
大气CO2浓度升高和氮添加对镉污染下两种树木叶片元素含量及回收率的影响
2023-08-05 03:51:44肖美娟尧波张桂华温达志
热带亚热带植物学报 2023年4期
肖美娟, 尧波, 张桂华, 温达志*
大气CO2浓度升高和氮添加对镉污染下两种树木叶片元素含量及回收率的影响
肖美娟1,2,3, 尧波4, 张桂华1,2, 温达志1,2*
(1. 中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广州 510650;2. 华南国家植物园,广州 510650;3. 中国科学院大学,北京 100049;4. 江西师范大学地理与环境学院,南昌 330022)
选择造林树种大叶相思()和樟树()为对象,采用开顶生长箱(open-top chamber, OTC)构建实验林,探讨镉(Cd)胁迫及其与大气CO2浓度升高、氮(N)添加的复合作用对2种树木叶片N、P养分利用策略和Cd积累特征的影响。试验设5个处理:对照(CK)、加Cd [10 kg Cd/(hm2·a)] (Cd)、加Cd与加CO2(700mol/mol) (CdC)、加Cd与加N [100 kg N/(hm2·a)] (CdN)、加Cd加CO2加N (CdCN)。处理约2.5 a后,测定两树种的成熟叶和衰老叶中N、磷(P)和Cd含量。结果表明,不同处理对两树种叶片P含量及P回收率均无显著影响,但显著影响叶片N含量及N回收率;CdN处理下两树种叶片N含量升高;Cd和CdN处理下樟树叶N回收率显著降低。Cd、CdC、CdN和CdCN处理下两树种叶片Cd积累浓度及樟树成熟叶中的Cd/N和Cd/P升高。大叶相思叶片N、P回收率显著高于樟树,Cd积累浓度显著低于樟树。速生豆科固氮树种大叶相思比普通非固氮树种樟树具有更好的N、P养分利用策略和抵御叶片积累Cd的能力,表明大叶相思可以作为Cd污染林地土壤生态修复的适宜树种。
镉污染;氮添加;CO2浓度升高;养分回收;化学计量比
随着工业发展和城市扩张的加快,我国土壤重金属污染日益突出。据调查,我国总的土壤污染超标率达16.1%,其中镉(Cd)的超标率高达7%,污染面积最为广泛[1]。同时,由于人类活动的影响如化石燃料的大量燃烧和土地利用方式的剧烈改变,大气CO2浓度和氮(N)沉降量持续上升。大气CO2浓度已由18世纪中叶的280mol/mol增加到当前的410mol/mol, 到21世纪末预计将超过700mol/mol[2]。据估计,1860年全球N沉降量为15 Tg N,1995年达到了156 Tg N,预计2050年将达到270 Tg N[3]。事实上,在我国快速城市化和工业化地区,重金属污染和高的大气CO2浓度、N沉降并存, 这些因子的存在势必对植物的生长产生重要影响。
大气CO2浓度升高和N添加对植物生长具有“施肥效应”,同时可以增强植物积累重金属Cd和耐受Cd胁迫的能力[4]。大气CO2浓度升高通过提高光合作用底物浓度刺激植物生长,增加地下碳分配,使根际沉积物如根系分泌物和有机酸等增加, 降低土壤pH,使土壤中的Cd活性增强,促进Cd从土壤向植物迁移[5];同时,大气CO2浓度升高可以增强植物抵抗非生物胁迫、调节养分或通过增加根系分泌物络合有毒离子的能力[6]。据报道,氮肥能促进植物生长,并产生各种解毒蛋白质,增强植物对Cd的耐受性[7],另一方面,氮肥使土壤酸化,土壤pH降低,增强Cd活性,促进植物对Cd的吸收[8]。这些研究表明单独的CO2浓度升高、N添加在促进植物生物量增加的同时也促进了对Cd的提取,但目前很少有研究报道CO2浓度升高和N添加复合作用对植物Cd吸收的影响。
“养分回收”是指养分从植物衰老组织转移到其他生长组织,实现养分重复利用的过程[9]。植物在叶片凋落之前将叶片中的养分转移到其他组织中,避免了大量的营养元素随叶片凋落而损失,因此养分回收是植物保存养分的重要机制之一,可以减少植物对土壤养分供应的依赖,增强植物对贫瘠生境的适应能力[10]。N和磷(P)是植物生长所需的大量元素,通常被认为是生态系统生产力的限制元素[11]。根据生态化学计量理论,Han等[12]提出“相对回收假说”,该假说认为木本植物在养分充足的土壤环境中,养分回收率保持平衡(即N回收率等于P回收率),但如果受到N (或P)的限制,则应从衰老叶片中吸收更多的N (或P)。以往的研究表明大气CO2浓度升高往往使叶片C/N和C/P升高,造成N和P的相对缺乏[13],因此植物通过提高N和P的回收率来满足生长需求。长期的氮沉降导致土壤氮素的增加比其他元素的增加更多,造成养分失衡,植物叶片的N/P上升,使叶片N回收率下降,并导致植物生长受到P限制[14]。但是关于在Cd胁迫下,植物养分回收是如何响应CO2浓度升高、N添加及其复合作用的研究仍鲜见报道。
