
浙江九龙山香果树群落乔木层物种的多样性格局
2023-08-05 03:51:54郑子洪郭小华包小梅唐战胜周世旺骆争荣
热带亚热带植物学报 2023年4期
郑子洪, 郭小华, 包小梅, 唐战胜, 周世旺, 骆争荣
浙江九龙山香果树群落乔木层物种的多样性格局
郑子洪1, 郭小华1, 包小梅2, 唐战胜1, 周世旺1, 骆争荣3*
(1. 浙江九龙山国家级自然保护区管理中心,浙江 遂昌 323300; 2. 遂昌县生态林业发展中心,浙江 遂昌 323300;3. 丽水学院生态学院,浙江 丽水 323000)
为了解香果树()群落的物种多样性及其驱动因素,在浙江九龙山自然保护区建立了35个以香果树为中心15 m半径的样圆,调查样圆内胸径2.5 cm以上的乔木层树木的物种和胸径。通过线性混合效应模型分析多样性与海拔、坡向、香果树胸高断面积的关系,通过Mantel检验和方差分解分析样地间距离、海拔、坡向和香果树胸高断面积差异对多样性的影响。结果表明,九龙山香果树群落乔木层物种丰富(50科96属145种),且以落叶或半常绿树种占优势; 样地乔木层Shannon-Wiener指数为2.37~3.40,Simpson指数为0.86~0.94;群落乔木层多样性随海拔升高呈先升高后下降的变化趋势,但与样地坡向和香果树胸高断面积无关;群落乔木层Sorenson指数为0.15~0.95,物种周转组分对群落多样性的贡献达74.88%;样地间地理距离与乔木层多样性及其组分呈显著正相关,样地间海拔差异与其乔木层多样性及其物种丰富度差异组分呈显著正相关,而样地间香果树胸高断面积差异仅与物种周转组分显著正相关;样地地理距离对于乔木层多样性及其组分具有最高的解释度(23%~29%),海拔差异对于乔木层多样性也有较高的解释度(16%)。因此,九龙山香果树群落乔木层的多样性主要受海拔影响,而多样性及其组分受扩散限制、生境过滤或香果树-伴生种互作等因素的影响。
多样性;多样性; 珍稀植物; 海拔; 空间距离
香果树()为茜草科(Rubi- aceae)香果树属高大落叶乔木,是我国特有的第四纪孑遗植物,在我国的分布相对较广,陕西南部和甘肃南部、华中、华东、华南和西南地区15个省区都有分布[1–2]。香果树喜生于海拔430~1 630 m处岩石裸露度高、土层浅薄的山谷林中[1–3],在次生林和原生林中都有香果树分布[1,3]。在其自然分布区,香果树可以形成纯林,也可以与其他植物共生组成混交林[4–5]。由于人类的毁林开荒和乱砍滥伐,生境片段化加剧,香果树的生境受到了极大的破坏,加上其种子和幼苗在野外存活率低,天然更新能力差,存在较大灭绝风险,从1999年开始香果树就被列为国家Ⅱ级重点保护植物[2–5]。加强对香果树的保护生物学研究对于该物种的保护和利用具有重要的意义。
过去的几十年间,我国学者从香果树的传粉生物学、种子萌发、生理特性、解剖结构、物候特征、种群动态及遗传多样性、群落结构和繁育技术等方面对于香果树进行了广泛的研究,取得了重要的研究成果,但在群落方面的研究依然相对薄弱[6]。群落物种多样性能够体现群落的结构性、稳定性及演替过程,通常多样性高的群落的稳定性和抗干扰能力强[2]。群落多样性主要包括多样性、多样性、多样性3个方面,其中多样性衡量了一个群落或群落所处环境内部的物种多样性;多样性是指在某个环境梯度上,不同群落之间物种组成上的相异性,即从一个生境到另一个生境所发生的物种多样性变化的速率和范围[7]。研究多样性有助于阐明群落内和群落间异质性的大小及其对物种的影响[8]。然而,目前国内外学者对香果树群落物种多样性的研究并不多,且仅限于多样性的描述。国内针对珍稀植物群落多样性的研究非常少,且缺少对多样性格局形成机制的解析及其对珍稀植物影响的分析[9–10]。已有的调查结果表明,香果树所在植物群落的物种多样性大多较低[5,11–16],但也有较高的[17]。张明月等[18]的研究发现即使在同一地区,香果树群落的物种多样性也会表现出较大的差异。香果树群落多样性变化的规律及其背后的生态学机制至今未得到充分研究。群落物种多样性受多方面因素影响,如区域种库的大小、演替、环境梯度、生境异质性、生物间相互作用、取样面积等[19–20]。已有的研究表明演替阶段对香果树群落的多样性具有重要影响,原生林香果树群落的物种多样性明显大于次生林香果树群落的物种多样性[2]。在大盘山自然保护区的香果树群落中,香果树种内竞争大于其与伴生种的种间竞争,香果树与杉木()等伴生乔木间具有较高的生态位重叠和竞争强度[21]。然而,群落中香果树种群与伴生种之间的关系如何影响群落物种多样性仍未得到深入研究。现有研究并未揭示香果树种群特征(如胸高断面积)和群落物种多样性的关系。另外,在影响植物群落和多样性的众多因子中,海拔和坡向被认为是具基础作用的因子[22–23],但仍不清楚这些基础因子对于香果树群落多样性的影响。
