
小贯小绿叶蝉(Empoasca onukii)的生长发育对自身视觉能力的影响
2023-07-03 07:02:14谭畅单瑶周孝森姚秋怡蔡晓明边磊
茶叶科学 2023年3期
谭畅,单瑶,2,周孝森,姚秋怡,蔡晓明,边磊*
1. 中国农业科学院茶叶研究所,浙江 杭州 310008;2. 河南科技学院资源与环境学院,河南 新乡 453003
视觉信息是昆虫搜索食物、定位寄主和飞行导航等行为的重要线索,昆虫获取的视觉信息质量会受到自身复眼视觉能力的限制[1]。随着昆虫的生长发育,其复眼的尺寸和结构会发生显著变化,如一些蚁类和黄蜂复眼上小眼的数目会逐渐增加[2-4],小眼直径增大[5-7]。复眼尺寸增加可以提高昆虫识别视觉图像的质量,小眼数目的增加能提高视觉分辨率[8],小眼内光受体的生长可以提高其光灵敏度[9]。
昆虫复眼的生长与体型的生长存在异速差异,复眼和体型的比例会随着个体的生长发育逐渐缩小[6]。生物体的异速生长规律符合幂函数[10],部分器官与主体特征之间异速生长的关系可以用来预测器官在未来的变化和分析昆虫对环境的适应策略[11]。异速生长指数和初始生长指数是衡量生物体器官异速生长的重要参数,通过参数之间的对比可以分析器官在功能上的变化特征, 如沙漠蚂蚁(Cataglyphis albicans、Cataglyphis bicolor和Cataglyphis fortis)和黑腹果蝇(Drosophila melanogaster)复眼上小眼间角的异速生长指数(-0.40 和-0.21)均低于小眼数目的异速生长指数(0.75 和0.58)[3,12],说明这些昆虫复眼的视敏度会随着体型的增大而提高。异速生长还可以表现在复眼的不同区域上,使这些区域分化为不同的特异性功能区[13],如苜蓿黄蝶(Colias eurytheme)羽化后个体之间在体型上存在差异,个体间复眼小眼间角的异速生长指数在不同区域也存在差异[14],说明个体体型会影响苜蓿黄蝶复眼上特异性功能区的视觉能力。因此,联系昆虫复眼参数的异速生长和视觉能力的变化特征,对衡量昆虫的生长发育是否伴随着视觉能力的提升是必要的。
小眼间角是衡量并置复眼视觉分辨率的重要参数,可通过组织学方法直接测定区域小眼的理论视敏度[15-16]。除此之外,通过观测昆虫区分垂直光栅或水平光栅所做出的行为反应[17-19],明确昆虫能区分的最细光栅条纹,同样可以从行为上评估目标昆虫复眼的分辨率[20-21]。组织学方法得到的理论值可以提供复眼视敏度的具体范围,由于昆虫习性、光学像差和空间因素的影响,行为学测试得到的复眼分辨率通常低于组织学方法测定的理论上限[22-23],但却能更好地代表昆虫整体的视觉分辨率[24-25]。复眼的灵敏度是指光感受器捕获光子的能力,与区域内的小眼直径、小眼接受角和视杆长度相关[26],由于这些因素都受复眼体积的制约,所以复眼的光学灵敏度与复眼的大小密切相关。
小贯小绿叶蝉(Empoasca onukii)是茶树上的重要害虫,属于不完全变态昆虫,若虫有5 个龄期[27],每次蜕皮后体型增大,复眼尺寸和小眼数量明显增加。为了探明叶蝉的生长发育对其视觉能力的影响,本研究采用组织学和行为学方法,测定不同龄期叶蝉的体型大小、复眼的光学灵敏度和视觉敏锐度,并采用异速生长函数分析复眼视觉能力的异速生长特征,最后通过模拟叶蝉的视觉能力评估其生长发育过程中有效视距的变化,以期为深入研究叶蝉的行为生态学和研发无公害防治的技术提供理论基础。
1 材料与方法
1.1 试验昆虫