植物修复技术可以减少Cd污染地区的水土流失及Cd迁移,进而改善生态环境,对于矿区废弃地的生态恢复具有现实意义,这一技术的要点在于筛选能够耐受高浓度Cd胁迫并能正常生长发育的树种。同时,具有高效的养分利用策略的植物更容易定植在养分贫瘠的重金属污染地区。了解Cd胁迫下,不同植物叶片的Cd耐受性和养分回收特征对全球变化的响应,可以为Cd污染地区的植物修复治理筛选合适的造林树种。大叶相思()为豆科速生常绿乔木,耐旱耐贫瘠, 固氮能力强,是我国南方地区造林绿化、水土保持和作薪炭材的主要树种之一。樟树()为樟科常绿阔叶乔木,树冠发达,落叶丰富、且地下部分有强大的根系,是水土保育的良好树种。因此,本研究基于开顶生长箱(open-top chamber, OTC),探讨在Cd胁迫下,大气CO2浓度升高、N添加及其复合作用对大叶相思和樟树N、P养分吸收及Cd积累特征的影响,拟解决的科学问题是:(1)不同树种叶片N、P养分和重金属Cd浓度以及回收率在不同处理间是否存在显著差异?(2)不同树种的叶片N、P养分和重金属Cd浓度以及回收率与土壤、叶片的营养状态有何关联?这些问题的探讨有助于理解在全球变化背景下,生长在Cd污染环境中的植物养分利用策略以及对Cd的耐受机制,有利于筛选用于Cd污染场地造林的优良树种。
1 材料和方法
1.1 研究区概况
研究地位于广州市中国科学院华南植物园科研区(23°10′46.4″ N,113°21′9.1″ E),该区域海拔18 m,属于典型的南亚热带季风气候,年平均气温约21.5 ℃,年降雨量为1 600~1 900 mm,旱季和雨季分明,其中4月—9月为降雨季,降雨量约占全年的80%[15]。
1.2 样地设置
研究采用开顶生长箱(OTC)的近自然法。OTC由底座、箱体及充气系统组成。底座为下沉式不锈钢体(2.5 m×2.5 m×0.6 m)。底座上方为圆柱形箱体,直径3.0 m,箱体由8根不锈钢支柱固定, 铁丝网逐级缠绕加固,之后围上透光塑料膜。充气系统包括PVC管、气瓶、鼓风机及CO2流量计。PVC管(内径为7.5 cm)围绕箱体一周,朝中央一侧每隔1 cm开1个0.1 cm的小孔。气瓶中的CO2在鼓风机作用下通过PVC管上均匀分布的小孔扩散到OTC内, CO2浓度通过流量计调节。OTC内的土壤(0~10、10~30和30~60 cm)采自受干扰少的罗浮山自然保护区, 并与自然林土壤层一致。经检测,生长箱内的光强度是箱外开放空间的97%,未检测到光谱变化,开顶保持空气流动,OTC内气温略高于周边但温差不显著。
本研究设置了对照(CK:不加Cd、N,自然环境下的CO2浓度)、Cd [仅加10 kg Cd/(hm2·a)]、CdC (加Cd与CO2浓度升高)、CdN (加Cd和N)和CdCN (加Cd、N与CO2浓度升高)共5个处理,每处理3个重复,共15个OTC。每天8:00—17:00 (雨天除外)按700mol/mol的CO2浓度进行充气,由于气流、天气状况的影响,OTC内CO2浓度约为(700± 20)mol/mol。据报道广州市大气N沉降量为39~ 49 kg N/(hm2·a)[16],同时为模拟矿山周边的Cd污染状况, 将N和Cd的添加量均设定为100 kg N/(hm2·a)和10 kg Cd/(hm2·a)。分别将配置好的CdCl2和NH4NO3溶液,每月喷施到土壤表面,将等量的水喷施到对照组。2016年9月选择1 a生大叶相思和樟树树苗各3株,等比例和等距离种植于OTC内,待树苗在OTC中生长稳定后,于2017年4月开始进行实验处理。
1.3 样品的采集与测定
于2019年9月同时采集大叶相思和樟树的成熟叶和衰老叶。成熟叶选取树冠东南西北4个方向的枝条上由基部向顶端的第5~6片、完全展开的健康叶片。衰老叶为未脱落叶,但已经在功能上分离的叶片,其特征为颜色变红或黄,轻摇植株就脱落或用手轻触可脱落。每个OTC中每株大叶相思和樟树采集成熟叶、衰老叶20~30片,叶片带回实验室后,放置于60 ℃烘箱中烘干至恒重, 使用球磨仪将样品粉碎至粉末状,用于测定叶片Cd、N和P元素含量。Cd含量测定参考GB 5009.268—2016《食品安全国家标准食品中多元素的测定》, N含量测定采用靛酚蓝分光光度法,P含量测定采用钼锑抗分光光度法。
于2019年10月采集不同处理下的表层(0~10 cm)土壤,测定其化学性质(表1)。土壤pH测定采用电位法,土壤总氮测定采用靛酚蓝分光光度法,土壤总磷测定采用钼锑抗比色法,土壤铵态氮测定采用氯化钾浸提-靛酚蓝比色法,土壤硝态氮测定采用酚二磺酸比色法,土壤有效磷测定采用盐酸-氯化铵浸提-钼锑抗比色法。土壤全镉测定参照HJ 832—2017《土壤和沉积物金属元素总量的消解》,土壤有效镉测定参照HJ 804—2016《土壤8种有效态元素的测定二乙烯三胺五乙酸浸提-电感耦合等离子体发射光谱法》。
1.4 数据分析
成熟叶和衰老叶的Cd、N、P化学计量特征采用质量含量比表示。用回收率(resorption efficiency, RE)和回收度(resorption proficiency, RP)来量化元素回收,前者是指叶片在衰老过程中回收元素含量与成熟叶中对应元素含量的比值,后者则直接用衰老叶中的元素含量表示,衰老叶中元素含量越低表示其回收度越高[17–18]。叶片元素回收率(resorption efficiency, RE)基于质量含量计算[17],RE(1–N/N×MCLF)100%,其中,N和N分别为衰老叶和成熟叶中的元素含量(mg/g),MLCF为质量损失修正系数,阔叶树种值为0.78,用CdRE、NRE和PRE分别表示叶片Cd、N和P的回收率,CdRP、NRP和PRP分别表示叶片Cd、N和P的回收度。
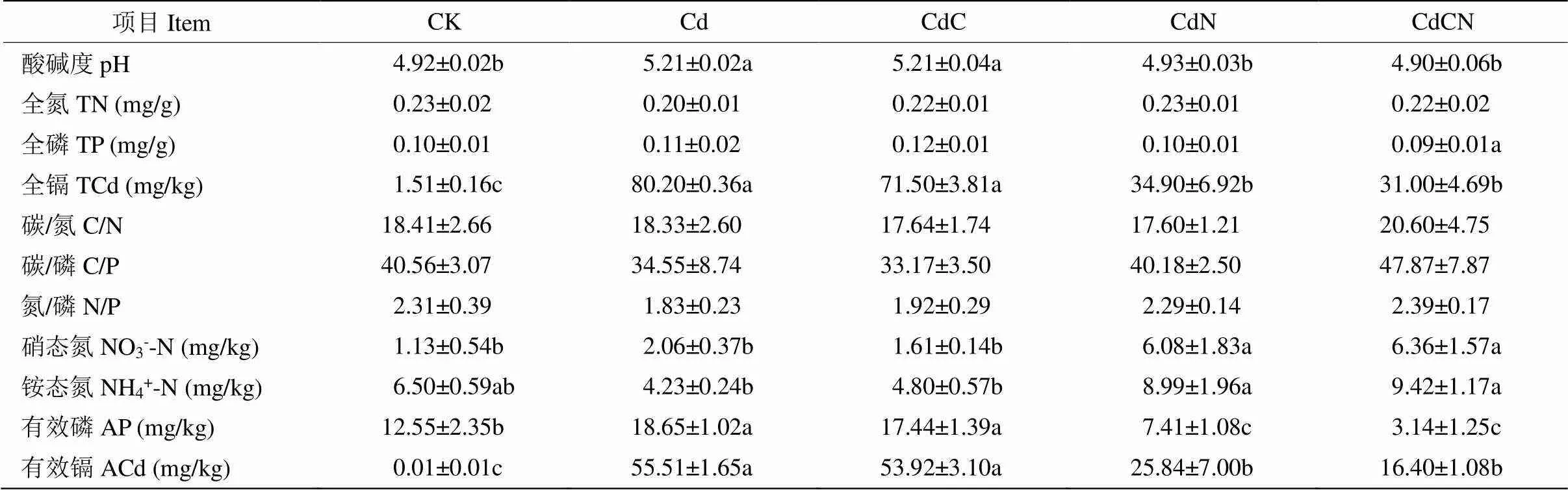
表1 0~10 cm土壤化学性质
3; CK: 对照; Cd: +Cd; CdC: +Cd+CO2; CdN: +Cd+N; CdCN: +Cd+CO2+N。同行数据后不同字母表示差异显著(0.05)(Duncan’s检验)。下同
