
巴氏新小绥螨对同一栖息地枸杞瘿螨的捕食功能反应
2023-08-01 08:24:56曹源智贾永红付晓彤李玉晶孟瑞霞
环境昆虫学报 2023年3期
曹源智,贾永红,付晓彤,李玉晶,孟瑞霞
(内蒙古农业大学园艺与植物保护学院,呼和浩特市 010019)
枸杞LyciumbarbarumL.是我国重要的传统药材与滋补食品(Amagase and Farnsworth,2011;杨孟可,2020;王莹等,2022)。然而,由于枸杞的嫩叶、嫩枝和果实水分充足营养丰富,且营养生长与生殖生长几近同时发生,所以极易遭受虫害的威胁(陈君等,2003;刘爱萍等,2007;徐常青等,2014)。其中,枸杞瘿螨AceriapallidaKeifer是枸杞上为害最严重的有害生物之一(徐林波和段立清,2005;刘爱萍等,2007;徐常青等,2014;严海霞和郭洁,2019;杨宁权等,2021),并因形成虫瘿,导致整株枸杞长势衰弱,果实减产,严重影响着我国枸杞的产量与质量(容汉诠和王华荣,1983;徐林波和段立清,2005;孙学海,2007;刘赛等,2019)。
枸杞瘿螨由于体型微小,大部分又潜藏于虫瘿中为害,在生产中有效控制枸杞瘿螨的手段并不多(刘赛等,2016;杨宁权等,2021),现在仍主要使用化学杀螨剂等进行防治(李建领等,2017;马莹和张文华,2019)。大量不规范施用化学农药防治枸杞瘿螨现象非常普遍(徐林波和段立清,2005;徐常青等,2014),引起枸杞农药残留超标及出口海外受阻(徐常青等,2014;李建领等,2017)。因此,寻求枸杞瘿螨的有效天敌而加快生物防治等绿色防控技术的研发,对保障枸杞质量安全以及枸杞产业健康发展都有深远影响。在本地生态系统中,本土天敌是一个固定的生物因子,调查筛选害虫发生地的本土天敌种类,增加本土天敌的保护与利用,可不必考虑引进天敌所必需经历的风险评估和对新环境的适应等问题(郭同斌,2020)。因此,开发利用本土天敌种类是害虫生物防治中最安全有效的途径。
刘俊清等(2019)于2017-2018年对内蒙古自治区巴彦淖尔市乌拉特前旗野生枸杞地的植绥螨进行了调查,表明有3种植绥螨与枸杞瘿螨可同时发生在野生枸杞上,包括巴氏新小绥螨NeoseiulusbarkeriHughes、安德森钝绥螨AmblyseiusandersoniChant和新小皱新小绥螨NeoseiulusneoreticuloidesLiang &Hu,并且在已经破裂成熟的枸杞瘿螨瘿瘤里可发现各个发育阶段的植绥螨。Fu等(2021)研究了其中两种植绥螨安德森钝绥螨和新小皱新小绥螨捕食枸杞瘿螨的功能反应,表明安德森钝绥螨有潜力成为防治枸杞瘿螨的生防作用物。另外一种植绥螨巴氏新小绥螨对各种小型害虫害螨均有较好的防治效果,并已实现了大规模化饲养与商业化生产(徐学农等,2013),广泛应用于我国的农业生防实践中(Wuetal.,2017),而且巴氏新小绥螨也是内蒙古地区的捕食螨优势种(张鹏飞等,2012)。国外发现巴氏新小绥螨可控制椰子瘿螨AceriaguerreronisKeifer(Limaetal.,2015),但巴氏新小绥螨对枸杞瘿螨捕食潜力的研究国内外还未见报道。