本研究以浙江九龙山分布的香果树群落乔木层(胸径大于2.5 cm的乔、灌木)为研究对象,通过广泛的样地调查,探究不同香果树群落乔木层的和多样性随海拔和坡向的变化规律,同时分析香果树群落乔木层多样性与香果树在样地中的胸高断面积的关系,以期为香果树群落和种群的合理保护提供理论依据。
1 研究区概况
浙江九龙山国家级自然保护区位于浙、闽、赣交界处的丽水遂昌西南部,位于28°19′10″~28°24′ 43″ N、118°49′38″~118°55′03″ E。九龙山自然条件优越,在海拔千米以上的山地保留有接近万亩的原始状态的天然林。该地区的气候属中亚热带湿润季风气候,年均温约17 ℃,最冷月为1月,极端最低温–17℃,最热月为7月,极端最高温37 ℃;年均降水量约1 855 mm,雨量充足,年均相对湿度83%;四季特征明显,夏季高温多雨,冬季寒冷潮湿;因其地形复杂,九龙山保护区的气候具有垂直地带性,山顶部风大气候变化复杂、高海拔区域气候近似温带,南北坡气候存在差异。土壤属富铝土纲, 红壤土类,包括老红壤、红壤、黄红壤和红黄壤等亚类。随着海拔的变化,各亚类按序替代。土壤中有机质、全氮及钾素丰富,有机质转化迅速,以多水高岭土和三水铝石为主,腐殖质组成以富里酸占优势[24]。该保护区植物物种丰富,是香果树在浙江的主要集中分布地之一[25];植被以亚热带常绿阔叶林最为典型,分布最为广泛,也是华东地区原始植被保持最优良的地区之一[24]。
2 研究方法
2.1 样地设置与调查
2021年7月中旬至10月初,在九龙山国家级自然保护区内的大岩前、岩背坑、陈坑、源大坑、内阴坑、内北坪等6处香果树分布地共设置35个固定样地。按照植被调查取样的一般原则以及对样地面积的要求[26],本研究以香果树为圆心,15 m为半径建立样圆作为调查样地(面积706.5 m2)。通过中心树的选择尽量让同一分布点的香果树都被圈入样圆,纳入长期监测。利用手机两步路APP测量每个样地的海拔和地理坐标,判断样地所处的坡向(分为东、南、西、北、东南、西南、东北、西北8个方向)。用塑料绳拉出样地范围后,给样地所有胸径2.5 cm以上的乔、灌木挂上(用铁钉钉或用不锈钢丝绑)带有唯一编号的铝牌,鉴定物种名,估测树高,用游标卡尺(树木胸径小于5 cm时使用)或胸径尺(树木胸径大于5 cm时使用)测量胸径(DBH),并用莱卡手持激光测距仪测量每棵符合标准的乔、灌木到中心香果树的水平距离[17,23], 记录所有数据。树木在1.3 m高度以下有分叉的,测量并记录各个分叉的胸径。
2.2 数据统计分析
2.2.1 物种分类与重要值计算
物种学名和分类以为准,叶的生长习性(常绿、半常绿和落叶)根据《浙江植物志》的记载。用重要值表示香果树及其伴生种在群落中的重要程度[26],VI=(rD+rBA+rFR)/3,其中,VI表示物种的重要值,物种的相对密度(rD)为该物种的密度除以所有物种的密度之和,物种的相对显著度(rBA)为该物种的所有个体(包括分叉)的胸高断面积之和除以所有物种所有个体的胸高断面积之和,物种的相对频度(rFR)为出现该物种的样圆数除以各物种出现的样圆数之和。
2.2.2 群落多样性分析
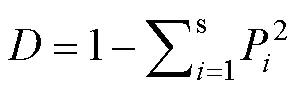
为了分析不同样地乔木层间的多样性的差异与海拔、坡向等地形因子以及样地中香果树种群特征的关系,本研究以各样地乔木层的和指数作为应变量,样地的海拔、坡向、香果树种群的胸高断面作为自变量(固定效应)进行回归分析。考虑到多样性随海拔升高可能存在先升高后降低或先降低后升高的现象,本研究同时将海拔和海拔的二次方作为自变量考虑。由于有时在同一香果树分布地(如大岩前)设置的样地距离较近,这些样地的多样性本身可能存在较强的空间自相关。为了避免多样性指数空间自相关对结果的显著性检验造成影响,本研究根据香果树分布地将35个样圆分成6组,作为影响截距的随机效应加入回归方程,建立线性混合效应模型。利用检验对各自变量固定效应的显著性进行检验。
2.2.3 群落多样性分析
当代学者普遍认为,多样性格局来源于物种替换和物种得失两个方面,前者导致群落间物种丰富度的差异,后者导致群落间共有种减少及各自独有种增加[28–29]。本研究采用基于多度的Sorenson相异性指数来作为配对样方乔木层之间的多样性(即群落之间的物种组成的差异,total)的度量, 并将多样性分解为物种周转组分(epl)和物种丰富度的差异组分(diff)[23]。total=(+)/(2++),epl=2min(,)/(2++),Diff=|–|/(2++),其中表示两个配对样地中每种植物在两个样地间的最小多度的总和;表示两个配对样地中样地1的总多度与的差值;表示两个配对样地中样地2的总多度与的差值。物种丰富度差异组分和物种周转组分所占比分别是Diff/total与epl/total。当Diff/total>0.5时,表明多样性主要由物种丰富度差异组分主导,epl/total>0.5时,表明多样性主要由周转组分占主导[30]。