3. CK: Control; Cd: +Cd; CdC: +Cd+CO2; CdN: +Cd+N; CdCN: +Cd+CO2+N. Data followed different letters at the same line indicate significant difference at 0.05 level by Duncan’s test. The same below
采用双因素方差分析(Two-Way ANOVA)比较不同处理、树种及其交互作用对成熟和衰老叶Cd、N、P含量及化学计量比、元素回收的影响;采用单因素方差分析(One-Way ANOVA)和Duncan检验对不同处理下的植物成熟叶和衰老叶Cd、N、P含量及化学计量比、元素回收率的差异和显著性进行比较。采用Pearson相关分析法,对叶片Cd、N、P含量及回收率与叶片、土壤元素含量、生态化学计量比进行相关分析。数据统计分析在Excel 2012、SPSS 22.0和R 3.5.1软件中进行。
2 结果和分析
2.1 对成熟叶片N、P和Cd含量及其化学计量比的影响
从表2可见,成熟叶Cd、N含量及化学计量比值(Cd/N、Cd/P)受处理的影响显著,且因不同树种而异;处理与树种的交互作用仅对成熟叶Cd含量影响显著(0.05)。比较而言,CdN处理下的两树种成熟叶N含量均最高,同时大叶相思成熟叶N含量显著高于樟树(图1);成熟叶P含量在不同处理和不同树种之间的差异均不显著。与对照相比, 加Cd处理(Cd、CdC、CdN和CdCN)均促进Cd在两树种尤其樟树叶片中的积累,樟树成熟叶Cd含量比对照增加了11.1~13.1倍(0.05);加Cd处理对大叶相思成熟叶Cd/N、Cd/P和N/P均无显著影响,但显著提高了樟树成熟叶Cd/N (对照的952~1 567倍)和Cd/P (对照的934~1 451倍),打破了樟树成熟叶的元素平衡,加剧了Cd毒害风险。
2.2 对衰老叶元素含量及回收的影响
从表2可见,处理对叶片NRP、CdRP (即衰老叶N和Cd含量)的影响显著,树种对叶片NRP、PRP、CdRP (即衰老叶N、P、Cd含量)以及NRE、PRE的影响显著,处理与树种的交互作用仅对NRP (即衰老叶N含量)的影响显著(0.05)。两树种衰老叶N和Cd含量在处理间均存在显著差异: Cd污染下加N处理(CdN和CdCN)的大叶相思衰老叶N含量显著高于不加N处理(Cd和CdC),CdN处理的樟树衰老叶N含量显著高于对照和CdC处理;CdN处理的大叶相思衰老叶Cd含量显著高于对照,加Cd处理(Cd、CdC、CdN和CdCN)的樟树衰老叶Cd含量均显著高于对照(0.05, 图2)。与对照相比,加Cd处理(Cd、CdC、CdN和CdCN)的两树种衰老叶P含量总体呈增加趋势(Cd处理的大叶相思除外), 但不同处理间的差异不显著。
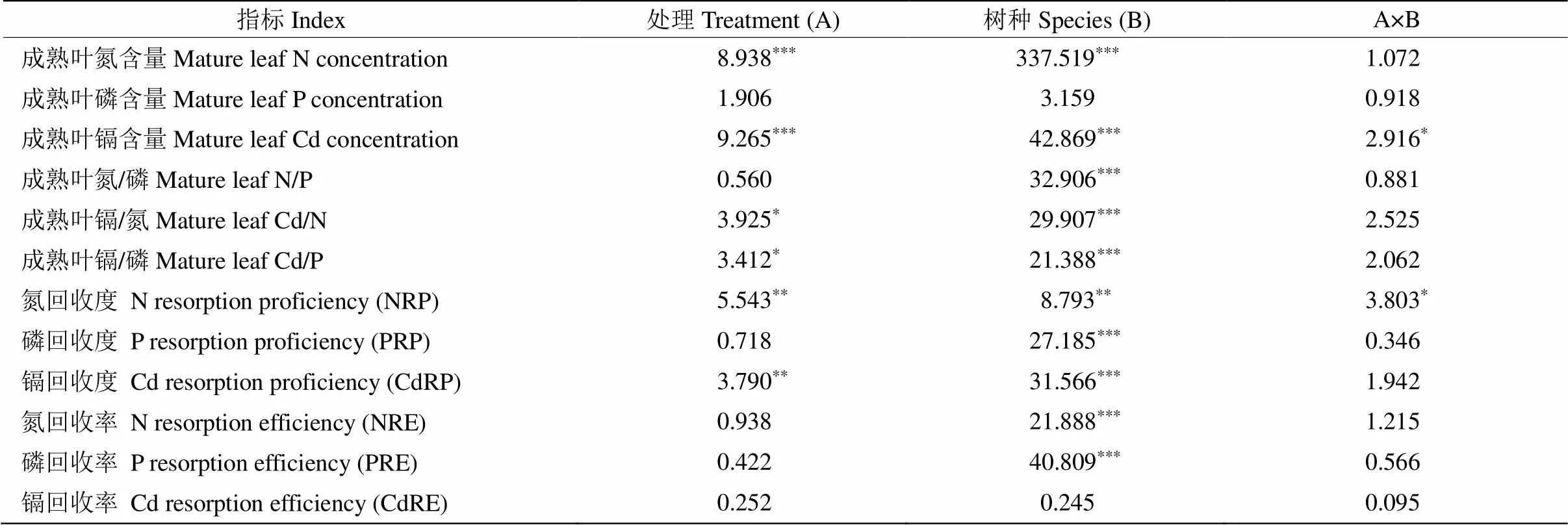
表2 处理和树种对叶片元素含量、化学计量比及元素回收影响的F值(双因素方差分析)
*:<0.05; **:<0.01; ***:<0.001。下同
*:<0.05; **:<0.01; ***:<0.001. The same below
大叶相思叶片N、P和Cd的回收率(NRE、PRE和CdRE)在处理间差异均不显著(图2)。樟树叶片CdRE和PRE在处理间不存在显著差异,但NRE在处理间差异显著,与对照相比,Cd和CdN处理的樟树叶片NRE显著下降(-44%、-40%) (0.05)。大叶相思和樟树衰老叶的P、Cd含量以及叶片NRE、PRE存在显著差异:樟树衰老叶P和Cd含量显著高于大叶相思,NRE和PRE显著低于大叶相思。

图2 不同处理下叶片元素回收特征
2.3 叶片N、P和Cd含量及其回收率的影响因子
利用Pearson相关分析方法分析大叶相思和樟树叶片Cd、N、P含量及回收率与叶片、土壤元素含量及化学计量比的相关性。从表3可见,大叶相思成熟叶N含量与成熟叶P含量、衰老叶N和P含量以及土壤全氮(TN)、TN/全磷(TP)、土壤有效氮(AN)呈显著正相关,与NRE、土壤全镉(TCd)、TCd/TN、TCd/TP、有效镉(ACd)/AN呈显著负相关; 成熟叶P含量与成熟叶N含量、衰老叶N和P含量以及土壤TP、AN呈显著正相关,与PRE呈显著负相关;成熟叶Cd含量与成熟叶Cd/N、Cd/P呈显著正相关;衰老叶N含量与成熟叶N和P含量、衰老叶P含量以及土壤TN/TP、AN呈显著正相关,与NRE以及土壤TCd/TN、TCd/TP、ACd/AN呈显著负相关; 衰老叶P含量与成熟叶N和P含量、衰老叶N含量以及土壤TP、AN呈显著正相关,与PRE显著负相关;衰老叶Cd含量与成熟叶Cd/N、Cd/P呈显著正相关,与CdRE呈显著负相关;NRE与成熟叶N含量、衰老叶N含量、CdRE呈显著负相关,与土壤ACd、ACd/AN呈显著正相关;PRE与成熟和衰老叶P含量以及土壤TP呈显著负相关,与土壤TN/TP呈显著正相关;CdRE与衰老叶Cd含量、NRE以及土壤ACd、ACd/AN呈显著负相关(0.05)。
