
连续炭基肥替代化肥对菜园土壤性质和细菌群落结构的影响
2023-08-05 03:51叶菁王义祥刘岑薇林怡黄家庆翁伯琦
热带亚热带植物学报 2023年4期
叶菁, 王义祥,2*, 刘岑薇, 林怡, 黄家庆, 翁伯琦
连续炭基肥替代化肥对菜园土壤性质和细菌群落结构的影响
叶菁1, 王义祥1,2*, 刘岑薇1, 林怡1, 黄家庆1, 翁伯琦1
(1. 福建省农业科学院农业生态研究所, 福建省红壤山地农业生态过程重点实验室,福州 350013;2. 福建省农业科学院土壤肥料研究所,福州 350013)
为了解生物炭基肥替代化肥减量施用的田间长期效应,利用定位试验研究连续5 a炭基肥替代化肥对蔬菜产量、土壤理化性质和细菌群落结构的影响。结果表明,连续5 a实施炭基有机肥替代化肥,土壤pH提高了0.13~0.25,土壤有机质、碱解氮和有效磷含量也分别提高了2.1%~62.2%、5.8%~86.0%和0.4%~103.1%,炭基肥替代化肥处理的荠菜()产量提高了4.0%~14.8%,但75%替代处理较50%替代处理有所降低。炭基肥替代化肥处理的土壤菌群Sobs、Shannon、Ace和Chao指数均高于单施化肥处理,且均以75%替代处理最高。炭基肥替代化肥显著降低了土壤中硝化菌属()、拟无枝酸菌属()、芽单胞菌属()等的丰度,增加了纤维素降解菌菌群(、)的丰度。芽单胞菌属、Ilumatobacteraceae、Methyloligellaceae等的丰度与土壤全氮、全磷、有机质间具有显著的相关性。可见,连续炭基肥替代化肥明显改善了菜地土壤的理化性质,从而影响了土壤细菌群落结构和多样性,适量替代可以提高蔬菜产量,但应注意长期连续施用对土壤pH、碳/氮的影响,避免过量施用抑制作物对养分的吸收利用。
菜园;生物炭基肥;化肥;土壤;细菌群落
我国是化肥生产和使用大国,2019年我国农用化肥施用折纯量为5.404×107t。近年来随着化肥减量增效政策的实施,我国化肥施用总量首次呈现略微降低的态势,但与发达国家相比仍存在较大的差距。尤其是对蔬菜作为高耗肥农作物,1997—2018年我国蔬菜平均化肥施用量为602.6 kg/hm2,是主粮作物的1.8倍[1]。在“双碳”目标下,为保障蔬菜生产,改善施肥种类以减少化肥使用和提高养分利用率是现代蔬菜产业发展亟需解决的重要问题。
生物炭基肥作为一种新型肥料,通过改善土壤理化性质、调节土壤微生物活性,有利于提高植物对养分的利用效率以及减少土壤中养分流失,近年来备受农业与环保领域的广泛关注和研究应用[2]。生物炭基肥的类型有炭基无机肥、炭基有机肥、炭基复混肥等,可用于大田作物水稻()、玉米()、小麦()以及设施蔬菜等,作物应用上总体表现为正调控效应,具有增产提质、节肥增效及固碳减排等作用[3],但目前多数生物炭基肥的研究中应用年限相对较短,其田间应用评价仍亟待加强[4]。生物炭基肥对作物生长、土壤改良效果与其种类、组成成分、土壤类型等因素有关,需进一步研究[5–6]。微生物在土壤养分循环、有机质分解以及土壤结构形成和稳定性方面发挥重要作用[7],其多样性指数的变化也是反映土壤健康状态的敏感指标[8]。有研究表明,生物炭施用提高了土壤pH和养分利用水平[9],生物炭的多孔性结构可为微生物提供适宜的生存环境[10],还可通过吸附养分或改良土壤性质间接影响微生物群落[11],但生物炭自身养分有限,不能满足作物生长的需要[12]。炭基有机肥可以发挥生物炭和有机肥的互补或协同作用,有利于提高肥效和弥补生物炭养分不足[13],因此,本文研究生物炭基肥替代化肥减量施用对菜园土壤性质和细菌群落结构的影响,旨为蔬菜生产中科学应用生物炭基肥提供参考。
1 材料和方法
1.1 材料