用Spearman秩相关系数计算样地间地理距离与香果树群落乔木层多样性及其组分的相关性,通过Mantel检验分析这些相关系数是否显著。利用Spearman秩相关系数和偏Mantel检验分析控制样地间地理距离(协变量)条件下,其它因子(如样地的海拔、坡向和样地内香果树胸高断面积)对香果树群落乔木层多样性及其组分的单独影响[23,30]。然后,采用基于距离的冗余分析(dbRDA)的方差分解来探究上述分析得到的对九龙山香果树群落乔木层多样性及其组分有显著影响的因素的相对重要性[23]。方差分解分析中,响应变量是九龙山香果树群落乔木层多样性及其2个组分的3个距离矩阵,解释变量为经Mantel检验得到的对香果树群落乔木层多样性有显著影响的海拔、坡向、样地内香果树胸高断面积或样地间的空间距离等因素。空间距离因子由邻体矩阵主坐标分析(PCNM)引入,即将样地间空间距离矩阵经PCNM分析后,选择正相关的PCNM特征值作为空间因子指标。该方法可以有效获取样方间的空间结构,常用于分析群落空间变化的解释变量[31]。
2.2.4 数据分析
所有数据分析都在R 4.2.1软件中完成。Shannon-Weiner指数和Simpson指数计算采用vegan 2.6-2程序包[32]中的diversity函数实现,线性混合效应模型分析通过lme4 1.1-29程序包[33]中的lmer函数实现。多样性分解分析通过R 4.2.1软件的adespatial 0.3-16程序包[34]的beta.div.comp函数实现,Mantel和偏Mantel检验分别通过vegan 2.6.-2程序包中的mantel和meantel.partial函数实现,PCNM分析和方差分解分别通过vegan 2.6-2程序包中pcnm和varpart函数实现。
3 结果和分析
3.1 群落的物种组成
调查的35个香果树样地乔木层共有木本维管束植物50科96属145种,其中裸子植物3科4属4种,被子植物47科92属141种。145种植物中常绿树种56种,占总种数的38.62%,落叶或半常绿树种89种,占总种数的61.38%。含5种及以上的有8科,分别为壳斗科(Fagaceae,11种)、山茶科(Theaceae, 10种)、樟科(Lauraceae, 9)、杜鹃花科(Eric- aceae, 9种)、蔷薇科(Rosaceae, 8种)、忍冬科(Capri- foliaceae, 6种)、冬青科(Aquifoliaceae, 5种)、槭树科(Aceraceae, 5种)。
乔木层重要值>2的优势物种有杉木(10.31)、香果树(5.31)、毛脉槭(, 5.14)、川鄂山茱萸(, 3.92)、缺萼枫香(, 3.91)、木荷(, 3.82)、浙闽樱(, 2.87)、格药柃(, 2.70)、青榨槭(, 2.54)、褐叶青冈(, 2.42)、建润楠(, 2.10)、柳杉(, 2.03)。从科的重要值(该科所有种的重要值之和)来看,九龙山香果树群落乔木层的优势科主要有杉科(Taxodi- aceae,=12.35)、山茶科(10.84)、槭树科(8.28)、山茱萸科(Cornaceae, 7.25)、蔷薇科(6.40)、茜草科(5.42)、壳斗科(5.37)、樟科(5.22)。九龙山香果树群落乔木层落叶或半常绿树种重要值之和达59.48, 而常绿树种为40.52。落叶树种在群落中明显占优势。
3.2 群落α多样性及其变化
九龙山香果树群落乔木层的Shannon-Wiener指数为2.37~3.40,均值为2.88;Simpson指数为0.86~0.94,均值0.92。线性混合效应模型分析表明坡向和样地内香果树胸高断面积对香果树群落乔木层的Shannon-Wiener指数和Simpson指数的等效物种数(即和)无显著影响;而海拔及其二次方对Shannon-Wiener指数和Simpson指数的等效物种数都有显著影响(<0.05),九龙山香果树群落乔木层的多样性随着海拔呈现出先升高后下降的趋势(图1)。根据模型估计,Shannon-Wiener指数等效物种数在海拔1 175 m处达到最高,而Simpson指数等效物种数在海拔1 200 m处达到最高。

图1 香果树群落乔木层Shannon-Wiener指数(H')和Simpson指数(D')随海拔的变化
3.3 群落β多样性组分特征及其影响因素