小贯小绿叶蝉采自中国农业科学院茶叶研究所周边茶园,采用龙井43 水培茶枝饲养。养虫室温度为24~27 ℃,相对湿度为65%~75%,光周期为14L∶10D。茶枝上叶蝉的卵孵化后,采集5 个龄期的若虫和羽化1 d后的雌雄成虫作为组织学和行为学试验样本。
1.2 叶蝉复眼的外部结构分析
采集试虫低温麻醉后,用橡皮泥固定在观测平台上,采用超景深3D 显微镜(基恩士,VHX-6000,日本)搭载镜头VH-Z100 UR 在(500~700)×放大倍数下,扫描样本的背侧整体形态和单个复眼的3D 表面结构。各龄期若虫与雌雄成虫的生物学重复为7 次。
通过超景深3D 显微镜的分析软件测量试虫前胸背板宽度(Inter tegular width,ITW,图1A),复眼上小眼的数量(N),中部区域的小眼间角(Δϕ)、小眼直径(D)和视杆长度(L,图2A 和图2B),其中Δϕ为所测小眼3 个方向上小眼间角的平均值,D为所测小眼垂直与水平两个方向角膜直径的平均值。

图1 叶蝉若虫和雌雄成虫背侧图Fig. 1 Dorsal view of nymph and adult of E. onukii

图2 叶蝉复眼内结构参数的测量Fig. 2 Measurement of structural parameters in compound eye of E. onukii
1.3 叶蝉复眼的内部结构分析
将采集试虫低温麻醉后取下头部,放入4%多聚甲醛中固定0.5 h,用0.1 mol·L-1pH 7.4的PBS 缓冲液冲洗3~5 次,每次冲洗10 min,70%、85%、95%梯度酒精分别脱水15 min 后,100%酒精脱水2 次,每次5 min,接着过渡至2∶1 和1∶1 的二甲苯酒精稀释溶液,分别处理 20 min,然后采用梯度正丁醇溶液进行处理。最后用二甲苯处理10~20 min。二甲苯与液态石蜡分别以2∶1 和1∶1 混合均匀加入待浸蜡组织,真空负压条件下浸蜡30 min 后,转入纯石蜡溶液中浸蜡1~1.5 h。浸蜡组织包埋后进行修块,通过超薄切片机(Leica EM UC7)获得厚3 μm 的复眼半薄切片,苏木精-伊红染色后,中性树脂胶封片。
采用超景深3D 显微镜对制作好的复眼切片进行观测和拍摄,放大倍数为(800~1 000)×。通过ImageJ(V1.8.0,美国)测量复眼中部区域内视杆的长度L(图2C),L为区域内5个视杆长度的平均值。各龄期若虫与雌雄成虫的生物学重复为3 次。
1.4 行为试验
小贯小绿叶蝉的复眼属于并置复眼[28],其分辨率的高低在行为上表现为可以远距离识别明暗光栅的能力[21]。叶蝉若虫对环境的光强度极为敏感,并对黑色表现出明显的偏好性及趋性运动。若虫的行为试验设计参考Kirwan 等[21]的方法,设计明暗相间的黑白光栅作为叶蝉若虫的刺激(吸引)目标,通过行为反应测定不同龄期叶蝉若虫的视敏度。
行为测定装置如图3A 所示,圆形反应室直径15 cm、高度2 cm,反应室下方放置辐射度为425 µw·cm-2的发光板,该亮度为晴朗天气时下午16:00—17:00 叶蝉活跃时间段[29],芽下第二叶处的光照强度。反应室侧面背景为灰色,刺激目标为明暗相间的黑白光栅(图3B),暗区呈黑色(反射光强度最低),两侧逐渐过渡至白色(反射光强度最高)后再过渡至灰色,反射光强度的变化符合正弦函数[21],黑白光栅的平均反射光强度等于灰色背景的反射光强度(图3C)。每次行为测试之前,反应室使用无水乙醇擦洗并晾干。
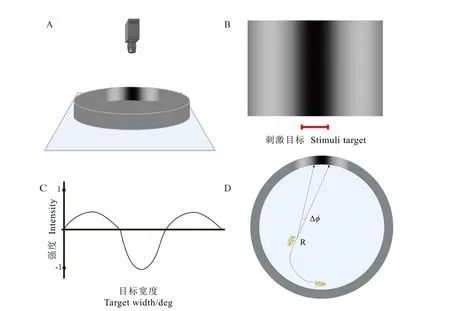
图3 小贯小绿叶蝉若虫的行为试验装置Fig. 3 Apparatus for the behavioral experiment of E. onukii nymphs