从表4可见,樟树成熟叶N含量与成熟叶P含量、衰老叶N和P含量、衰老叶N/P以及土壤TN/TP、AN呈显著正相关,与NRE、土壤ACd/AN呈显著负相关;成熟叶P含量与成熟叶N含量、衰老叶N和P含量、PRE以及土壤TP、TN/TP、AN呈显著正相关;成熟叶Cd含量与衰老叶Cd含量、成熟叶Cd/N和Cd/P以及土壤TCd、ACd呈显著正相关; 衰老叶N含量与成熟叶N和P含量、衰老叶P和Cd含量、土壤AN呈显著正相关,与NRE、CdRE呈显著负相关; 衰老叶P含量与成熟叶N和P含量、衰老叶N含量、成熟叶Cd/N和Cd/P、土壤TP呈显著正相关,与成熟叶N/P、NRE、PRE呈显著负相关; 衰老叶Cd含量与成熟叶Cd含量、Cd/N和Cd/P以及衰老叶N含量呈显著正相关,与NRE、CdRE呈显著负相关。NRE、PRE和CdRE三者间呈显著正相关;此外, NRE与成熟叶N含量、衰老叶N、P、Cd以及土壤TCd、TCd/TN、TCd/TP、ACd、ACd/AN呈显著负相关; PRE与成熟叶P含量、N/P以及土壤TN/TP呈显著正相关,其与衰老叶P含量、成熟叶Cd/P、土壤TCd、TCd/TN、TCd/TP呈显著负相关;CdRE与衰老叶N和Cd含量呈显著负相关,与成熟叶Cd/N和Cd/P呈显著正相关(<0.05)。
3 结论和讨论
3.1 不同处理对叶片N、P含量及回收特征的影响
在本研究中, Cd处理的大叶相思和樟树成熟叶片N、P含量均低于对照,与谭长强等[19]的研究结果相似,这是因为Cd的积累会影响根系发育进而抑制养分吸收[20]。土壤重金属Cd污染后,樟树成熟叶N含量在大气CO2浓度升高下降低,这是因为N被植物根系吸收后主要通过蒸腾作用进入植物叶片,而CO2浓度升高使植物气孔导度下降,蒸腾作用减弱,从而降低了植物叶片的N含量[21];相反,CO2浓度升高下的大叶相思成熟叶N含量并没有降低,这可能是因为大叶相思属于豆科植物,可以通过结瘤固氮[22],增加叶片氮含量。在Cd污染土壤中,加N处理(CdN和CdCN)增加了土壤AN含量,两树种成熟叶中的N、P含量均升高,表明植物生长对N需求增加的同时P需求也增加,以维持N和P稳态平衡,与Crowley等[23]报道叶片P含量随着N添加而升高的结果一致。此外,加N处理(CdN和CdCN)的土壤有效磷(AP)含量低于不加N处理(Cd和CdC),这可能是因为N添加促进了植物生长对土壤P吸收增加,导致土壤AP含量降低,若不补给适量P素,植物生长不断消耗土壤AP而受P限制,Li等[24]也认为随着N沉降量的增加,P对生物量产量的限制将进一步加剧。CdCN处理下的两树种叶片N含量高于CdC处理但低于CdN处理,这可能是因为CO2浓度升高和N添加促进植物生长,使成熟叶N含量被稀释,但土壤AN的增加使叶片N含量升高,抵消了一部分生长稀释的作用。比较而言,CdCN处理对大叶相思成熟叶P吸收的促进作用大于CdC和CdN处理;对樟树成熟叶P吸收的促进作用则小于CdN处理、大于CdC处理,表明CO2浓度升高和N添加对大叶相思叶片P吸收表现为加和作用,对樟树叶片P吸收则可能存在交互作用。Huang等[25]的研究结果表明CO2浓度升高和N添加的复合作用有利于豆科植物与非豆科植物竞争P,本研究结果与之类似,在Cd污染土壤中, CO2浓度升高和N添加复合处理下的大叶相思(豆科植物)成熟叶P含量高于樟树(非豆科植物)成熟叶。

表3 大叶相思叶片N、P、Cd含量及回收率与叶片、土壤元素含量及化学计量比的Pearson相关系数

表4 樟树叶片N、P、Cd含量及回收率与叶片、土壤元素含量及化学计量比的Pearson相关系数
植物的生长不仅依赖于土壤中可利用的营养物质,还取决于各种营养物质之间的平衡,因此植物养分回收能力不仅与土壤养分的绝对含量有关, 还与养分的相对含量有关[26]。N/P比值可以作为反映植物生长养分限制的指标,N/P<14、N/P>16、16N/P14分别表示N限制、P限制、N和P共同限制[27]。对照和Cd、CdC、CdN处理下的大叶相思成熟叶N/P均大于16,而CdCN处理下的大叶相思成熟叶N/P为14~16,表明在Cd、CdC、CdN处理下,大叶相思主要受到P限制,而在CdCN处理下,大叶相思受N和P共同限制。对照和不同处理下的樟树成熟叶N/P均低于14,表明樟树主要受到N限制。Killingbeck[18]认为当植物衰老叶中的N含量低于7 mg/g,P含量低于0.5 mg/g时,N、P养分的回收是完全的;而当N含量大于10 mg/g、P含量大于0.8 mg/g,则N、P养分的回收是不完全的。本研究结果表明,各处理的大叶相思衰老叶N含量均大于10 mg/g,P含量均低于0.5 mg/g,表明固氮树种大叶相思衰老叶中的N养分回收不完全, 而P养分回收较完全。Cd、CdN和CdCN处理的樟树衰老叶N含量均大于10 mg/g, N养分回收均不完全, 对照和CdC处理的樟树衰老叶N含量为7~10 mg/g, N养分回收介于完全回收和不完全回收之间。仅对照的樟树衰老叶P含量低于0.5 mg/g,表现为P养分的完全回收,而不同处理的樟树衰老叶P含量为0.5~0.8 mg/g,P养分回收介于完全回收和不完全回收之间。这表明大叶相思对衰老叶P的回收是完全的,而且其PRE高于NRE,基本符合“养分回收假说”,表明大叶相思可以根据自身养分需求调节N和P养分的回收过程,对Cd污染环境的适应能力较强。樟树在受到N限制的情况下,没有对衰老叶N进行完全回收,而且樟树叶片CdRE与NRE显著正相关,之前的研究也证明大多数元素的回收模式具有很强的共变性和耦合性[28],表明樟树为了逃避Cd毒害,将更多的Cd积累在衰老叶中的同时减少了对衰老叶N的回收,导致其养分回收能力下降, 需要消耗能量从土壤中吸收N以满足生长需求,不利于生物量的积累。