试验地位于福建省福清市渔溪镇(25°25′ N, 119°36′ E),属亚热带海洋气候,年均温18.8 ℃, 年均无霜期365 d,年降水量1 350~1 500 mm。土壤为黏质土,有机质含量1.37%,全氮、全磷和全钾含量分别为0.09%、0.06%和2.51%,碱解氮、有效磷和速效钾含量分别为42.0、35.2和193.04 mg/kg, pH值6.85。试验用生物炭由花生壳在500 ℃下裂解制备,其有机碳、全氮、全磷和全钾含量分别为566.1、10.2、2.5和4.7 g/kg,pH值9.31。炭基有机肥由花生壳生物炭、腐熟有机肥复配而成,其中生物炭、氮、P2O5和K2O的质量分数分别为9%、1.4%、3.4%和1.8%,pH 7.8。
1.2 试验设计
田间试验采用单因素随机裂区设计,共设4个处理,即炭基肥替代化肥氮量分别为0 (HF)、25% (B1)、50% (B2)和75% (B3),每处理3个小区重复,小区面积为8 m2。各处理等氮量设计,氮素施用量为180 kg/hm2。化肥分别为尿素、过磷酸钙和氯化钾。HF处理的化肥用量分别为尿素390 kg/hm2、过磷酸钙750 kg/hm2、氯化钾220 kg/hm2。B1、B2和B3处理分别利用相应量的炭基有机肥替代尿素氮,磷、钾不足部分用过磷酸钙和硫酸钾补足。试验开始于2017年3月下旬,每年种植蔬菜两茬, 第一茬(3月—7月)种植叶用甘薯(),第二茬(9月—次年1月)种植荠菜(), 每茬施肥1次。
1.3 蔬菜产量测定
于2021年1月,采收每个小区全部当季种植的荠菜,称取质量进行产量测定。
1.4 土壤采集和理化性质分析
连续5 a炭基肥替代化肥后,于2021年4月在各试验小区内以“S”型多点混合法采集0~20 cm土样,去除植物碎屑和碎石,混合样品后放于冷藏箱内带回实验室。
pH采用pH计以水土比2.5:1浸提测定,全氮含量采用凯氏定氮仪测定,有机质含量采用重铬酸钾和硫酸亚铁滴定法,碱解氮含量采用碱解扩散- H3BO3吸收滴定法;全磷和速效磷采用钼锑抗比色法,全钾采用火焰光度法测定[14–15]。
1.5 土壤菌群结构分析
采用土壤DNA提取试剂盒(Omega Bio-tek Inc, Doravilla, GA, USA)提取菜园土壤细菌的总DNA,用1%琼脂糖凝胶电泳检测DNA的质量,然后送至上海美吉生物医药科技有限公司,以Illumina MiSeq平台进行高通量测序和分析。
1.6 数据处理
数据分析基于上海美吉生物医药科技有限公司提供的云服务(https://www.i-sanger.com)进行。参照王义祥等[16]的方法,分析前以“按最小样本序列数”进行数据抽平处理,其中多样性指数(-diversity)利用mothur version v.1.30.1 (http://www.mothur.org/)计算;Qiime计算采用-多样性距离矩阵,以R语言作图;相关性Heatmap图的软件及算法采用R语言pheatmap package提供的方法。数据组间的比较采用单因素方差分析(One-Way ANOVA)和克氏秩和检验(Kruskal-Wallis H test)。菌群结构关联与模型预测采用Network共现性网络分析和单因素相关性网络分析进行分析。环境因子关联分析采用欧氏距离法。基于Spearman等级相关系数制作了丰度前50属与环境因子的相关性Heatmap图[17]。
2 结果和分析
2.1 荠菜产量的变化
HF、B1、B2和B3处理的荠菜产量分别为27.93、29.06、32.07和30.94kg/hm2。炭基有机肥替代处理较单施化肥处理(HF)均不同程度提高了荠菜产量, 为4.0%~14.8%,其中以B2处理的增幅最大, 且B2处理显著高于HF处理(<0.05),但HF、B1和B3处理间的差异均不显著(>0.05)。B2处理比B3处理高3.7%,但两者间的差异不显著(>0.05)。
2.2 土壤理化性质的变化