九龙山香果树群落乔木层多样性为0.15~ 0.95, 均值为0.70,其中物种周转组分为0.12~0.90,均值0.53,物种丰富度差异组分为0.00~0.54,均值0.18。2个组分中,物种周转组分(epl=0.53)对九龙山香果树群落乔木层多样性的贡献较高,达74.88%,而物种丰富度差异组分对于九龙山香果树群落乔木层多样性的贡献仅占25.12% (图2)。
Mantel检验结果表明,样地间地理距离与香果树群落乔木层多样性呈显著正相关(=0.74,< 0.01),即两块样地乔木层间物种组成差异随样地间空间距离的增大而增大。同时,空间地理距离与物种周转组分(=0.56,<0.01)和物种丰富度差异组分(=0.23,=0.02)之间都呈现出显著正相关。
偏Mantel检验结果表明,控制样地间地理距离矩阵之后,香果树群落乔木层多样性(=0.15,= 0.05)及其物种丰富度差异组分(=0.19,=0.02)与样地间香果树胸高断面积差异显著正相关,而其物种周转组分与样地间香果树胸高断面积差异无显著关系(=0.76)。偏Mantel检验表明,控制样地间地理距离矩阵之后,香果树群落乔木层多样性(= 0.30,<0.01)及其物种丰富度差异组分(=0.25,<0.01)与样地间海拔差异呈显著正相关,而物种周转组分与样地间海拔差异无显著关系(=0.12);香果树群落乔木层物种周转与样地间坡向差异呈显著正相关(0.21,<0.01),香果树群落乔木层多样性及其物种丰富度差异组分与样地间坡向差异无显著关系(>0.05)。
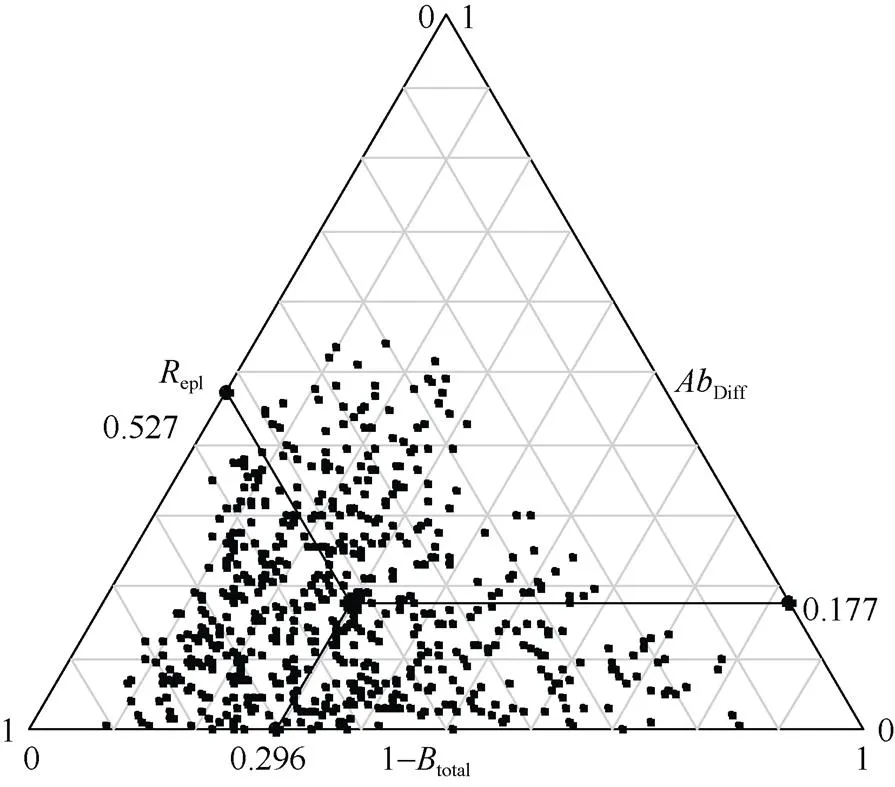
图2 果树群落乔木层β多样性(Btotal)及其物种周转组分(Repl)和物种丰富度差异组分(AbDiff)。黑点表示样方对,其位置由该样方对的物种组成相似性(1-Btotal)、物种周转组分和物种丰富度差异组分决定。黑色大点的位置表示所有小黑点的质心,位置由所有小黑点在3个轴上的均值决定。
方差分解分析结果表明,样地空间结构、海拔、坡向和样地香果树胸高断面积差异一共仅解释九龙山香果树群落乔木层多样性及其组分42%~49%的变异(图3)。其中样地的空间分布结构对于九龙山香果树群落乔木层多样性及其组分具有最高的解释度,单纯的空间结构分别解释23%的香果树群落乔木层多样性,以及23%物种丰富度差异和29%的物种周转。此外,海拔的空间结构解释了3%的香果树群落乔木层多样性;香果树胸高断面积的空间结构也解释了5%的物种丰富度差异;坡向的空间结构进一步解释了11%的群落物种周转。除了空间结构以外,海拔差异对于香果树群落乔木层多样性有较高的解释度,单纯的海拔差异因素可以解释16%的香果树群落乔木层多样性;而香果树胸高断面积差异对于群落物种丰富度差异有较高解释度,单纯的香果树胸高断面积差异可以解释16%的物种丰富度差异。单纯的坡向变化解释了9%的群落间物种周转。
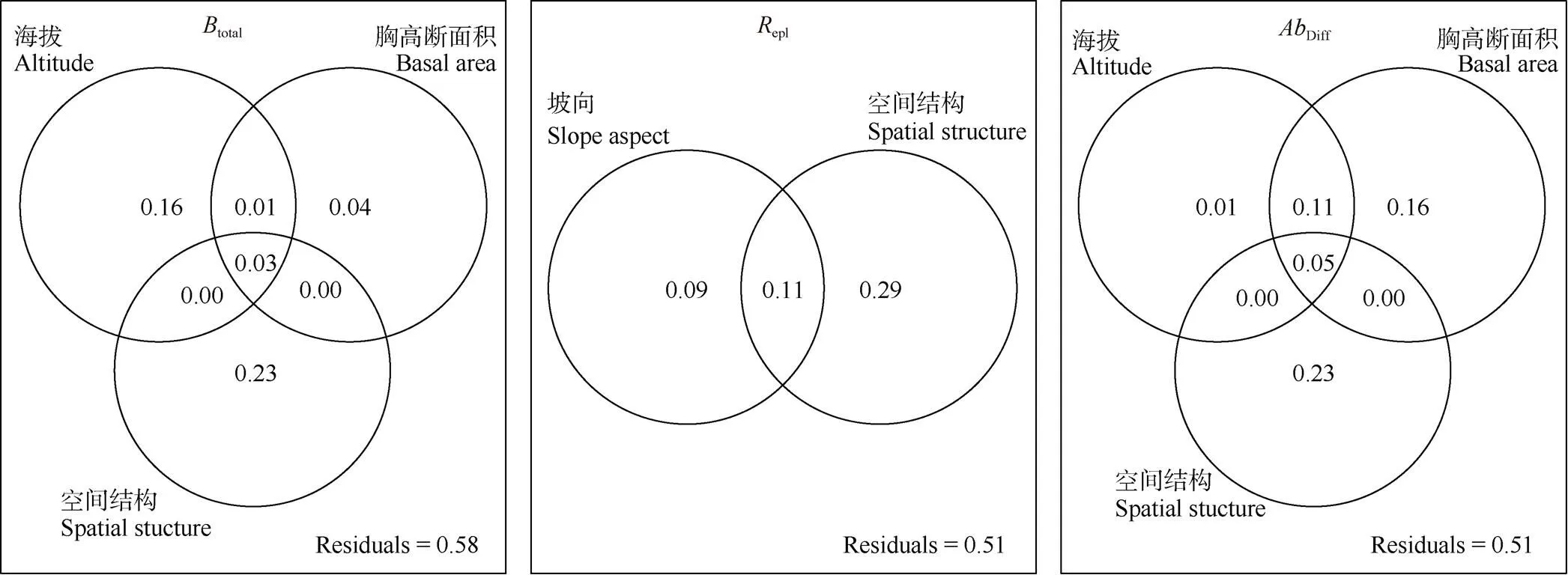
图3 海拔、坡向、空间分布结构或香果树胸高断面积差异对群落乔木层β多样性及其组分解释度的方差分解
4 结论和讨论
4.1 香果树群落的物种多样性
本研究在九龙山香果树群落乔木层中调查到木本维管束植物50科96属145种。由于来自各地的香果树群落调查的面积和起测胸径差异很大,精确比较全国各地香果树群落的多样性比较困难。从单个样地(706.5 m2)来看,九龙山香果树群落乔木层的Shannon-Wiener指数为2.37~3.40,Simpson指数为0.86~0.94。九龙山香果树群落乔木层的多样性高于浙江大盘山(2 000 m2,=1.84,=0.83)和江苏溧阳丘陵山地(800 m2,=2.05,=0.63)[5,15]。九龙山香果树群落乔木层的多样性处于中上水平。
整体来看,九龙山香果树群落乔木层种类最多的4科分别为壳斗科、山茶科、樟科、杜鹃花科, 其中前3科与古田山香果树群落种类最多的3科一致[17],九龙山和古田山地理距离较近,群落物种的主要科、属组成比较接近。然而,较高的分布海拔直接影响到九龙山香果树群落的物种组成,就古田山和九龙山香果树群落中重要值最大的12优势物种比较来看,仅有香果树、杉木和格药柃为两地香果树群落共有[17]。相较于常绿树种占优势的古田山香果树群落,九龙山香果树群落中落叶树种的重要性明显上升,落叶或半常绿树种重要值之和较常绿树种的重要值之和高18.96,槭树科、山茱萸科、蔷薇科等含有较多落叶树种的科的重要性提高。
4.2 香果树群落α多样性的影响因素