试虫暗适应10 min 后,在反应室距离光栅的最远端释放(图3D),观测若虫的运动轨迹并通过反应室上方的高速摄像机(Reverler,中国)记录。行为测试的时间上限为10 min,超过10 min 则认定为若虫对光栅无反应。每次试验重复时将反应室随机旋转90°,避免光栅方向产生的影响。每个龄期的若虫各重复15 只。
行为轨迹采用视频分析软件 Tracker(V6.0.8,美国)进行分析,将叶蝉若虫产生趋性反应的起始位置标记为识别点 R,点 R与黑色刺激区域之间的夹角即为若虫的小眼间角(图3D)。不同龄期若虫的行为学小眼间角Δϕ’取各重复试验的平均值。
1.5 数据分析
1.5.1 视觉敏锐度和光学灵敏度
复眼的视觉敏锐度(A)为小眼间角(Δϕ)的倒数,单位为cpd(Cycles per degree,周/度);视距评估中采用行为试验得到的小眼间角(Δϕ’)进行计算。
复眼的光学灵敏度(S)采用公式(2)计算,单位为μm2·sr。
式中,D为小眼直径,Δϕ为组织学试验得到的小眼间角,L为视杆长度,k值(0.006 7 μm-1)为节肢动物光受体的光吸收系数[26]。
1.5.2 复眼的异速生长规律
复眼的异速生长符合幂函数方程,
式中,Y为表征复眼不同特征的变量,包括N、D、Δϕ、L,x为表征叶蝉生长指标的变量ITW,a为复眼不同特征变量的异速生长指数,b为该变量的初始生长指数[10]。
1.5.3 有效视距评估
AcuityView 是R 语言中评估生物体有效视距的常用模型[30]。可通过生物体的视觉敏锐度、目标图像和不同观测距离,生成以生物体视觉敏锐度为标准的不同距离下观察到的目标图像。以叶蝉视觉敏锐度为标准,以与实物大小一致的茶梢图像(10 cm×10 cm,分辨率为1 024×1 024)为识别目标,采用R(4.2.0)生成不同龄期叶蝉识别茶梢的静态图像,模拟距离分别为2、5 cm 和10 cm,最后通过生成图像的清晰度评估试虫对茶梢的有效视距。
1.5.4 差异显著性分析
采用SPSS 19.0 软件进行数据分析,其中不同龄期叶蝉的前胸背板宽度和复眼的视觉参数采用单因素方差分析(ANOVA 和Duncan)进行差异显著性检验;前胸背板宽度和复眼的视觉参数采用幂函数(公式3,2个参数)进行线性拟合。
2 结果与分析
2.1 不同龄期叶蝉复眼的形态变化
随着龄期的增长,叶蝉复眼的整体形态和小眼形状发生显著变化。复眼整体形态由近似椭圆形变为肾形,1~3 龄若虫复眼中部的小眼呈圆形,排列松散且密度较小,每个小眼周围毗邻4 个小眼;4 龄之后,复眼中部的小眼呈六边形,排列紧密,每个小眼周围有6 个小眼(图4)。

图4 各龄期小贯小绿叶蝉复眼的形态Fig. 4 External morphology of compound eyes of E. onukii in different instars
2.2 不同龄期叶蝉复眼视觉参数的异速生长
小贯小绿叶蝉在生长发育过程中,每次蜕皮后其体型和各视觉参数均会发生显著性变化。叶蝉的前胸背板宽度(体型)随着生长发育会逐渐增大(F=204.6,df=6,P<0.05,图1),复眼的小眼数目(F=483.62,df=6,P<0.05,图4)和小眼直径显著增长(F=36.94,df=6,P<0.05),小眼间角持续减小(F=22.3,df=6,P<0.05),即叶蝉复眼的视敏度会随龄期增长而提高。叶蝉复眼的视觉参数符合典型的异速生长规律,其中复眼视敏度(a=-0.65<1)和小眼直径(a=0.35<1)的增长慢于叶蝉体型的增速,小眼数目(a=1.37>1)的增速比体型高(表1),即叶蝉在生长过程中视觉能力的提升主要依靠增加小眼的数目和增大复眼的体积来实现。