连续施用炭基肥5 a后,菜园土壤的pH值比单施化肥处理提高了0.13~0.25,其中B1和B2处理与HF处理间的差异达显著水平(<0.05)。从表1可见,随着炭基肥替代量的增加,土壤中全氮、有机质、全磷、全钾、碱解氮、有效磷、速效钾含量呈上升趋势, 其中土壤有机质含量增加了2.1%~62.2%,全氮增加了13.3%~48.9%,碱解氮含量提高了5.8%~86.0%, 有效磷含量提高了0.4%~103.1%,B2和B3处理与HF间的差异均达显著水平(<0.05),HF与B1处理间的差异不显著;但各处理间的全钾含量变化不大。

表1 土壤的理化性质
HF、B1、B2、B3分别表示炭基肥替代化肥氮量为0、25%、50%、75%;同列数据后不同小写字母表示差异显著(<0.05)。下同
HF, B1, B2 and B3 represent 0, 25%, 50% and 75% nitrogen replacement of biochar-based fertilizer, respectively. Data followed different letters within column indicate significant differences at 0.05 level. The same below
2.3 土壤菌群多样性的分析
多样性分析结果表明(表2),土壤菌群的Sobs、Shannon、Ace和Chao指数均以B3处理最高,且各指数均为B3>B2>B1>HF。Simpson指数以B1处理最高,且B1与B2、B3处理间的差异均达显著水平(<0.05)。覆盖度以HF处理最高,且与B3处理间的差异达显著水平(<0.05)。
对于-多样性,基于Bray-Curtis距离的PCoA分析表明,施肥处理显著影响了土壤菌群的分布(PERMANOVA,<0.05)。施肥处理能够解释菌群组成总体差异的29.1% (图1: A)。根据炭基肥中生物炭的用量对样本进行分组,炭基肥中生物炭的用量对解释菌群组成总体差异的34.0% (图1: B)。
2.4 土壤菌群的物种组成分析
在门水平上(图2: A),各处理土壤中主要的优势菌有放线菌门(Actinobacteriota)、变形杆菌门(Proteo- bacteria)、厚壁菌门(Chloroflexi)、酸杆菌门(Acido- bacteria)、厚壁菌门(Firmicutes)、拟杆菌门(Bacte- roidota)、芽单胞菌门(Gemmatimonadota)、Myxoco- ccota和Patescibacteria,其中放线菌门、变形杆菌门、厚壁菌门、酸杆菌门的丰度占80.7%以上。B2与B3处理的放线菌门相对丰度较HF处理分别降低了17.4%和15.3%,而酸杆菌门、厚壁菌门和Myxoco- ccota的相对丰度分别提高了61.5%和38.4%、40.4%和48.4%、20.0%和34.9%。
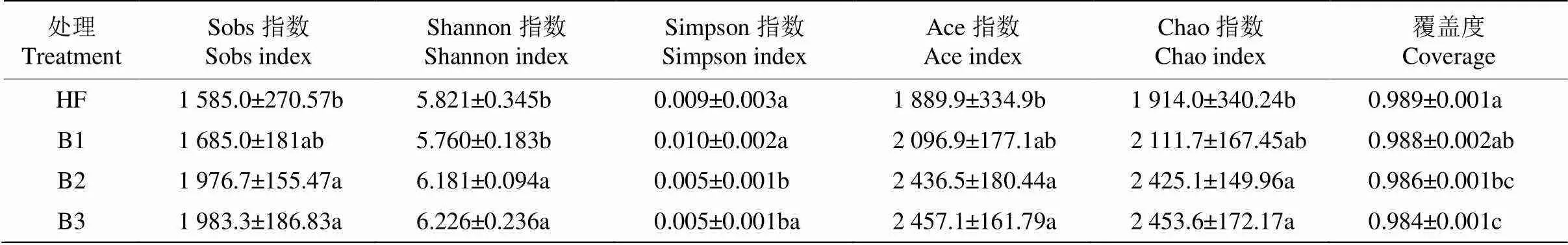
表2 土壤细菌的物种丰富度和多样性指数的变化
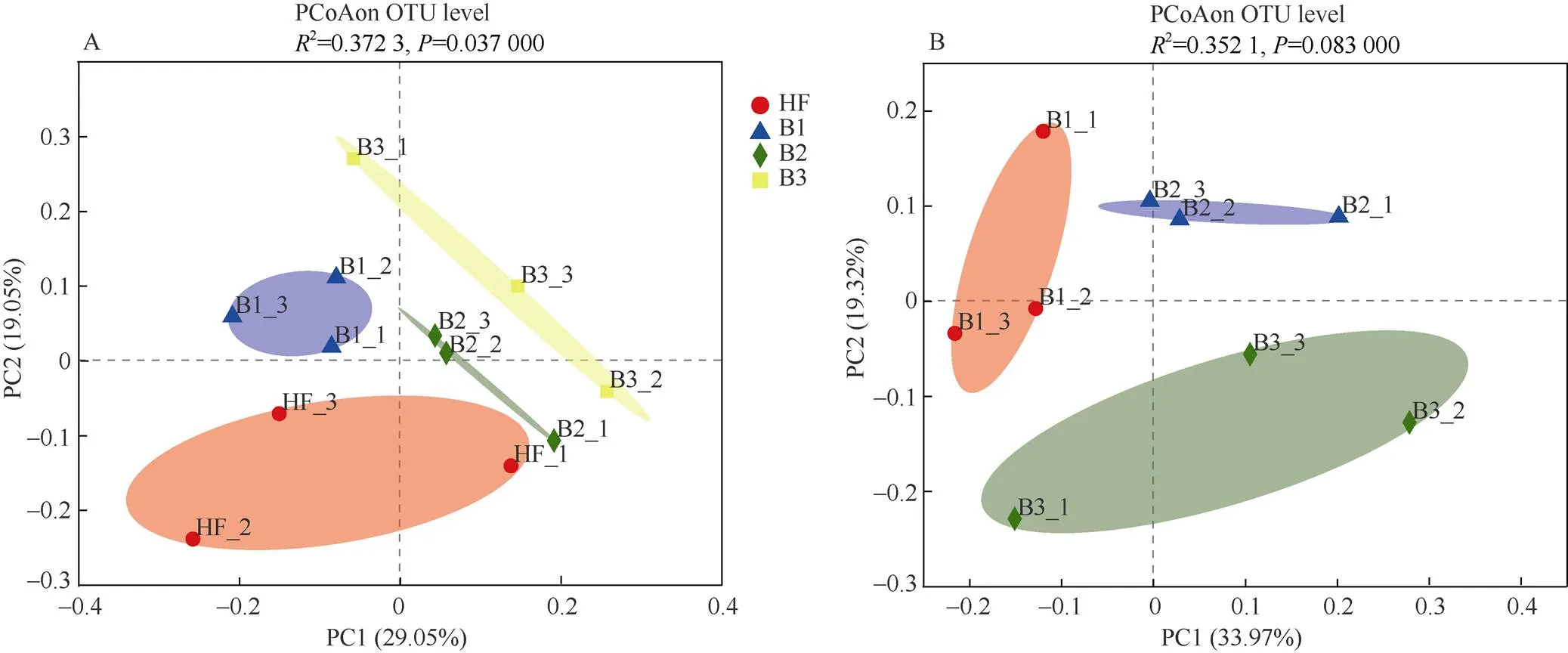
图1 土壤菌群的PCoA+Adonis分析。A: 施肥处理; B: 生物炭用量。
在属水平上,所有土壤样品中共注释到790属(图3: A), 各处理包含菌属的排序为B3>B2>B1>HF。其中4个处理的共有菌有517属,HF处理独有的20属,B1为16属,B2为18属,B3为31属。诺卡氏菌属(, 5.33%~8.66%)、鞘氨醇单胞菌属(, 4.04%~7.42%)、JG30-KF-CM45 (3.35%~6.00%)均是主要的优势菌属。除此之外, HF处理的优势菌属还有(3.35%)、大理石雕菌属(, 2.86%)和链霉菌属(, 2.77%)等(图3: B); B1处理的还有节杆菌属(, 3.24%)、Roseiflexaceae(3.13%)和KD4-96 (2.96%)。