有研究表明叶片养分回收率与土壤养分供应、叶片养分状况的关系并不总是一致,养分回收率可能与绿叶养分状况无关、负相关或者正相关,也可能与土壤养分供应无关、负相关或者正相关[26]。另一些研究者则认为衰老叶养分变化是控制养分回收能力的主导因子[18]。本研究结果表明大叶相思和樟树的N、P回收度和回收率与土壤养分供应和叶片养分状况均存在相关关系。通常认为豆科植物的NRE比非豆科植物低,PRE比非豆科植物高[29], 本研究结果表明,大叶相思(豆科植物)的NRE和PRE均高于樟树(非豆科植物),与之前的结果有所不同。前人[30]研究表明豆科植物的NRE比非豆科植物低的原因是豆科植物除了能利用土壤中的N素外还可以通过形成根瘤固定大气中的N2,在N素吸收方面优于非豆科植物。本研究结果表明大叶相思的成熟叶N含量高于樟树,表明大叶相思对N素的需求高于樟树,同时有研究表明重金属Cd抑制豆科植物的结瘤作用[31],因此大叶相思通过提高NRE和NRE以满足其快速生长。综上所述,叶片的N、P养分回收能力因树种和生存环境而异。
3.2 不同处理对叶片Cd含量及积累特征的影响
本研究结果表明,加Cd处理(Cd、CdC、CdN和CdCN)的大叶相思和樟树叶片Cd含量均显著高于对照,这是因为施加Cd使土壤Cd含量大幅提高,同时Cd在土壤-植物系统中流动性强,具有很高的植物积累指数[32]。CO2浓度升高有利于植物生长, 同时会增加根系H+或有机酸的释放,降低土壤pH,而土壤pH越低,Cd的生物有效性越高,植物对Cd的吸收越多[33]。Wang等[34]研究表明大气CO2浓度升高增加了杨树和柳树的生物量和Cd吸收总量,但叶片Cd含量并没有改变,表明植物叶片的Cd含量由于植物生物量的增加而被稀释。本研究结果与之类似,但原因不同,我们之前的研究结果表明在Cd污染土壤中,CO2浓度升高对大叶相思和樟树生长的促进作用不显著,土壤pH值在CdC与Cd处理间也没有显著差异,因此在Cd污染土壤中, CO2浓度升高处理下,大叶相思和樟树成熟叶Cd含量没有显著变化[35]。之前的研究表明土壤N含量的增加促进了植物对Cd的吸收和积累[36],本研究表明在Cd污染土壤中,加N处理(CdN和CdCN)使大叶相思叶片Cd含量呈现增加的趋势,研究结果类似。其原因可能包括以下两个方面:首先,N添加导致土壤酸化(pH值下降),增加了Cd的生物有效性;其次,植物没有专门吸收Cd2+的途径,Cd2+往往通过Ca2+通道或与其他二价离子(Fe2+、Mn2+和Zn2+)共用的转运蛋白进入植物根系[37],而施N引起土壤酸化导致金属阳离子(如K+、Ca2+、Mg2+等)流失加剧[38],大大提高了Cd进入植物根系的机会,进而促进了植物积累Cd。与此相反,N添加使樟树成熟叶Cd含量略呈现下降趋势(–1.2%),我们之前的研究发现N添加显著促进樟树生长[35],因此生长带来的稀释作用是N添加导致樟树叶片Cd含量下降的主要原因,Lin等[39]的研究也表明水稻Cd含量由于稀释作用而下降。在Cd污染土壤中, CO2浓度升高和N添加对大叶相思和樟树成熟叶Cd含量影响的交互作用不显著,这可能是因为单独的CO2浓度升高对大叶相思和樟树成熟叶Cd含量没有显著影响,单独N添加同时促进了植物生长和Cd吸收,而生长带来的稀释作用导致叶片Cd含量变化不大,最终表现为CO2浓度升高和N添加对成熟叶Cd浓度影响的交互作用不显著。
据报道,当成熟叶片中Cd含量超过3 mg/kg时, Cd对植物具有毒害作用[40]。本研究中,加Cd处理(Cd、CdC、CdN和CdCN)的大叶相思成熟叶Cd含量均低于3 mg/kg,而樟树成熟叶Cd含量均高于3 mg/kg,表明上述加Cd处理对大叶相思成熟叶的毒害作用小,但对樟树成熟叶的毒害作用增加。植物吸收并积累Cd会诱发元素计量比的失衡、代谢紊乱,抑制植物生长[41]。N和P养分对于植物的生长和解毒尤为重要,N可以促进叶绿素、解毒蛋白质的合成来缓解Cd的毒害作用[42],P可以促进DNA、RNA及核糖体RNA (rRNA)等合成,帮助细胞分裂,加速根尖、叶片等组织分生,促进植物生长[43]。因此,降低Cd/N和Cd/P可以促进植物快速生长、减少Cd的毒害作用。本研究表明大叶相思成熟叶的Cd/N、Cd/P在对照与加Cd处理(Cd、CdC、CdN和CdCN)之间均没有显著差异,反映出其元素计量化学的内稳态特征,这可能是通过较高的NRE、PRE来实现。相反,樟树成熟叶在Cd胁迫下Cd/N、Cd/P显著高于对照,这可能是成熟叶中较高的Cd积累和较低的N、P回收所致, 打破了营养元素与有毒元素之间的稳态平衡,大大增加Cd毒害的风险。我们前期研究表明,大叶相思的生物量是樟树的10倍[35],进一步证明大叶相思能够通过快速生长、较高的养分(N、P)回收和维持Cd、N、P化学计量稳态平衡等策略来抵御Cd的毒害。
在Cd污染土壤中,CO2浓度升高、N添加及其复合作用对两树种衰老叶中的Cd含量(即Cd回收度)以及CdRE的影响均不显著,并且两树种的CdRE均为负值,表明Cd在衰老叶中富集,与万雪琴等[44]对杨树()的结果类似。这是因为Cd不同于N、P等营养元素,并不是植物生长发育的必需元素,同时Cd的回收积累会加剧对植物的毒害作用,因此植物进行元素迁移回收时, 并不会主动回收Cd,而是将Cd富集在衰老叶中,通过叶片凋落的形式将Cd排出植物体,进而避免Cd对植物成熟叶中叶绿素合成和光合作用的负面影响,降低Cd对植物的毒害作用。这种解毒作用在樟树中尤其明显,可能因为樟树成熟叶中较高的Cd含量所致。值得注意的是,叶片衰老过程中富集Cd是植物解毒的一个重要策略,但通过凋落叶又重新归还到土壤,这部分Cd的形态、稳定以及是否引起次生污染需进一步研究。