功能反应是评估捕食者捕食率和捕食者控制有害生物种群能力的重要参数(Skalski and Gilliam,2001;Songetal.,2016;De Sousa Netoetal.,2019)。在捕食者与猎物的系统中,功能反应提供了关于天敌对特定害虫的控制效率的数据(Rahmanetal.,2012),可用于预测捕食者与猎物之间互作行为的潜在机制,以提高捕食者控制猎物的潜力(Sepúlveda and Carrillo,2008;Delong and Uiterwaal,2022)。本研究利用功能反应,评价了来自同一栖息地的巴氏新小绥螨雌、雄成螨对枸杞瘿螨的捕食潜力,以期为开发枸杞瘿螨的本地有效天敌进行生物防控提供依据。
1 材料与方法
1.1 供试虫源
巴氏新小绥螨于2017年9月采自巴彦淖尔市野生枸杞地,以甜果螨Carpoglyphuslactis饲养在25±1℃、RH 65%±10%、16 L∶8 D的智能人工气候箱中,并成功建立稳定种群。
枸杞瘿螨与巴氏新小绥螨为同一地点采集,将带有枸杞瘿螨虫瘿的枸杞枝条带回室内后,放置于4℃的冰箱中待用。
1.2 试验装置
试验装置为由三层透明亚克力板(35 mm×20 mm×3 mm)、一层枸杞叶片和一层滤纸组成的小室。三层亚克力板中,其中有两层亚克力板的中心有直径为10 mm的圆孔,枸杞叶片与滤纸大小与亚克力板的尺寸相当。小室从上到下依次由完整的亚克力板、中心开孔的亚克力板、叶背向上的枸杞叶片、滤纸及中心开孔的亚克力板组成,并用燕尾夹夹紧完整小室的两端。随后将其放置在塑料托盘上,托盘上铺有湿纱布用以保湿。
1.3 试验方法
枸杞瘿螨可分为4个发育阶段,分别为卵、幼螨、若螨和成螨。其卵(近球形,直径为39~42.5 μm)、幼螨(体长74~109.6 μm)和若螨体型微小(容汉诠和王华荣,1983),成螨也只有200~240 μm(匡海源,1983)。卵在挑取的过程中极易破损,而幼螨、若螨极易钻入小室亚克力板与叶片之间的缝隙中而躲避巴氏新小绥螨的捕食,因此本研究只针对枸杞瘿螨成螨进行了实验。
巴氏新小绥螨用枸杞瘿螨瘿瘤块饲养3代后用于试验。由于未成熟的枸杞瘿螨瘿瘤几乎是没有缝隙的,所以用小刀将直径为2~5 mm的枸杞瘿螨瘿瘤从枸杞叶片切下,并沿瘿瘤中央剖开,使瘿瘤中的枸杞瘿螨完全暴露于捕食螨,以利于捕食螨进行捕食。为使巴氏新小绥螨的发育历期保持一致,在种群中随机挑取150头巴氏新小绥螨已交配雌成螨,待其产卵24 h后将雌成螨去除,在卵孵化以后继续用枸杞瘿螨饲养直至开始实验。将其中14日龄(确保捕食螨已交配)的巴氏新小绥螨雌、雄成螨随机各挑取80头单独放置于实验小室中饥饿24 h,再将枸杞瘿螨成螨挑入小室,其密度分别设为30、50、70、90、110、130、150、170头,24 h后记录被捕食猎物的数量,每个密度处理重复10次。所有小室都放置在25±1℃、RH 65%±10%、16 L∶8 D的智能人工气候箱(PRX-350C,宁波海曙赛福实验仪器厂)中。
1.4 数据分析
功能反应类型采用Logistic模型来确定功能反应的类型。
Ne/N0=exp(P0+P1N0+P2N02+P3N03)/[ 1+exp(P0+P1N0+P2N02+P3N03)]
(1)