B2和B3处理的还有盖勒氏菌属(Gaiella, 2.94%~2.98%) (图2: B)。HF和B1处理丰度最高的菌属为诺卡氏菌属、鞘氨醇单胞菌属和JG30-KF-CM45;B2处理丰度最高的为诺卡氏菌属、JG30-KF-CM45B3和KD4-96;B3处理为诺卡氏菌属、和鞘氨醇单胞菌属。在属水平上的单因素方差分析(One-Way ANOVA)和Tukey- Kramer检验表明,4个处理间的硝化菌属()、薄壁芽孢杆菌属()、拟无枝酸菌属()、、、、JG30-KF-CM45和的丰度存在显著差异(<0.05),而芽单胞菌属()和的丰度存在极显著差异(<0.01)。
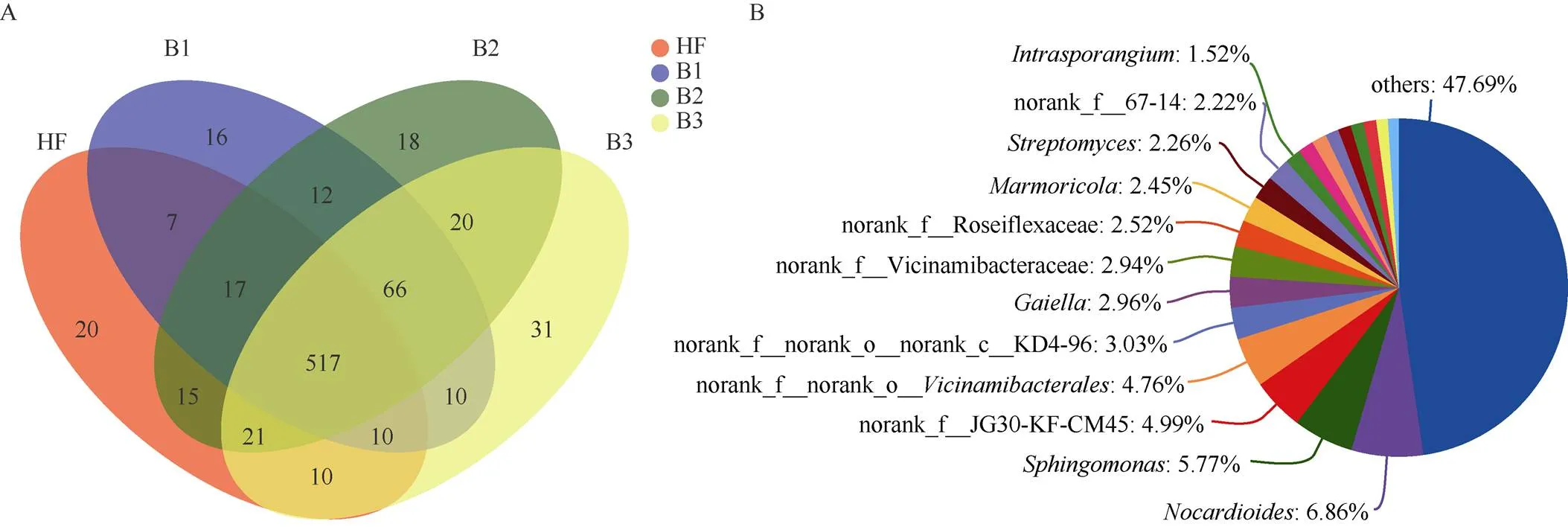
图3 基于属水平的菌群的venn图(A)和共有物种组成饼图(B)
2.5 土壤菌群结构关联与模型预测分析
共现性网络分析表明,属水平上所有物种共有2 643个有效节点(图4: A),HF、B1、B2和B3处理分别有617、655、686和685个节点。与4个处理均共线的属约有55.3%,至少有75.4%的属在2组以上的处理中共线。4个处理中有11属的共线率超过2%, 其中(6.93%)、(5.99%)、JG30-KF-CM45 (4.99%)、(4.62%)、(2.86%)、Vicinamibacteraceae (2.84%)、Rosei- flexaceae(2.55%)、(2.42%)和(2.23%);有9属共线率超过1%,其中(1.71%)、(1.58%)、Gemmati- monadaceae (1.46%)、(1.23%)和(1.01%)。以上20属确定为菜地土壤细菌群落的核心菌群。相关性网络分析表明(图4: B),网络中显示有19个节点和111条边,说明土壤菌群存在较高的连接性。Latescibacterota、Armatimonadota、Deinococcota、Acidobacteriota、Dadabacteria、NB1-j、Nitrospirota、Entotheonellaeota、RCP2-54、Myxoco- ccota等门的相关性较大,在整个菌群网络中起关键作用,其中Deinococcota与其他门呈负相关。
2.6 与环境因子的关联分析
环境因子对菌群结构影响的RDA分析表明, RDA1轴和RDA2轴对菌群结构差异的解释度分别为36.63%和16.69%。较高炭基肥替代量的B2和B3处理与HF和B1处理在RDA1轴方向上被分离开(图5: A)。Heatmap相关性分析表明(图5: B),的丰度与土壤pH、全氮、全磷、有机质、速效磷、速效钾、铵态氮间均具有显著的相关性(<0.05);、Ilumatobacteraceae、Methyloligellaceae的丰度与土壤全氮、有机质和全磷含量有显著相关性(<0.05);、、、、Xanthobacteraceae的丰度与土壤有机质含量具有显著的相关性(<0.05); Xanthobacter- aceae、、、、Roseiflexaceae、、Vicinamibacteraceae的丰度与土壤全磷含量有显著相关性(<0.05); 但大多数菌属与土壤全钾含量无显著相关性(>0.05)。
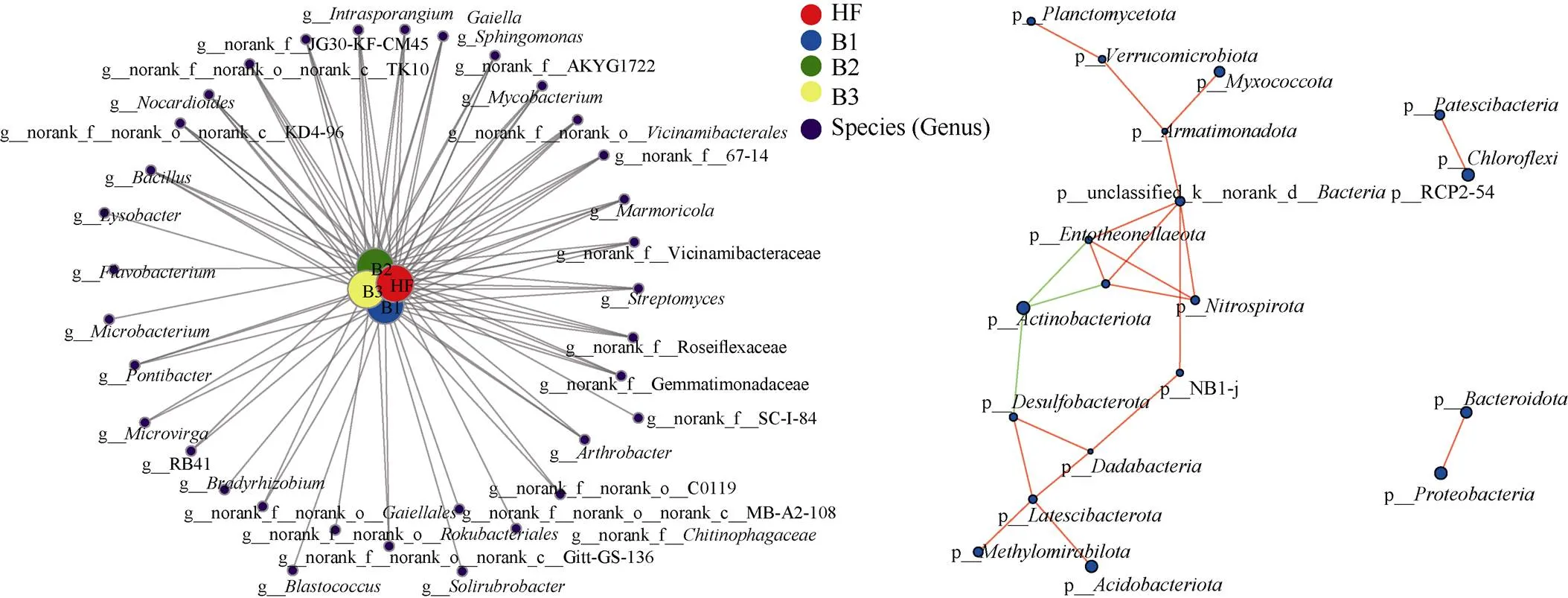
图4 共现性网络(A)和相关性网络图(B)
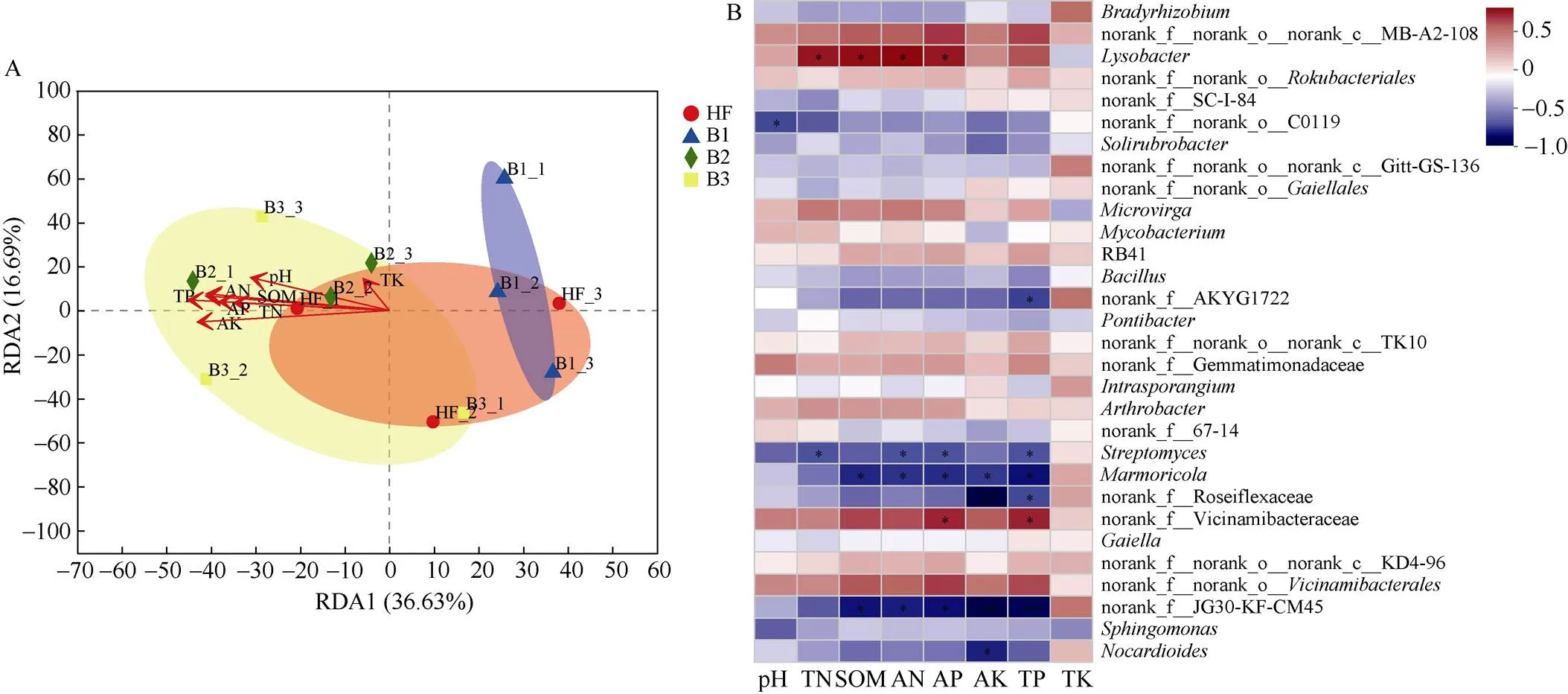
图5 土壤菌群结构与环境因子的冗余分析(A)和相关性分析(B)。TN: 全氮; SOM: 有机质; AN: 碱解氮; AP: 有效磷; AK: 速效钾; TP: 全磷; TK: 全钾。
3 结论和讨论