综上,在Cd污染土壤中,大气CO2浓度升高、N添加及其复合作用对叶片N、P、Cd含量及其回收率产生不同程度的影响。N添加显著提高了大叶相思和樟树成熟叶N含量以及前者衰老叶N含量,N添加与CO2浓度升高的复合作用显著提高大叶相思衰老叶N含量。N、P养分回收和Cd积累的种间差异显著,大叶相思叶片Cd积累浓度显著低于樟树,NRE和PRE则显著高于樟树,表明大叶相思具有更强的抵御Cd吸收的能力和更高的N、P利用效率。本研究还表明,樟树与大叶相思在Cd污染胁迫下具有不同的适应策略,樟树通过将吸收的Cd转移到衰老叶,随凋落叶排出体外;而大叶相思则通过维持养分元素与Cd元素之间的化学计量比(Cd/N和Cd/P)内稳态调节机制,即从土壤中吸收Cd同步从衰老叶中回收更多的N和P,以抵消Cd带来的不利影响。由于樟树凋落叶Cd重新归还到土壤, 并且其生物量远低于大叶相思,因此,比较而言,速生豆科树种大叶相思是Cd污染林地生态修复早期的优选树种。
[1] Ministry of Environmental Protection and Ministry of Land and Resources of the People’s Republic of China. Report on the national general survey of soil contamination [EB/OL]. (2014-04-17) [2021-12- 10]. [环境保护部, 国土资源部. 全国土壤污染状况调查公报 [EB/OL]. (2014-04-17) [2021-12-10]. http://www.gov.cn/foot/2014-04/ 17/content_2661768.htm.]
[2] IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovern- mental Panel on Climate Change [M]. Cambridge: Cambridge University Press, 2013.
[3] GALLOWAY J N, TOWNSEND A R, ERISMAN J W, et al. Transfor- mation of the nitrogen cycle: Recent trends, questions, and potential solutions [J]. Science, 2008, 320(5878): 889–892. doi: 10.1126/science. 1136674.
[4] LUO Y Q, HUI D F, ZHANG D Q. Elevated CO2stimulates net accumulations of carbon and nitrogen in land ecosystems: A meta- analysis [J]. Ecology, 2006, 87(1): 53–63. doi: 10.1890/04-1724.
[5] NIE M, LU M, BELL J, et al. Altered root traits due to elevated CO2: A meta-analysis [J]. Glob Ecol Biogeogr, 2013, 22(10): 1095–1105. doi: 10.1111/geb.12062.
[6] JIA X, LIU T, ZHAO Y H, et al. Elevated atmospheric CO2affected photosynthetic products in wheat seedlings and biological activity in rhizosphere soil under cadmium stress [J]. Environ Sci Pollut Res, 2016, 23(1): 514–526. doi: 10.1007/s11356-015-5288-7.
[7] ZHANG S M, YANG C, CHEN M M, et al. Influence of nitrogen availability on Cd accumulation and acclimation strategy ofleaves under Cd exposure [J]. Ecotoxicol Environ Saf, 2019, 180: 439– 448. doi: 10.1016/j.ecoenv.2019.05.031.
[8] BAI Z Q, ZHU L, CHANG H X, et al. Enhancement of cadmium accumulation in sweet sorghum as affected by nitrate [J]. Plant Biol, 2021, 23(1): 66–73. doi: 10.1111/plb.13186.