海拔是影响山地森林植物群落多样性的重要因素[7,19,22,35]。九龙山香果树群落主要分布在海拔965~1 447 m,明显高于浙江大盘山(740~910 m)、浙江古田山(612~851 m)、江苏溧阳丘陵山地(<541 m)、湖北九宫山(630 m)、安徽天堂寨(600~1 100 m)、河南桐柏山(300~900 m)等地的香果树分布的海拔高度[5,11–13,15–17]。本研究结果表明,九龙山香果树群落乔木层的多样性随着海拔呈现出显著的先升高后下降的趋势,约在海拔1 200 m处群落乔木层的多样性达到最高。由于在建立省级保护区之前,九龙山区多地都有农民居住或从事农林生产经营活动,低海拔地区受干扰更严重,因此较低海拔地区香果树群落的多样性较低。而高海拔地区的群落发育比较成熟,群落内树木经过激烈的种间和种内竞争,个体数相对较少,从而导致多样性下降。由于我国自然保护地在设立前普遍经历过人为干扰,群落物种多样性随海拔变化的这种趋势可能在全国普遍存在[22]。分布海拔适中可能也是九龙山香果树群落乔木层多样性高于其它地方的重要原因。
前人[11–13]普遍认为多样性低、稳定性差的群落不利于香果树的生存。然而,本研究表明香果树群落乔木层的多样性与香果树在样方中的总胸高断面积无关。这说明香果树的生长不影响其群落乔木层的多样性,同时群落乔木层多样性可能也不会影响香果树在群落中的生存。由于本研究仅用了一次调查的静态数据,该判断仍需进一步的动态研究加以证实。利用固定样地对香果树的开花、结实、幼苗萌发和存活进行长期动态监测,并分析这些参数与群落多样性的关系将有助于回答这一问题。
4.3 香果树群落β多样性的影响因素
研究群落多样性格局,解析其形成的驱动因素可以为探讨群落构建机制提供视角,也有助于了解珍稀植物与群落环境以及其他物种关系[8–10,20,22,31]。本研究的结果表明,九龙山香果树群落乔木层多样性较高,Sorenson相异性指数均值为0.70,这一结果说明香果树能够适应不同的生境和伴生物种。虽然香果树喜生于岩石裸露度高、土层浅薄的山谷林中,对小生境有一定要求,但该物种在我国15个省份分布,其分布海拔也从低海拔扩展到中高海拔[1–3]。这充分说明香果树对于不同的气候环境具有良好的适应性。九龙山香果树群落乔木层多样性的2个组分中,物种周转组分占比远大于物种丰富度差异组分,这说明九龙山香果树群落间的物种组成差异主要来源于样地间的物种替换,这与国内外很多研究的结果类似[23,30–31,36]。导致物种周转组分占比较高的原因较多,如生境异质性、竞争、扩散限制和优先效应等[20,23]。九龙山香果树群落间的乔木层物种周转随样地间空间距离的增大而显著增大,单纯的样地空间分布结构是解释九龙山香果树群落乔木层物种周转最主要的因素,这可能是由香果树伴生种的扩散限制和一些本研究未检测的环境因子(如土壤养分、微地形)的空间结构造成的[31]。除此之外,本研究分析表明坡向差异是香果树群落间乔木层物种周转的重要解释因子,九龙山南北坡的气候存在较大差异,一般南坡光照充足,相对干燥,而北坡较阴湿,这一气候差异导致了分布在九龙山保护区内不同坡向的香果树群落的物种替代。本研究结果表明单纯的空间分布结构解释了23%的香果树群落乔木层物种丰富度差异。这说明扩散限制和本研究未检测的环境因子的空间结构对香果树群落乔木层的伴生物种得失产生了重要影响。除空间因素外,样地间海拔的差异对香果树群落间乔木层伴生种的得失也有影响,但是其解释度并不高,单纯的海拔因素的生境过滤效应仅解释香果树群落乔木层1%的物种丰富度变化。
本研究结果表明香果树胸高断面差异显著提高香果树群落间乔木层物种组成(多样性)以及物种丰富度变化,但不影响物种周转。单纯的香果树胸高断面积变化解释16%的香果树群落乔木层伴生物种得失;同时,与海拔(以及空间结构)有关的香果树胸高断面积差异解释另外16%香果树群落乔木层伴生物种得失,这说明香果树与其他伴生种的种间互作对于香果树群落的物种得失具有十分重要的作用,且比上述扩散限制和其他环境因素的空间效应更重要。康华靖等[21]在浙江大盘山的研究表明,香果树与杉木间具有最高的生态位重叠值和最强的种间竞争,而与尖连蕊茶()间的生态位重叠和种间竞争相对较低。今后的研究应就香果树与伴生种互作如何影响香果树更新与死亡开展进一步分析。
致谢 感谢九龙山自然保护区潘成椿同志、丽水学院陈旭波和张亚芬老师,李文、柏正伟和郑蓉同学在野外调查过程中提供的支持和帮助。
[1] Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora Reipublicae Popularis Sinicae, Tomus 71 [M]. Beijing: Science Press, 1999: 242. [中国科学院中国植物志编辑委员会. 中国植物志, 第71卷 [M]. 北京: 科学出版社, 1999: 242.]
