4): 442–450. [郭水良, 曹同. 长白山地区森林生态系统树附生苔藓植物群落分布格局研究 [J]. 植物生态学报, 2000, 24(4): 442–450.]
[31] GUO S L, CAO T. Studies on relationships of epiphytic bryophytes and environmental factors in forest ecosystems in Changbai Mountain forests [J]. Acta Ecol Sin,2000, 20(6): 922–931. [郭水良, 曹同. 长白山森林生态系统树附生苔藓植物分布与环境关系研究 [J]. 生态学报, 2000, 20(6): 922–931. doi: 10.3321/j.issn:1000-0933. 2000.06.004.]
[32] MA W Z, LIU W Y, YANG L P, et al. Edge effects on epiphytes in montane moist evergreen broad-leaved forest [J]. Biodiv Sci, 2008, 16(3): 245–254. [马文章, 刘文耀, 杨礼攀, 等. 边缘效应对山地湿性常绿阔叶林附生植物的影响 [J]. 生物多样性, 2008, 16(3): 245– 254. doi: 10.3321/j.issn:1005-0094.2008.03.006.]
Effects of Bryophytes Diversity and Gap in a Dark Coniferous Forest in Xizang Sygera Mountain
MA Heping1, SHI Yulong2, ZHAO Wenyin1, MU Jinyong1, LI Jinhang1
(1. Institute of Tibet Plateau Ecology, Agriculture & Animal Husbandry University, National Key Station of Field Scientific Observation & Experiment, Tibet Key Laboratory of Forest Ecology in Plateau Area, Key Laboratory of Alpine Vegetation Ecological Security,Nyingchi 860000, Tibet, China; 2. Ya’an Forestry Bureau,Ya’an 625015, Sichuan, China)
In order to understand the diversity of bryophytes and their response to gap disturbance in dark coniferous forests in Xizang Sygera Mountain, the biomass characteristics of epiphytic bryophytes on the ground, rotting wood and tree in different habitats (forest gap, forest edge, understory) were studied in five plots of western slope in Xizang Sygera Mountains. The results showed that the biomass per unit area of ground bryophytes were the highest with an average of 910.10 g/m2, followed by saprophytic bryophytes (221.90 g/m2) and epiphytes (53.59 g/m2). The biomass per unit area of ground moss in the forest gap was the highest with an average of 360.47 g/m2, followed by understory (305.51 g/m2), and forest edge (244.11 g/m2). There was a significant difference in the biomass per unit area among three habitats (<0.05). Along altitude gradients, the biomass per unit area of bryophytes grown in forest gap increased at first and then decreased. Therefore, it was suggested that the forest gap had a significant effect on the biomass per unit area of ground moss in dark coniferous forest in Xizang Sygera Mountain.
Bryophytes; Forest gap; Biomass; Xizang Sygera Mountain
10.11926/jtsb.4655
2022-04-19
2022-07-04
中国科学院植物研究所植被与环境变化国家重点实验室开放课题(LVEC-2020kf01)资助
This work was supported by the Open Project of State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences (Grant No. LVEC-2020kf01).
马和平(1977年生), 男, 博士, 教授,主要从事苔藓植物生态学的相关研究。E-mail: 285477889@qq.com
猜你喜欢
疯狂英语·新读写(2023年10期)2023-11-22 03:31:30
西南林业大学学报(2022年5期)2022-10-17 09:56:38
军事文摘(2021年18期)2021-12-02 01:28:36
资源导刊(2021年9期)2021-10-02 13:05:15
农业机械学报(2020年1期)2020-03-13 00:27:38
现代农业科技(2019年21期)2019-12-25 01:17:41
儿童故事画报·自然探秘(2017年7期)2018-03-14 19:05:32
西部林业科学(2018年4期)2018-02-14 22:43:50
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:56
西南林业大学学报(2014年6期)2014-07-24 19:02:11