在等式(1)中,Ne代表被捕食猎物的数量,N0代表初始猎物数量,(Ne/N0)为猎物被捕食的概率。P0为截距,P1、P2、P3分别为线性系数、二次系数和三次系数(Xiao and Fadamiro,2010)。功能反应的类型由线性系数和二次系数的符号确定,当线性系数为负值时,捕食者表现出II型功能反应;当线性系数为正,二次系数为负时,捕食者表现出III型功能反应(Juliano,2001)。
采用非线性最小二乘回归估计攻击率(α)处理时间(Th)(Rogers,1972;Hassell,1978)。
Ne=N0{ 1-exp [α(ThNe-T)] }
(2)
在等式(2)式中,Ne为被捕食猎物的数量,N0为初始猎物数量,α为攻击率,Th为每个猎物的处理时间,T为实验时间。
根据等式(2)所得出的攻击率α与处理时间Th计算搜寻效应S。
S=α/(1+αThN0)
(3)
在等式(3)式中,S为搜寻效应,α为攻击率,N0为初始猎物数量,Th为每个猎物的处理时间。
采用SAS 9.4软件,通过Logistic模型(等式1)确定巴氏新小绥螨雌、雄成螨对枸杞瘿螨成螨的捕食功能反应类型,采用非线性最小二乘回归(等式2)估计攻击率和处理时间,并根据(等式3)计算搜寻效应。采用单因素方差分析以及Duncan检验比较不同猎物密度对巴氏新小绥螨捕食量的影响,并应用独立样本t检验比较巴氏新小绥螨不同性别对其捕食量的影响。使用GraphPad Prism 8绘制功能反应曲线及搜寻效应曲线。
2 结果与分析
2.1 巴氏新小绥螨对枸杞瘿螨成螨的捕食功能反应
枸杞瘿螨的密度对巴氏新小绥螨雌、雄成螨的捕食量均有显著影响(雌成螨:F7,64=212.304,df=7,P<0.05;雄成螨:F=21.267,df=7,P<0.05),巴氏新小绥螨雌、雄成螨对枸杞瘿螨的捕食量均随着猎物密度的增加而增加,当瘿螨密度达到一定数量时,巴氏新小绥螨雌、雄成螨的捕食量逐渐趋于稳定(表1)。巴氏新小绥螨雌、雄成螨对枸杞瘿螨的捕食量通过独立样本t检验后,结果表明巴氏新小绥螨雌成螨的捕食量均显著大于雄成螨(P<0.05)(表1)。

表1 巴氏新小绥螨对枸杞瘿螨的捕食量(平均值±标准误)Table 1 Number of Aceria pallida consumed by Neoseiulus barkeri at different prey density (mean±SE)

图1 巴氏新小绥螨捕食枸杞瘿螨的功能反应曲线Fig.1 Functional response curves of Neoseiulus barkeri fed on Aceria pallida
巴氏新小绥螨雌、雄成螨Logistic回归的线性系数P1分别为-0.6805与-0.0299,皆小于0(表2),表明巴氏新小绥螨雌、雄成螨捕食枸杞瘿螨的功能反应均为II型。
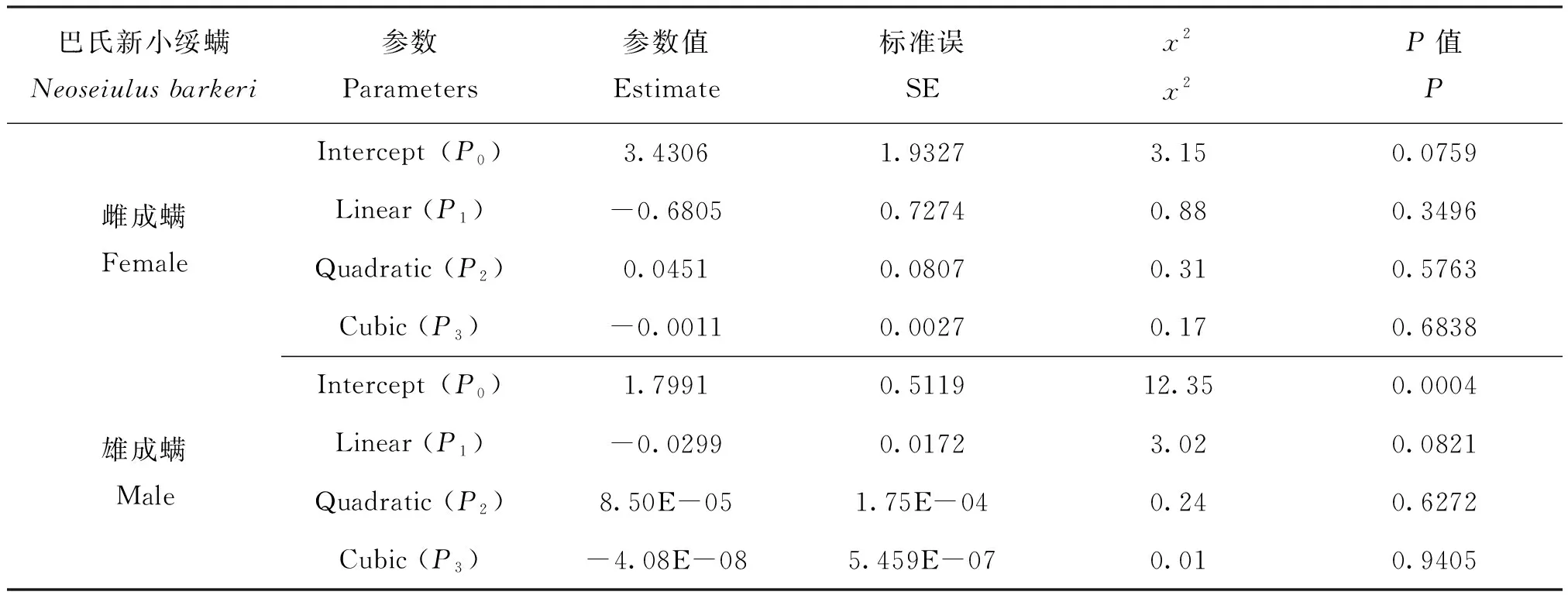
表2 巴氏新小绥螨对枸杞瘿螨功能反应的逻辑回归系数估计Table 2 Maximum-likelihood estimates of the proportion of Aceria pallida consumed by Neoseiulus barkeri as a function of initial prey density from logistic regressions