[2] PENG X L. Population dynamics and community diversity ofin primary and secondary forest [D]. Nanjing: Nanjing Normal University, 2018. [彭仙丽. 原生林与次生林中香果树的种群动态与群落多样性 [D]. 南京: 南京师范大学.]
[3] SHANG K K, CHEN B, ZHANG X J, et al. Topographic influences on the population persistence of a tertiary relict deciduous treeOliv. on Mt. Tianmu, eastern China [J]. Appl Ecol Environ Res, 2020, 18(5): 6053–6067. doi: 10.15666/aeer/1805_60536067.
[4] FANG Y P, XIANG J, HU Y, et al. Present situation ofresource protection in Dabieshan in eastern Hubei [J]. J Anhui Agric Sci, 2007, 35(6): 1756–1757. [方元平, 项俊, 胡扬, 等. 鄂东大别山区香果树资源现状及其保护 [J]. 安徽农业科学, 2007, 35(6): 1756–1757. doi: 10.3969/j.issn.0517-6611.2007.06.090.]
[5] CHEN Z L, KANG H J, LIU P, et al. Community structure features of(Rubiaceae) in Dapanshan National NaturalReserve of Zhejiang Province [J]. Acta Bot Yunnan, 2007, 29(4): 461– 466. [陈子林, 康华靖, 刘鹏, 等. 大盘山自然保护区香果树群落结构特征 [J]. 云南植物研究, 2007, 29(4): 461–466. doi: 10.3969/j.issn. 2095-0845.2007.04.016.]
[6] ZHENG Z H, ZHENG W C, ZHENG R, et al. Research progress on conservation biology of[J]. J Zhejiang For Sci Technol, 2022, 42(4): 94–101. [郑子洪, 郑伟成, 郑蓉, 等. 香果树保护生物学研究进展 [J]. 浙江林业科技, 2022, 42(4): 94–101. doi: 10.3969/j.issn.1001-3776.2022.04.015.]
[7] WHITTAKER R H. Vegetation of the Siskiyou Mountains, Oregon and California [J]. Ecol Monogr, 1960, 30(3): 279–338. doi: 10.2307/194 3563.
[8] MA K, LI Y N, JIN H, et al. A comparison of the species diversity ofcommunity in different habitats [J]. Pratac Sci, 2011, 28(8): 1467–1472. [马凯, 李永宁, 金辉, 等. 不同生境类型金莲花群落物种多样性比较 [J]. 草业科学, 2011, 28(8): 1467–1472.]
[9] HONG Y, ZHOU L L, ZHU Q F. Community structure characteristics of endangered plant[J]. Chin J Ecol, 2010, 29(10): 1907–1911. [红雨, 邹林林, 朱清芳. 珍稀濒危植物蒙古扁桃群落结构特征 [J]. 生态学杂志, 2010, 29(10): 1907–1911. doi: 10.13292/j. 1000-4890.2010.0264.]
[10] CHU W K, ZHOU Y Y, CHEN Z L, et al. On the community classi- fication and species diversity of[J]. J Hangzhou Norm Univ (Nat Sci), 2013, 12(3): 240–244. [褚文珂, 周莹莹, 陈子林, 等. 珍稀植物华顶杜鹃群落分类和物种多样性研究[J]. 杭州师范大学学报(自然科学版), 2013, 12(3): 240–244. doi: 10. 3969/j.issn.1674-232X.2013.03.010.]
[11] XU X Y, YAO C H, PAN J. Studies on structure features ofcommunity in the Jiugongshan Nature Reserve [J]. J SW For Coll, 2002, 22(1): 5–8. [徐小玉, 姚崇怀, 潘俊. 湖北九宫山香果树群落结构特征研究 [J]. 西南林学院学报, 2002, 22(1): 5–8. doi: 10.3969/j.issn.2095-1914.2002.01.002.]
[12] YANG K J, ZHANG X P, ZHANG Z X, et al. Analysis of actuality ofcommunity in Tiantangzhai of Anhui Province [J]. J Plant Resour Environ, 2007, 16(1): 79–80. [杨开军, 张小平, 张中信, 等. 安徽天堂寨保护植物香果树群落现状分析 [J]. 植物资源与环境学报, 2007, 16(1): 79–80. doi: 10.3969/j.issn.1674-7895. 2007.01.019.]
[13] MENG Q F, WANG M G, GAO H L, et al. Research on resources distribution and community frame ofOliv. of Tongbai Mountain in Henan Province [J]. Henan Sci, 2009, 27(1): 51– 54. [孟庆法, 王民庚, 高红莉, 等. 河南桐柏山香果树资源分布与群落结构研究 [J]. 河南科学, 2009, 27(1): 51–54. doi: 10.3969/j.issn. 1004-3918.2009.01.015.]
[14] LIU C Y, LIAO J H, CHEN Y H, et al. Structure characteristics and species diversity ofcommunity in Dawei Mountain of Hunan [J]. J CS Univ For Technol, 2011, 31(11): 110–113. [刘成一, 廖建华, 陈月华, 等. 湖南大围山香果树群落特征及物种多样性分析[J]. 中南林业科技大学学报, 2011, 31(11): 110–113. doi: 10.3969/j.issn.1673-923X.2011.11.022.]