[9] KILLINGBECK K T. The terminological jungle revisited: Making a case for use of the term resorption [J]. Oikos, 1986, 46(2): 263–264. doi: 10.2307/3565477.
[10] FRESCHET G T, CORNELISSEN J H C, VAN LOGTESTIJN R S P, et al. Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: What is the link with other resource economics traits? [J]. New Phytol, 2010, 186(4): 879–889. doi: 10.1111/j.1469-8137. 2010 03228.x.
[11] XU M P, ZHU Y F, ZHANG S H, et al. Global scaling the leaf nitrogen and phosphorus resorption of woody species: Revisiting some commonly held views [J]. Sci Total Environ, 2021, 788: 147807. doi: 10.1016/j.scitotenv.2021.147807.
[12] HAN W X, TANG L Y, CHEN Y H, et al. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants [J]. PLoS One, 2013, 8(12): e83366. doi: 10.1371/ journal.pone.0083366.
[13] HESSEN D O, ÅGREN G I, ANDERSON T R, et al. Carbon seque- stration in ecosystems: The role of stoichiometry [J]. Ecology, 2004, 85(5): 1179–1192. doi: 10.1890/02-0251.
[14] ZHENG J, SHE W W, ZHANG Y Q, et al. Nitrogen enrichment alters nutrient resorption and exacerbates phosphorus limitation in the desert shrub[J] Ecol Evol, 2018, 8(20): 9998–10007. doi: 10.1002/ece3.4407.
[15] HUANG W J, ZHOU G Y, LIU J X, et al. Mineral elements of subtropical tree seedlings in response to elevated carbon dioxide and nitrogen addition [J]. PLoS One, 2015, 10(3): e0120190. doi: 10.1371/ journal.pone.0120190.
[16] LI J, FANG Y T, YOH M, et al. Organic nitrogen deposition in preci- pitation in metropolitan Guangzhou City of southern China [J]. Atmos Res, 2012, 113: 57–67. doi: 10.1016/j.atmosres.2012.04.019.
[17] VERGUTZ L, MANZONI S, PORPORATO A, et al. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants [J]. Ecol Monogr, 2012, 82(2): 205–220. doi: 10.1890/ 11-0416.1.
[18] KILLINGBECK K T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency [J]. Ecology, 1996, 77(6): 1716–1727. doi: 10.2307/2265777.
[19] TAN C Q, HUANG Z L, PENG Y H, et al. Influences of cadmium and nitrogen on the accumulation and distribution of dry matter and N, P, K, Cd in[J]. Chin J Ecol, 2017, 36(7): 1847–1854. [谭长强, 黄志玲, 彭玉华, 等. Cd胁迫下施N对台湾桤木()干物质及N、P、K、Cd积累与分配的影响 [J]. 生态学杂志, 2017, 36(7): 1847–1854. doi: 10.13292/j.1000-4890.201707.016.]
[20] THAN T A, POPOVA L P. Functions and toxicity of cadmium in plants: Recent advances and future prospects [J]. Turk J Bot, 2013, 37(1): 1–13. doi: 10.3906/bot-1112-16.
[21] MCDONALD E P, ERICKSON J E, KRUGER E L. Research note: Can decreased transpiration limit plant nitrogen acquisition in elevated CO2? [J]. Funct Plant Biol, 2002, 29(9): 1115–1120. doi: 10.1071/ fp02007.
[22] LONG S R. Rhizobium-legume nodulation: Life together in the under- ground [J]. Cell, 1989, 56(2): 203–214. doi: 10.1016/0092-8674(89) 90893-3.
[23] CROWLEY K F, MCNEIL B E, LOVETT G M, et al. Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern united states? [J]. Ecosystems, 2012, 15(6): 940–957. doi: 10.1007/s10021-012-9550-2.
[24] LI Y, NIU S L, YU G R. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis [J]. Glob Change Biol, 2016, 22(2): 934–943. doi: 10.1111/gcb.13125.
[25] HUANG W J, ZHOU G Y, LIU J X, et al. Effects of elevated carbon dioxide and nitrogen addition on foliar stoichiometry of nitrogen and phosphorus of five tree species in subtropical model forest ecosystems [J]. Environ Pollut, 2012, 168: 113–120. doi: 10.1016/j.envpol.2012. 04.027.
[26] ZHOU L L, ADDO-DANSO S D, WU P F, et al. Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry ofwith stand development in southern China [J]. J Soils Sediments, 2016, 16(5): 1448–1459. doi: 10.1007/s11368-016-1352-2.
[27] GÜSEWELL S. N:P ratios in terrestrial plants: Variation and functional significance [J]. New Phytol, 2004, 164(2): 243–266. doi: 10.1111/j. 1469-8137.2004.01192.x.
[28] PRIETO I, QUEREJETA J I. Simulated climate change decreases nutrient resorption from senescing leaves [J]. Glob Change Biol, 2020, 26(3): 1795–1807. doi: 10.1111/gcb.14914.
[29] JIANG D L, GENG Q H, LI Q, et al. Nitrogen and phosphorus resorption in planted forests worldwide [J]. Forests, 2019, 10(3): 201. doi: 10.3390/f10030201.
[30] MOKHELE B, ZHAN X J, YANG G Z, et al. Review: Nitrogen assimilation in crop plants and its affecting factors [J]. Can J Plant Sci, 2012, 92(3): 399–405. doi: 10.4141/cjps2011-135.
[31] CHEN Y X, HE Y F, YANG Y, et al. Effect of cadmium on nodulation and N2-fixation of soybean in contaminated soils [J]. Chemosphere, 2003, 50(6): 781–787. doi: 10.1016/S0045-6535(02)00219-9.
[32] YANG D P, GUO Z Q, GREEN I D, et al. Effect of cadmium accu- mulation on mineral nutrient levels in vegetable crops: Potential implications for human health [J]. Environ Sci Pollut Res, 2016, 23(19): 19744–19753. doi: 10.1007/s11356-016-7186-z.
[33] LI L Z, WU H F, VAN GESTEL C A M, et al. Soil acidification increases metal extractability and bioavailability in old orchard soils of Northeast Jiaodong Peninsula in China [J]. Environ Pollut, 2014, 188: 144–152. doi: 10.1016/j.envpol.2014.02.003.
[34] WANG R G, DAI S X, TANG S R, et al. Growth, gas exchange, root morphology and cadmium uptake responses of poplars and willows grown on cadmium-contaminated soil to elevated CO2[J]. Environ Earth Sci, 2012, 67(1): 1–13. doi: 10.1007/s12665-011-1475-0.