巴氏新小绥螨雌成螨捕食枸杞瘿螨的攻击率α值(4.4121)高于雄成螨(2.4403),处理时间Th雌成螨(0.0094 d)则短于雄成螨(0.0196 d)(表3)。T/Th为巴氏新小绥螨24 h内捕食枸杞瘿螨成螨的理论最大日捕食量,结果显示巴氏新小绥螨雌成螨对枸杞瘿螨成螨的理论最大日捕食量(106.61头)高于雄成螨的理论最大日捕食量(51.02头),且是雄成螨的2.10倍(表3)。

表3 巴氏新小绥螨对枸杞瘿螨攻击率以及处理时间Table 3 Attack rate and handling time for Neoseiulus barkeri feeding on Aceria pallida
2.2 巴氏新小绥螨对枸杞瘿螨成螨的搜寻效应
巴氏新小绥螨雌、雄成螨对枸杞瘿螨成螨的搜寻效应随着猎物密度的增加而减小,并且雌成螨在所有密度条件下的的搜寻效应皆大于雄成螨(图2)。当在低猎物密度30头时,巴氏新小绥螨雌、雄成螨的搜寻效应均达到最大(雌成螨:1.97;雄成螨:1.00);当猎物密度达到170头时,巴氏新小绥螨雌、雄成螨的搜寻效应均最低(雌成螨:0.55;雄成螨:0.27)(图2)。

图2 巴氏新小绥螨对枸杞瘿螨的搜寻效应Fig.2 Searching efficiency of Neoseiulus barkeri fed on Aceria pallida
3 结论与讨论
本研究测定了本地巴氏新小绥螨对同一栖息植物上枸杞瘿螨的捕食功能反应,表明巴氏新小绥螨雌、雄成螨捕食枸杞瘿螨均表现为Holling-II型功能反应。这与在同一栖息地采集到的安德森钝绥螨、新小皱新小绥螨捕食枸杞瘿螨成螨也表现为Holling-II型功能反应是一致的(刘庆岩,2019;Fuetal.,2021)。在众多植绥螨种类中,Holling-II型功能反应较为常见,如拉哥钝绥螨AmblyseiuslargoensisMuma捕食荔枝瘤瘿螨AcerialitchiiKeifer(程立生等,2005)、加州新小绥螨NeoseiuluscalifornicusMcGregor捕食二斑叶螨TetranychusurticaeKoch(Netoetal.,2019)、拉哥钝绥螨若螨和雌成螨捕食印度雷须螨RaoiellaindicaHirst(Liraetal.,2021)及条纹新小绥螨NeoseiulusstriatusWu捕食二斑叶螨各螨态(曾科科,2021)。巴氏新小绥螨对害螨猎物的捕食也大多表现为Holling-II型功能反应,Li等(2018)测定了巴氏新小绥螨捕食乱跗线螨TarsonemusconfususEwing不同发育阶段的功能反应,对乱跗线螨卵和静止期的幼螨表现为Holling-II型功能反应类型;巴氏新小绥螨捕食二斑叶螨各个发育阶段的功能反应也均为Holling-II型(王蔓等,2019)。Fathipour等(2021)研究了补充食物玉米花粉ZeamaysL.对巴氏新小绥螨捕食二斑叶螨的影响,表明无论花粉存在与否,巴氏小绥螨对二斑叶螨的卵和若螨都表现Holling-II型功能反应。此外,巴氏新小绥螨对其他猎物的捕食功能反应也表现为Holling-II型,如巴氏新小绥螨对水稻干尖线虫AphelenchoidesbesseyiChristie的捕食(李曼,2019);巴氏新小绥螨对芒果茶黄蓟马ScirtothripsdorsalisHood 1龄若虫与2龄若虫的捕食(周炀等,2020)。