表1 各龄期叶蝉的前胸背板宽度、视觉参数(mean±SE)及其异速生长参数Table 1 The width of the pronotum, visual parameters (mean±SE) and their allometric growth parameters for each instar of E. onukii
2.3 不同龄期叶蝉复眼的光学灵敏度和视觉敏锐度
随着叶蝉龄期增加,复眼中部区域的视杆长度显著增长(F=278.59,df=6,P<0.05,图5A),同样符合异速生长规律(a=1.22,b=0.036 ,R2=0.924) , 从 1龄若虫的(17.51±0.42)μm 增长至5 龄的(63.06±0.88)μm,待若虫羽化后,雌雄虫之间视杆长度存在显著性差异(雌虫L=86.46 μm,雄虫L=78.14 μm,P<0.05)。伴随复眼中视杆长度的增长,其光学灵敏度从1 龄若虫的0.04 μm2·sr 逐步提升至 5 龄的 0.11 μm2·sr,符合异速生长规律(a=0.88,b=4.47×10-4,R2=0.822,P=0.034);但若虫羽化后复眼视觉灵敏度降低,雌虫复眼的灵敏度显著低于雄虫(雌虫S=0.069 μm2·sr,雄虫S=0.08 μm2·sr,P<0.05),成虫复眼光学灵敏度的下降导致其不再符合异速生长规律(R2=0.467,P=0.091,图5B)。

图5 各龄期叶蝉复眼中部区域视杆的长度和光学灵敏度的异速生长特征Fig. 5 The rhabdom lengths of each instar and the allometric growth characteristics of optical sensitivity in the center partial of compound eye of E. onukii
行为试验中,空白对照内若虫的行为轨迹无方向性,呈曲线和螺旋状(图6A);存在刺激光栅的情况下,叶蝉若虫释放后会在反应室内随机运动,待若虫探测到刺激光栅后,开始进行无往返的趋向运动(图6B~F),并且所有龄期的若虫在行为试验中均对黑色光栅呈现出趋向运动,1 龄和2 龄若虫爬行速度缓慢,在搜寻过程中会有短暂的停顿,3 龄后的若虫爬行速度变快,能够很快接触到刺激光栅。通过对行为试验中R 点和刺激光栅的夹角分析,1 龄若虫到5 龄若虫的小眼间角显著降低(F=22.15,df=6,P<0.05),分别为(11.62±0.21)°、(11.48±0.27)°、(10.60±0.24)°、(9.80±0.25)°、(8.90±0.13)°,介于组织学结构得到的小眼间角范围内(表1),叶蝉1~5 龄若虫的视敏度分别为 0.086、0.088、0.094、0.102、0.112 cpd,若虫复眼的视敏度符合异速生长规律(a=0.28,b=0.019,R2=0.925)。

图6 行为试验中若虫的运动轨迹图Fig. 6 Movement behavior trajectories of nymph of E. onukii in behavioral experiments
2.4 不同龄期若虫复眼的有效视距评估
依托行为试验得到的叶蝉小眼间角,模拟叶蝉在0、2、5 cm 和10 cm 下识别的茶梢图像(图7),随着龄期的增大,叶蝉复眼对茶梢的有效识别视距逐渐增加;但是随着观测距离的增大,感知图像的清晰度迅速降低,说明叶蝉若虫对图像细节的分辨能力非常有限,叶蝉若虫的复眼仅具有低分辨率视觉。