炭基肥替代化肥连续施用5 a后,土壤理化性质发生了较大变化, 菜地土壤pH提高了0.13~0.25,土壤有机质含量增加了2.1%~62.2%,且土壤中的碱解氮、有效磷和速效钾等速效养分含量的增幅高于全量养分含量的增幅,这可能是炭基肥有机肥中生物炭等具有大的表面积与良好的介孔结构,可吸附固定养分以降低土壤氮磷钾的流失[18]。生物炭基肥也能够抑制硝化细菌[19]、氨氧化变形杆菌[20]的活性,从而影响氮的硝化循环,降低氮素的损失。也有研究表明,生物炭基肥可提高土壤中磷溶解细菌、钾溶解细菌的活性,从而显著增加土壤中速效磷和速效钾的含量[2,21]。从对荠菜产量的影响来看,炭基肥替代化肥处理的荠菜产量均有不同程度的提高,但随着替代量的增加荠菜产量逐渐下降,这可能是因为荠菜适宜生长的土壤为微酸性或中性, 连续5 a的炭基肥施用导致土壤pH不断升高,而土壤过酸或过碱都不利于提高养分的生物有效性[22]。已有研究表明,土壤中C、N、P化学计量比决定着植物、微生物的养分可利用性,土壤C/N又是影响微生物活动导致土壤养分有效性变化的关键因素[23]。本试验初始时的土壤C/N相对较低,可供微生物利用的碳源较少,微生物活性降低,从而影响养分的有效性,进而不利于植物的生长发育[24],但随着炭基肥投入量的逐年累积,土壤C/N增大。当土壤C/N较高时,土壤微生物的繁殖要消耗一部分氮素,出现微生物与植物共同竞争土壤氮素,进而影响植物的生长[25]。因此,炭基肥连续施用应注意土壤类型以及对土壤pH、C/N的影响,避免过量施用抑制作物对养分的吸收利用。
土壤微生物群落与土壤理化性质关系密切,如土壤养分、pH、C/N等的改变都可能引起群落结构的变化。本研究结果表明,相对单施化肥处理,不同炭基肥处理的土壤菌群的Sobs、Shannon、Ace和Chao指数均有不同程度的提高;施肥处理显著影响了土壤菌群的分布,且能够单独解释菌群组成总体差异的29.1%,炭基肥中生物炭用量对解释菌群组成总体差异的34.0%。环境因子的关联分析表明,芽单胞菌属、Ilumatobacteraceae、Methyloligellaceae等与土壤全氮、全磷、有机质含量间具有显著的相关性,但与土壤全钾含量间无显著相关性。碳、氮和磷是微生物生长繁殖必需的营养物质,除了炭基肥中所含生物炭为微生物提供良好的栖息场所外,炭基肥是以生物炭与有机肥料配比制得,含有丰富的C、N、P等营养成分,能够直接为微生物生长提供所需的养分[26]。从土壤菌群结构来看,放线菌门、变形杆菌门、厚壁菌门、酸杆菌门、厚壁菌门、拟杆菌门、芽单胞菌门等是土壤中主要的优势菌门;诺卡氏菌属、鞘氨醇单胞菌属是主要的优势菌属。共现性网络分析表明,诺卡氏菌属、鞘氨醇单胞菌属、是本研究菜地土壤最为核心的菌群。已有研究表明,诺卡氏菌属、鞘氨醇单胞菌属中一些菌株对土壤除草剂具有生物降解作用[27–28];还可促进植物抵抗多种植物病原菌,如对引起黄萎病的真菌有拮抗作用[29]。从菌群的物种组成来看,相较于单施化肥处理,炭基肥处理显著降低了土壤中硝化菌属、拟无枝酸菌属、芽单胞菌属、卢德曼氏菌属、的丰度,增加了薄壁芽孢杆菌属、扁平丝菌苏黎世杆菌属、的丰度。硝化菌属是土壤氮素转化的功能菌群之一,主要是参与把亚硝酸盐氧化成硝酸盐的过程[30]。炭基肥的施用通过改变土壤性状和影响土壤中氮素转化,直接或间接地影响了硝化菌属的相对丰度[16]。本研究结果还表明,随着炭基肥替代量的增大,土壤中纤维素降解菌群(如、[31])的丰度呈升高的趋势,而嗜酸性菌群(如、)[32]的丰度则降低。
综上,炭基肥引发土壤性状改变(土壤pH、全氮、全磷、有机质含量提高),进而导致土壤菌群结构和多样性发生变化,改善了土壤微生物生态,本研究为炭基肥应用于菜园土壤培肥改良提供了重要的科学参考。