[15] KONG L, ZHU Y, SHEN J J, et al. Community composition and species diversity ofcommunity in Liyang County of Jiangsu province [J]. J CS Univ For Technol, 2015, 35(3): 84–89. [孔磊, 朱莹, 沈静静, 等. 江苏溧阳香果树群落组成及物种多样性分析[J]. 中南林业科技大学学报, 2015, 35(3): 84–89. doi: 10.14067/j.cnki.1673-923x.2015.03.017.]
[16] PENG X L, LI L, ZHANG G F, et al. Species composition and diversity characteristics ofcommunities in five patches in the mountainous area of southern Jiangsu [J]. J Plant Resour Environ, 2017, 26(4): 93–100. [彭仙丽, 李莉, 张光富, 等. 苏南山区5个斑块香果树群落物种组成及多样性特征[J]. 植物资源与环境学报, 2017, 26(4): 93–100. doi: 10.3969/j.issn.1674-7895.2017.04.12.]
[17] LU W, TANG Z S, ZHENG Z J, et al. The community composition and structural characteristics offorest, an endangered species in Gutianshan, Zhejiang Province [J]. Ecol Environ Sci, 2018, 27(6): 1052–1059. [芦伟, 唐战胜, 郑振杰, 等. 浙江古田山濒危植物香果树群落组成和结构特征 [J]. 生态环境学报, 2018, 27(6): 1052–1059. doi: 10.16258/j.cnki.1674-5906.2018.06.008.]
[18] ZHANG M Y, LIU N N, LIU J, et al. Age structure and succession dynamic comparison ofpopulations in Dawei Mountain and Bamian Mountain, Hunan Province [J]. Acta Bot Boreali- Occid Sin, 2017, 37(8): 1603–1615. [张明月, 刘楠楠, 刘佳, 等. 湖南大围山和八面山香果树种群的年龄结构和演替动态比较 [J]. 西北植物学报, 2017, 37(8): 1603–1615. doi: 10.7606/j.issn.1000-4025. 2017.08.1603.]
[19] JEFFRIES M J. Biodiversity and Conservation [M]. London, New York: Routledge, 1997: 39–73.
[20] CHEN S B, OUYANG Z Y, XU W H, et al. A review of beta diversity studies [J]. Biodiv Sci, 2010, 18(4): 323–335. [陈圣宾, 欧阳志云, 徐卫华, 等. Beta多样性研究进展 [J]. 生物多样性, 2010, 18(4): 323– 335. doi: 10.3724/SP.J.1003.2010.323.]
[21] KANG H J, CHEN Z L, LIU P, et al. Intra-specific competition ofand its accompanying species in the Dapanshan National Nature Reserve of Zhejiang Province [J]. Acta Ecol Sin, 2008, 28(7): 3456–3463. [康华靖, 陈子林, 刘鹏, 等. 大盘山香果树()种内及其与常见伴生种之间的竞争关系 [J]. 生态学报, 2008, 28(7): 3456–3463.]
[22] JIN H, ZHAO Y, ZHAO W, et al. Elevational changes in species diversity and similarity analysis of acommunity on Changbai Mountain [J]. Acta Ecol Sin, 2015, 35(1): 125–133. [金慧, 赵莹, 赵伟, 等. 长白山牛皮杜鹃群落物种多样性的海拔梯度变化及相似性 [J]. 生态学报, 2015, 35(1): 125–133. doi: 10.5846/stxb201405120971.]
[23] SHE D Q, ZHANG X T, XIAO L, et al. Plant beta diversity and its influence factors in the Liangshui National Nature Reserve in the central region of the Xiaoxing’an Mountains [J]. Biodiv Sci, 2022, 30(3): 21274. [佘丹琦, 张喜亭, 肖路, 等. 小兴安岭凉水国家级自然保护区植物beta多样性及其影响因素 [J]. 生物多样性, 2022, 30(3): 21274. doi: 10.17520/biods.2021274.]
[24] ZHENG W C, ZHU A J, ZHANG F G, et al. Conservation priorities for plants in Jiulongshan National Nature Reserve [J]. J Zhejiang For Sci Technol, 2012, 32(6): 39–43. [郑伟成, 朱爱军, 张方纲, 等. 九龙山自然保护区国家重点保护野生植物优先保护序列研究 [J]. 浙江林业科技, 2012, 32(6): 39–43. doi: 10.3969/j.issn.1001-3776.2012.06.008.]
[25] PAN J G, WEI Z. Studies on Natural Resources of Jiulongshan Nature Reserve in Zhejiang [M]. Beijing: Forestry Press of China, 1995: 1–13, 139–157. [潘金贵, 韦直. 浙江省九龙山自然保护区自然资源研究 [M]. 北京: 中国林业出版社, 1995: 1–13,139–157.]
[26] SONG Y C. Vegetation Ecology [M]. 2nd ed. Beijing: High Education Press, 2017: 35–61,585–591. [宋永昌. 植被生态学 [M]. 第2版. 北京: 高等教育出版社, 2017: 35–61,585–591.]
[27] JOST L. Partitioning diversity into independent alpha and beta compo- nents [J]. Ecology, 2007, 88(10): 2427–2439. doi: 10.1890/06-1736.1.
[28] LENNON J J, KOLEFF P, GREENWOOD J J D, et al. The geogra- phical structure of British bird distributions: Diversity, spatial turnover and scale [J]. J Anim Ecol, 2001, 70(6): 966–979. doi: 10.1046/j.0021- 8790.2001.00563.x.
[29] SI X F, ZHAO Y H, CHEN C W, et al. Beta-diversity partitioning: Methods, applications and perspectives [J]. Biodiv Sci 2017, 25(5): 464–480. [斯幸峰, 赵郁豪, 陈传武, 等. Beta多样性分解: 方法、应用与展望 [J]. 生物多样性, 2017, 25(5): 464–480. doi: 10.17520/ biods.2017024.]