[35] YI Y F. Growth dynamics and nutrient utilization strategies of selected subtropical woody species under combined treatments of rising atmo- spheric CO2, elevated nitrogen and cadmium [D]. Beijing: University of Chinese Academy of Sciences, 2019: 1–49. [易亚凤. 大气CO2浓度升高、N添加与Cd胁迫复合影响下亚热带木本植物生长动态与养分利用策略[D]. 北京: 中国科学院大学, 2019: 1–49.]
[36] YANG Y J, XIONG J, TAO L X, et al. Regulatory mechanisms of nitrogen (N) on cadmium (Cd) uptake and accumulation in plants: A review [J]. Sci Total Environ, 2020, 708: 135186. doi: 10.1016/j.scito tenv.2019.135186.
[37] CHEN X H, OUYANG Y N, FAN Y C, et al. The pathway of trans- membrane cadmium influx via calcium-permeable channels and its spatial characteristics along rice root [J]. J Exp Bot, 2018, 69(21): 5279–5291. doi: 10.1093/jxb/ery293.
[38] STEVENS C J, DISE N B, GOWING D J. Regional trends in soil acidification and exchangeable metal concentrations in relation to acid deposition rates [J]. Environ Pollut, 2009, 157(1): 313–319. doi: 10. 1016/j.envpol.2008.06.033.
[39] LIN Y L, CHAO Y Y, HUANG W D, et al. Effect of nitrogen deficiency on antioxidant status and Cd toxicity in rice seedlings [J]. Plant Growth Regul, 2011, 64(3): 263–273. doi: 10.1007/s10725-011-9567-0.
[40] ISMAEL M A, ELYAMINE A M, MOUSSA M G, et al. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers [J]. Metallomics, 2019, 11(2): 255–277. doi: 10.1039/c8mt00247a.
[41] SARWAR N, SAIFULLAH, MALHI S S, et al. Role of mineral nutri- tion in minimizing cadmium accumulation by plants [J]. J Sci Food Agric, 2010, 90(6): 925–937. doi: 10.1002/jsfa.3916.
[42] ZHANG F, WAN X Q, ZHONG Y. Nitrogen as an important detoxi- fication factor to cadmium stress in poplar plants [J]. J Plant Interact, 2014, 9(1): 249–258. doi: 10.1080/17429145.2013.819944.
[43] YANG Y J, CHEN R J, FU G F, et al. Phosphate deprivation decreases cadmium (Cd) uptake but enhances sensitivity to Cd by increasing iron (Fe) uptake and inhibiting phytochelatins synthesis in rice () [J]. Acta Physiol Plant, 2016, 38(1): 28. doi: 10.1007/s11738- 015-2055-9.
[44] WAN X Q, ZHANG F, XIA X L, et al. Effects of cadmium stress on absorption and distribution of mineral nutrients in poplar plants [J]. Sci Silv Sin, 2009, 45(7): 45–51. [万雪琴, 张帆, 夏新莉, 等. 镉胁迫对杨树矿质营养吸收和分配的影响 [J]. 林业科学, 2009, 45(7): 45–51.]
Effects of Elevated Atmospheric CO2Concentration and Nitrogen Addition on Element Concentration and Resorption in Leaves of Two Tree Species Under Cadmium Pollution
XIAO Meijuan1,2,3, YAO Bo4, ZHANG Guihua1,2, WEN Dazhi1,2*
(1. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystem, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. South China National Botanical Garden, Guangzhou 510650, China; 3. University of Chinese Academy of Sciences,Beijing 100049, China; 4. School of Geography and Environment, Jiangxi Normal University,Nanchang 330022, China)
In order to investigate the effects of cadmium (Cd) pollution and its combinations with elevated atmospheric CO2concentration and nitrogen (N) addition on N and phosphorus (P) utilization strategies and Cd accumulation in leaves, two afforestation tree species (and) were selected, and the open-top chambers (OTCs) was used to construct the simulated experimental forests of the two tree species. The five treatments were control (CK), adding Cd [10 kg Cd/(hm2·a)] (Cd), adding Cd and CO2(700mol/mol) (CdC), adding Cd and N [100 kg N/(hm2·a)] (CdN), adding Cd, CO2and N (CdCN). After treatment for about 2.5 years, the mature and senescent leaves of two tree species were collected to measure the concentrations of N, P and Cd. The results showed that the foliar P concentration and P resorption efficiency had no significant changes under different treatments, but the N concentration and N resorption efficiency in leaves were significantly affected. The N concentration in leaves of two species increased under CdN treatment, and the N resorption efficiency inleaves decreased significantly under Cd and CdN treatments. Under the treatments of Cd, CdC, CdN and CdCN, the Cd accumulation in leaves of two species, as well as the Cd/N and Cd/P ratios in mature leaves ofincreased. The N and P resorption efficiencies were significantly higher, while the Cd accumulation was significantly lower inthan those in. It was demonstrated that the fast-growing leguminous N-fixing tree species () had better N and P utilization strategies and stronger resistance to Cd accumulation than the common non-N-fixing tree species (), suggesting thatcould be suitable for ecological restoration in Cd polluted forest sites.
Cadmium pollution; Nitrogen addition; Elevated CO2concentration; Nutrient resorption; Stoichiometric ratio
10.11926/jtsb.4598
2021-12-23
2022-02-27
国家自然科学基金项目(31570483); 广东省自然科学基金重点项目(2015A030311029)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31570483), and the Key Project for Natural Science in Guangdong (Grant No. 2015A030311029).
肖美娟(1995年生),女,硕土研究生,主要从事环境生态研究。E-mail: xiaomeijuan@scbg.ac.cn
通讯作者 Corresponding author.E-mail: dzwen@scib.ac.cn
猜你喜欢
新作文·小学低年级版(2022年4期)2022-08-30 07:23:00
北方音乐(2019年6期)2019-07-16 07:50:53
现代园艺(2018年2期)2018-03-15 08:00:35
现代园艺(2018年3期)2018-02-10 05:18:14
现代园艺(2017年23期)2018-01-18 06:57:54
现代园艺(2017年11期)2017-06-28 11:32:18
兽医导刊(2016年6期)2016-05-17 03:50:53
中国林业产业(2016年5期)2016-04-03 00:33:02
中国林业产业(2016年5期)2016-04-03 00:32:38
武夷学院学报(2014年5期)2014-07-19 10:08:27