[1] Tao Y, Zhou Y X, Hu J L. Research on decomposition of driving factors of chemical fertilizer application intensity in China and control paths [J]. Acta Agric Zhejiang, 2021, 33(10): 1956–1970. [陶源, 周玉玺, 胡继连. 中国化肥施用强度的驱动因素分解与控制路径研究[J]. 浙江农业学报, 2021, 33(10): 1956–1970. doi: 10.3969/j.issn. 1004-1524.2021.10.20.]
[2] ZHAO Z Z, WANG X L, LI H B, et al. Slow-release property and soil remediation mechanism of biochar-based fertilizers [J]. J Plant Nutr Fert, 2021, 27(5): 886–897. [赵泽州, 王晓玲, 李鸿博, 等. 生物质炭基肥缓释性能及对土壤改良的研究进展[J]. 植物营养与肥料学报, 2021, 27(5): 886–897. doi: 10.11674/zwyf.20472.]
[3] LI Y M, ZHANG X C, LIAO S Q, et al. Research progress on synergy technologies of carbon-based fertilizer and its application [J]. Trans Chin Soc Agric Mach, 2017, 48(10): 1–14. [李艳梅, 张兴昌, 廖上强, 等. 生物炭基肥增效技术与制备工艺研究进展分析[J]. 农业机械学报, 2017, 48(10): 1–14. doi: 10.6041/j.issn.1000-1298.2017.10.001.]
[4] ZAMA E F, REID B J, ARP H P H, et al. Advances in research on the use of biochar in soil for remediation: A review [J]. J Soils Sedi, 2018, 18(7): 2433–2450. doi: 10.1007/s11368-018-2000-9.
[5] CHEW J, ZHU L L, NIELSEN S, et al. Biochar-based fertilizer: Super- charging root membrane potential and biomass yield of rice [J]. Sci Total Environ, 2020, 713: 136431. doi: 10.1016/j.scitotenv.2019.136431.
[6] LI Y L, CHENG J Z, LEE X Q, et al. Effects of biochar-based fertilizers on nutrient leaching in a tobacco-planting soil [J]. Acta Geochim, 2019, 38(1): 1–7. doi: 10.1007/s11631-018-0307-2.
[7] WANG S Q, LI T X, ZHENG Z C, et al. Soil aggregate-associated bacterial metabolic activity and community structure in different aged tea plantations [J]. Sci Total Environ, 2019, 654: 1023–1032. doi: 10. 1016/j.scitotenv.2018.11.032.