攻击率和处理时间是确定功能反应大小的重要指标,通常认为高攻击率(α)和短处理时间(Th)的天敌是更有效的害虫生防作用物(Pervez and Omkar,2006;Fathi and Nouri-Ganbalani,2010;Salehietal.,2016)。Fu等(2021)研究了与巴氏新小绥螨采自同一野生枸杞地的安德森钝绥螨对枸杞瘿螨成螨的捕食功能反应,其设定温度、小室内空间大小和捕食螨饥饿程度皆与本实验相同。安德森钝绥螨雌成螨捕食枸杞瘿螨成螨时,其处理时间(Th=0.0140 d)长于巴氏新小绥螨雌成螨(Th=0.0094 d),即使其攻击率(α=5.961)大于巴氏新小绥螨雌成螨(α=4.4121),但安德森钝绥螨雌成螨对枸杞瘿螨成螨的理论最大日捕食量(71.43头)小于巴氏新小绥螨雌成螨(106.61头)。安德森钝绥螨雌成螨在枸杞瘿螨成螨密度为100头时的捕食量为57.6头;而巴氏新小绥螨雌成螨在枸杞瘿螨成螨密度为90头时的捕食量就已经达到72.4头,远大于安德森钝绥螨雌成螨的捕食量。因此,虽然与枸杞瘿螨采自同一栖息植物的两种本地植绥螨-巴氏新小绥螨和安德森钝绥螨都适宜作为枸杞瘿螨的生防作用物,但巴氏新小绥螨对枸杞瘿螨表现有更好的捕食潜力,对枸杞瘿螨具有更快找到并消耗的能力。
Delong和Uiterwaal(2022)整理了来自世界各地以植食性害螨和蚜虫为猎物的400多个功能反应,重新分析使之标准化,发现温度、捕食者的体型大小、试验装置内空间的大小和捕食者饥饿程度都对其捕食猎物的功能反应的曲线高度和形状起着作用,进而会影响功能反应的各项参数。
本研究中的试验温度、试验小室空间和捕食者饥饿程度都是恒定的,只有巴氏新小绥螨雌、雄成螨在体型大小上有差异,雌成螨的体型大于雄成螨。一般来说,捕食者体型越大,捕食者对于相同空间里猎物的攻击率就越强(Delong and Uiterwaal,2022)。捕食者的体型大小还与搜索速度、探测距离或捕食猎物的能力相关,进而影响捕食者对猎物的处理时间(Delong and Uiterwaal,2022)。较小的捕食者由于其从更远的距离感知猎物的能力相对较弱,从而会花费更长的时间去寻找猎物(McGill and Mittelbach, 2006)。巴氏新小绥螨雌成螨捕食枸杞瘿螨成螨时表现出较高的攻击率α(雌成螨:4.4121;雄成螨:2.4403)和较短的处理时间Th(雌成螨:0.0094 d;雄成螨:0.0196 d)。在Fu等(2021)的研究中也有类似结果,安德森钝绥螨在捕食枸杞瘿螨成螨时,雌成螨比雄成螨有更高的攻击率和更短的处理时间,这均应为捕食螨雌、雄成螨的体型大小差异所导致的结果。
搜寻效应是捕食者在取食猎物过程所中表现的一种行为效应,其大小与猎物密度、攻击率和处理时间相关,搜寻效应会随着猎物密度的增加而降低(丁岩钦,1994)。在本研究中,巴氏新小绥螨雌、雄成螨捕食枸杞瘿螨成螨时也呈现出这种规律;巴氏新小绥螨在捕食水稻干尖线虫和芒果茶黄蓟马时也有相同规律(李曼等,2019;周炀等,2020)。然而,巴氏新小绥螨在捕食枸杞瘿螨成螨时其搜寻效应的最大值,皆大于巴氏新小绥螨捕食水稻干尖线虫和芒果茶黄蓟马的最大值(李曼等,2019;周炀等,2020),这可能是因为枸杞瘿螨成螨的体型过于微小,为防止枸杞瘿螨逃逸,本研究只能选择小型实验装置,而实验装置内空间的大小会影响捕食者对猎物的攻击率α和处理时间Th(Delong and Uiterwaal,2022),进而导致巴氏新小绥螨在捕食枸杞瘿螨成螨的搜寻效应大于巴氏新小绥螨捕食水稻干尖线虫和芒果茶黄蓟马的搜寻效应。