[30] DONG J Y, SUN X, ZHAN Q P, et al. Patterns and drivers of beta diversity of subtidal macrobenthos community on the eastern coast of Laizhou Bay [J]. Biodiv Sci, 2022, 30(3): 21388. [董建宇, 孙昕, 詹启鹏, 等. 莱州湾东岸潮下带大型底栖动物群落beta多样性格局及其驱动因素 [J]. 生物多样性, 2022, 30(3): 21388. doi: 10.17520/ biods.2021388.]
[31] YAO Z L, WEN H D, DENG Y, et al. Driving forces underlying the beta diversity of tree species in subtropical mid-mountain moist evergreen broad-leaved forests in Ailao Mountains [J]. Biodiv Sci, 2020, 28(4): 445–454. [姚志良, 温韩东, 邓云, 等. 哀牢山亚热带中山湿性常绿阔叶林树种beta多样性格局形成的驱动力 [J]. 生物多样性, 2020, 28(4): 445–454. doi: 10.17520/biods.2019356.]
[32] OKSANEN J, SIMPSON G L, BLANCHET F G, et al. Vegan: Community ecology package [EB/OL]. [2022-07-05]. https://CRAN.R- project.org/package=vegan.
[33] BATES D, MÄCHLER M, BOLKER B, et al. Fitting linear mixed- effects models using lme4 [J]. J Stat Softw, 2015, 67(1): 1–48. doi:10.18637/jss.v067.i01.
[34] DRAY S, BAUMAN D, BLANCHET G, et al. Adespatial: Multivariate multiscale spatial analysis [EB/OL]. [2022-07-05]. https://CRAN.R- project.org/package=adespatial.
[35] WENG C L, ZHANG T T, WU D H, et al. Drivers and patterns of- and-diversity in ten main forest community types in Gutianshan, eastern China [J]. Biodiv Sci, 2019, 27(1): 33–41. [翁昌露, 张田田, 巫东豪, 等. 古田山10种主要森林群落类型的和多样性格局及影响因素 [J]. 生物多样性, 2019, 27(1): 33–41. doi: 10.17520/biods. 2018171.]
[36] SOININEN J, HEINO J, WANG J J. A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosys- tems [J]. Glob Ecol Biogeogr, 2018, 27(1): 96–109. doi:10.1111/geb. 12660.
Species Diversity Pattern of Arbor Layer Species inCommunities in Jiulongshan, Zhejiang Province
ZHENG Zihong1, GUO Xiaohua1, BAO Xiaomei2, TANG Zhansheng1, ZHOU Shiwang1, LUO Zhengrong3*
(1. Administration Center of Jiulongshan National Nature Reserve, Suichang 323300, Zhejiang, China; 2. Ecological Forestry Development Center of Suichang County,Suichang 323300, Zhejiang, China; 3. College of Ecology, Lishui University,Lishui 323000, Zhejiang, China)
To understand the changes in species diversity ofcommunity and its driving factors, 35 sample circles with a 15 m radius centered on antree were set up in the Jiulongshan National Nature Reserve, Zhejiang Province. The species and diameter at breast height (DBH) of arbor layer trees (i.e. woody plants with a DBH more than 2.5 cm) within the sample circles were investigated. The relationship betweendiversity of arbor layer trees and altitude, and slope aspect, and DBH ofineach sample circle were analyzed by linear mixed model, and the influences of spatial structure, altitude, and slope aspect, and DBH ofondiversity were studied using the mantel test and variation partition analyses. The results showed that the number of species in arbor layer ofcommunities was rich, with 145 woody species belonging to 98 genera and 50 families, and dominated by deciduous or semi-evergreen species. The Shannon-Wiener index ranged from 2.37 to 3.40 and the Simpson index from 0.86 to 0.94. Thediversity indices increased at first then decreased with elevation, and neither related to the slope aspect nor DBH of. The Sorenson index of community pairs ranged from 0.15 to 0.95 and the contribution of turnover components to the totaldiversity of arbor layer was 74.88%. The geographical distance between sample circles was significantly positively correlated withdiversity and its components of arbor layer trees; while the altitude difference between sample circles was significantly positively correlated withdiversity and species-richness difference of arbor layer trees. The DBH difference ofbetween sample circles was only positively correlated with species turnover of arbor layer trees. The geographical distance of the sample circles is the likeliest explanation fordiversity and its two components (23%–29%) of arbor layer trees. The altitude difference is also a probable explanation fordiversity of arbor layer trees (16%). Therefore,diversity of arbor layer trees in thecommunity in Jiulongshan was mainly affected by altitude, whilediversity and its two components of arbor layer trees were affected by dispersal limitation, habitat filtering, and interspecific interaction betweenand associated species.
-diversity;diversity; Rare species; Altitude; Spatial distance
10.11926/jtsb.4707
2022-07-26
2022-10-11
浙江省自然科学基金项目(LY20C030002);遂昌县科技局县校合作项目(2020-hz27)资助
This work was supported by Project for Natural Science in Zhejiang (Grant No. LY20C030002), and the Project for County-School Cooperation in Science and Technology Bureau of Suichang (Grant No. 2020-hz27).
郑子洪(1968年生),男,本科,主要从事植物资源保护与开发利用研究。E-mail: zzh19680803@163.com
通讯作者 Corresponding author.E-mail: zrluo@126.com
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
中华戏曲(2020年1期)2020-12-04 04:12:11
中华戏曲(2020年1期)2020-02-12 02:28:26
测绘学报(2019年11期)2019-11-20 01:31:42
甘肃农业(2017年12期)2017-07-25 09:18:40
土壤与作物(2015年3期)2015-12-08 00:46:55
河北工业科技(2015年4期)2015-02-27 13:15:35
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
植物营养与肥料学报(2012年3期)2012-10-26 02:54:52