[8] SUN L, XUN W B, HUANG T, et al. Alteration of the soil bacterial community during parent material maturation driven by different fertilization treatments [J]. Soil Biol Biochem, 2016, 96: 207–215. doi: 10.1016/j.soilbio.2016.02.011.
[9] STEINBEISS S, GLEIXNER G, ANTONIETTI M. Effect of biochar amendment on soil carbon balance and soil microbial activity [J]. Soil Biol Biochem, 2009, 41(6): 1301–1310. doi: 10.1016/j.soilbio.2009.03. 016.
[10] LEHMANN J, RILLIG M C, THIES J, et al. Biochar effects on soil biota: A review [J]. Soil Biol Biochem, 2011, 43(9): 1812–1836. doi: 10.1016/j.soilbio.2011.04.022.
[11] CHEN K, XU X N, PENG J, et al. Effects of biochar and biochar-based fertilizer on soil microbial community structure [J]. Sci Agric Sin, 2018, 51(10): 1920–1930. [陈坤, 徐晓楠, 彭靖, 等. 生物炭及炭基肥对土壤微生物群落结构的影响[J]. 中国农业科学, 2018, 51(10): 1920– 1930. doi: 10.3864/j. issn.0578-1752.2018.10.011.]
[12] SOHI S P, KRULL E, LOPEZ-CAPEL E, et al. A review of biochar and its use and function in soil [J]. Adv Agron, 2010, 105: 47–82. doi: 10.1016/S0065-2113(10)05002-9.
[13] YANG J F, JIANG T, HAN X R, et al. Effects of continuous appli- cation of biochar-based fertilizer on soil characters and yield under peanuts continuous cropping [J]. Soils Fert Sci China, 2015(3): 68–73. [杨劲峰, 江彤, 韩晓日, 等. 连续施用炭基肥对花生土壤性质和产量的影响[J]. 中国土壤与肥料, 2015(3): 68–73. doi: 10.11838/sfsc. 20150312.]
[14] BAO S D. Soil Agrochemical Analysis [M]. Beijing: China Agriculture Press, 2007: 1–495.鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2007: 1–495.
[15] WANG H H, REN T B, YANG H J, et al. Research and application of biochar in soil CO2emission, fertility, and microorganisms: A sustainable solution to solve China’s agricultural straw burning problem [J]. Sustainability, 2020, 12(5): 1922.
[16] WANG Y X, HUANG J Q, YE J, et al. Effects of different amount of biochar application on soil property and bacterial community structure in acidified tea garden [J]. J Plant Nutr Fert, 2020, 26(11): 1967–1977. [王义祥, 黄家庆, 叶菁, 等. 生物炭对酸化茶园土壤性状和细菌群落结构的影响[J]. 植物营养与肥料学报, 2020, 26(11): 1967–1977. doi: 10.11674/zwyf.20237.]
[17] CAO P X, LIU Y X, XU G Q, et al. Bacterial diversity in the root system soil of[J]. Acta Ecol Sin, 2020, 40(14): 4954–4965. [曹鹏熙, 刘怡萱, 许国琪, 等. 冰川棘豆()根系土壤细菌多样性特征[J]. 生态学报, 2020, 40(14): 4954–4965. doi: 10.5846/stxb201907311624.]
[18] ZHANG X L, LIAO H, LI C W, et al. Effects of simultaneous application of biochar and chemical fertilizer on the vertical migration of nitrogen and phosphorus, lettuce yield and quality and soil microbial quantity under field conditions [J]. Acta Sci Circum, 2021, 41(1): 21– 28. [张雪莲, 廖洪, 李昌伟, 等. 田间条件下生物炭与化肥配施对土壤氮磷纵向迁移、结球生菜产量品质及土壤微生物数量的影响[J]. 环境科学学报, 2021, 41(1): 21–28. doi: 10.13671/j.hjkxxb.2020. 0564.]
[19] ZHANG Q Q, SONG Y F, WU Z, et al. Effects of six-year biochar amendment on soil aggregation, crop growth, and nitrogen and phosphorus use efficiencies in a rice-wheat rotation [J]. J Clean Prod, 2020, 242: 118435. doi: 10.1016/j.jclepro.2019.118435.
[20] BERGLUND L M, DELUCA T H, ZACKRISSON O. Activated carbon amendments to soil alters nitrification rates in Scots pine forests [J]. Soil Biol Biochem, 2004, 36(12): 2067–2073. doi: 10.1016/j. soilbio.2004.06.005.
[21] ZHENG H F, ZENG Y R, WANG C J, et al. Effects of biochar on phosphate solubilizing bacteria number and soil available P content in red soil tea plantation [J]. Chin Agric Sci Bull, 2018, 34(18): 114–118. [郑慧芬, 曾玉荣, 王成己, 等. 生物炭对红壤茶园溶磷细菌数量和土壤有效磷含量的影响[J]. 中国农学通报, 2018, 34(18): 114–118.]
[22] SPRINGOB G, KIRCHMANN H. Bulk soil C to N ratio as a simple measure of net N mineralization from stabilized soil organic matter in sandy arable soils [J]. Soil Biol Biochem, 2003, 35(4): 629–632. doi: 10.1016/S0038-0717(03)00052-X.
[23] SCHIPPER L A, SPARLING G P. Accumulation of soil organic C and change in C׃N ratio after establishment of pastures on reverted scrubland in New Zealand [J]. Biogeochemistry, 2011, 104(1/2/3): 49– 58. doi: 10.1007/s10533-009-9367-z.
[24] GE S F, XU H G, JI M M, et al. Effects of soil C:N on growth and distribution of nitrogen and carbon ofseedlings [J]. Chin J Plant Ecol, 2013, 37(10): 942–949. [葛顺峰, 许海港, 季萌萌, 等. 土壤碳氮比对平邑甜茶幼苗生长和碳氮分配的影响[J]. 植物生态学报, 2013, 37(10): 942–949. doi: 10.3724/SP.J.1258.2013.00097.]
[25] FARRELL M, KUHN T K, MACDONALD L M, et al. Microbial utilisation of biochar-derived carbon [J]. Sci Total Environ, 2013, 465: 288–297. doi: 10.1016/j.scitotenv.2013.03.090.