图7 各龄期叶蝉复眼对静态图像感知的模拟图Fig. 7 Simulations of viewing distance of each instar of E. onukii
3 讨论
昆虫在生长发育过程中各部位的异速生长是为了适应外界环境进化保留下来的重要的发育特征[31]。小贯小绿叶蝉孵化后,1 龄若虫往往只在其孵化的茶枝上活动,3 龄之后则善爬善跳,活动范围明显扩大[32]。复眼视敏度的逐步提高,使叶蝉可以感知临近叶片或枝干的图像细节,准确地判断跳跃的位点,若虫羽化后开始具备飞行能力,复眼视敏度可达到0.14 cpd 以上,有效视距进一步增大。但是为了优先体型和运动相关器官的生长,叶蝉复眼整体视敏度的增速减缓,只有小眼数目的增速比体型快,表明叶蝉复眼在生长过程中更注重视野的增加,以便于捕捉更大范围内的视觉信号。
相较于视敏度,叶蝉复眼的光学灵敏度在生长过程中虽然也在增加,但并不符合异速生长规律。节肢动物在每次蜕皮后,新的小眼会出现在复眼的前端,导致原有的小眼会向后方移动。复眼后部区域的小眼直径通常很大,由于受到前端小眼的挤压,视杆会继续生长而变长,因此节肢动物在停止生长后,复眼后部区域的小眼光学灵敏度也最高[2],导致复眼不同区域在视觉功能上出现异质化。叶蝉羽化后中部区域的光学灵敏度降低,但是复眼后部小眼的灵敏度显著提升,同时在背侧的复眼形成了高视敏度的“敏锐区”,符合复眼不同区域异速生长形成特殊功能区的策略[13]。如欧洲熊蜂(Bombus terrestris)和皱猛蚁(Rhytidoponera inomata)均是通过在有限的复眼上分化出特殊功能区,满足对应龄期的行为需求[8-9]。雄性长尾管蚜蝇(Eristalis tenax)的复眼上存在高灵敏度的“亮区”,用来在明亮的环境中捕捉猎物或寻找配偶[33]。
值得注意的是,在尚未明确视觉信息到达视神经后处理机制的情况下,组织学测量得到的视觉参数仅仅是基于复眼结构的理论值。一些研究认为复眼视敏度在结构分析和行为测试得到结果差异不大[24-25],而另一些研究则认为行为测试远低于组织学方法[22-23],造成这种现象的原因多是由于复眼的区域化异速生长。由于行为试验建立在昆虫复眼的整体功能上,所以将复眼结构分析与行为试验相结合,能更精确地揭示昆虫视觉在特定环境下的视敏度[34]。虽然叶蝉复眼的视敏度和灵敏度在生长发育后都有显著提升,但是相较于活动范围更大或者飞行能力更强的昆虫,如蜜蜂、蝴蝶和苍蝇的视敏度通常介于0.3~1.0 cpd,蜻蜓、螳螂等捕食性天敌的视敏度在1.0 cpd 以上[35],叶蝉依然属于低视觉分辨率的昆虫。
猜你喜欢
农业工程学报(2022年10期)2022-08-22 07:21:20
天津农业科学(2022年5期)2022-05-31 21:01:11
中国典型病例大全(2022年9期)2022-04-19 21:26:28
今日农业(2021年15期)2021-11-26 03:30:27
山地农业生物学报(2018年2期)2018-06-22 12:42:06
作文周刊·小学六年级版(2016年25期)2017-03-06 18:27:11
四川动物(2017年1期)2017-02-18 07:05:13
长江大学学报(自科版)(2015年21期)2015-12-04 01:34:18
植物保护(2015年4期)2015-11-28 07:28:28
蚕桑茶叶通讯(2015年1期)2015-02-22 15:13:21