随着害虫生物防治技术的普及,人们也担心生物防治产品是否会影响环境和生态等问题,如天敌产品是否会降低施用地的生物多样性(暴可心等,2021),这样,本土天敌的开发与利用就显得尤为重要。本土天敌避免了引进天敌可能带来的风险,相对引进天敌也更容易定殖于目标植物,而且本土捕食者可能对其同一栖息地的有害生物具有更强的防控能力(郭同斌,2020)。Mendes等(2018)的研究表明,对目标猎物印度雷须螨已有捕食经验的拉哥钝绥螨对猎物捕食的处理时间更短,而且本土拉哥钝绥螨种群可能对目标猎物有更强的捕食能力。Patel and Zhang(2017)比较了两种植绥螨AmblydromaluslimonicusMuma和胡瓜新小绥螨NeoseiuluscucumerisOudemans捕食马铃薯/番茄木虱BactericeracockerelliSulc的功能反应,结果显示与马铃薯/番茄木虱采自同一温室辣椒植株上的植绥螨A.limonicus比商品化的胡瓜新小绥螨对猎物具有更高的攻击率以及更短的处理时间,表明与猎物来自同一栖息植物的A.limonicus是比商品化的胡瓜新小绥螨更有效的防控马铃薯/番茄木虱的生防作用物。本研究结果也显示了本土巴氏新小绥螨对同一栖息地植物上的枸杞瘿螨具有较好的捕食潜力,有望成为防控枸杞瘿螨的有效生防作用物。
此外,捕食者的捕食量和功能反应还可能与温度相关(Uszkoetal.,2017;Islametal.,2020)。功能反应的各项数据会随着平均环境温度的变化而改变,温度升高可能会降低捕食者的处理时间,这可能会使生物防治更有效(Delong and Uiterwaal,2022);但如果超过一定温度,捕食者因自身代谢速率过快,能量消耗过多,对猎物的寻找效率也会降低(Uiterwaal and DeLong,2020)。随着全球气候的变暖,当应用天敌对有害生物进行防治时,也应关注温度对捕食者的影响。因此,还有待进一步开展田间实际条件下的释放试验研究,以期为利用本土巴氏新小绥螨防控枸杞瘿螨提供依据。
猜你喜欢
中学生天地(A版)(2022年9期)2022-10-31 06:36:58
中国果业信息(2021年7期)2021-12-01 20:20:32
云南大学学报(自然科学版)(2021年1期)2021-02-05 08:04:04
幼儿100(2018年34期)2018-12-29 12:31:42
太原师范学院学报(自然科学版)(2018年2期)2018-08-17 12:21:50
东华大学学报(自然科学版)(2018年1期)2018-06-29 03:35:24
中国乳业(2016年4期)2016-11-07 09:50:29
中外文摘(2016年13期)2016-08-29 08:53:27
天津科技大学学报(2016年1期)2016-02-28 16:59:44
英语学习(2015年5期)2016-02-02 01:10:27