[26] DU H J, YU L Y, ZHANG Y Q. Recent advance on the genus: A review [J]. Acta Microbiol Sin, 2012, 52(6): 671–678. [杜慧竟, 余利岩, 张玉琴. 类诺卡氏属放线菌的研究进展[J]. 微生物学报, 2012, 52(6): 671–678. doi: 10.13343/j.cnki.wsxb.2012.06.010.]
[27] HU J, HE X H, LI D P, et al. Progress in research of[J]. Chin J Appl Environ Biol, 2007, 13(3): 431–437. [胡杰, 何晓红, 李大平, 等. 鞘氨醇单胞菌研究进展[J]. 应用与环境生物学报, 2007, 13(3): 431–437. doi: 10.3321/j.issn:1006-687X.2007.03.030.]
[28] BERG G, BALLIN G. Bacterial antagonists toKleb [J]. J Phytopathol, 1994, 141(1): 99–110. doi: 10.1111/j.1439-0434. 1994.tb01449.x.
[29] XU J Y, MAO Y P. From canonical nitrite oxidizing bacteria to complete ammonia oxidizer: Discovery and advances [J]. Microbiol China, 2019, 46(4): 879–890. [徐建宇, 毛艳萍. 从典型硝化细菌到全程氨氧化微生物: 发现及研究进展[J]. 微生物学通报, 2019, 46(4): 879–890. doi: 10.13344/j.microbiol.china.180194.]
[30] DING Y, HUANG Y, RUAN J S, et al. Selective isolation and diversity of acidophilic filamentous actinomycetes from acidic soils [J]. Acta Microbiol Sin, 2009, 49(6): 710–717. [丁芸, 黄英, 阮继生, 等. 嗜酸丝状放线菌的选择性分离与多样性 [J]. 微生物学报, 2009, 49(6): 710–717. doi: 10.3321/j.issn:0001-6209.2009.06.005.]
[31] ZHANG X M, XIAN H Q, LI Y H, et al. Functional detection of key microorganisms in tomato stalk composting by high-throughput mass spectrometer [J]. Shandong Agric Sci, 2020, 52(4): 73–78. [张小梅, 咸洪泉, 李雅华, 等. 高通量质谱检测番茄秸秆好氧堆肥中关键微生物的功能[J]. 山东农业科学, 2020, 52(4): 73–78. doi: 10.14083/ j.issn.1001-4942.2020.04.013.]
[32] WANG G Y, YANG Y, KONG Y L, et al. Key factors affecting seed germination in phytotoxicity tests during sheep manure composting with carbon additives [J]. J Hazard Mater, 2022, 421: 126809. doi: 10. 1016/j.jhazmat.2021.126809.
Effects of Continuous Biochar-based Fertilizer Replacement on Soil Properties and Bacterial Community Structure in Vegetable Garden
YE Jing1, WANG Yixiang1,2*, LIU Cenwei1, LIN Yi1, HUANG Jiaqing1, WENG Boqi1
(1. Agricultural Ecology Institute, Fujian Academy of Agricultural Science, Fujian Key Laboratory of Agricultural Ecological Process of Red Soil Mountain, Fuzhou 350013, China; 2. Institute of Soil and Fertilizer,Fujian Academy of Agricultural Science,Fuzhou 350013, China)
In order to understand the field long-term effect of biochar-based fertilizer instead of chemical fertilizer, the vegetable yield, soil physical and chemical properties and bacterial community structure with biochar-based fertilizer replacement for 5 consecutive years were studied by localization experiment. The results showed that soil pH increased 0.13-0.25 after 5 years, and the contents of organic matter, alkali-hydrolyzable nitrogen and available phosphorus in soils increased 2.1%-62.2%, 5.8%-86.0% and 0.4%-103.1%, respectively. The yield ofwith biochar-based fertilizersincreased by 4.0%-14.8% than that of the control, but the yield under 75% replacement was lower than that of 50% replacement. The indexes of Sobs, Shannon, Ace and Chao of soil bacteria with biochar-based fertilizer were higher than those of chemical fertilizer, and those under 75% replacement were the highest. Compared to chemical fertilizer, the abundances ofandwith biochar-based fertilizer decreased significantly, and the abundances of cellulose-degrading bacterial increased, such asand. There were significant correlations between the abundances ofIlumatobacteraceae, Methyloligellaceae, and soil total nitrogen, total phosphorus, organic matter. Therefore, the replacement of continuous biochar-based fertilizer significantly improved soil physical and chemical properties, and caused the changes in bacterial community structure and diversity. Appropriate replacement of biochar-based fertilizers could increase vegetable yield, but the effect of long-term continuous application on soil pH and C/N should be paid attentions, so as to avoid excessive application inhibiting nutrients uptake and utilization by crops.
Vegetable garden; Biochar-based fertilizer; Chemical fertilizer; Soil; Bacterial community
10.11926/jtsb.4624
2022-02-04
2022-05-27
中央引导地方科技发展专项(2021L3021);福建省科技厅公益项目(2020R1021003);福建省农科院协同创新工程(XTCXGC2021010)资助
This work was supported by the Project for Central Government Guides Local Science and Technology Development (Grant No. 2021L3021), the Project for Public Welfare Scientific Research in Fujian (Grant No. 2020R1021003), and the Project for Collaborative Innovation of Fujian Academy of Agricultural Sciences (Grant No. XTCXGC2021010).
叶菁(1985年生),女,助理研究员,研究方向为红壤保育与生态修复。E-mail: anna_0266@163.com
通讯作者 Corresponding author.E-mail: sd_wolong@163.com
猜你喜欢
中国化肥信息(2022年9期)2022-11-25
林业与环境科学(2022年5期)2022-02-02
今日农业(2021年19期)2022-01-12
今日农业(2021年19期)2021-11-27
今日农业(2021年4期)2021-06-09
中国化肥信息(2018年2期)2018-08-23
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
现代检验医学杂志(2015年1期)2015-02-06
特产研究(2014年4期)2014-04-10
