
奶牛乳房炎主要病原菌GapC 蛋白生物信息学分析及抗原表位预测
2023-06-27 11:16:52刘慧婷刘学静宋成祺陈晶张帆高珊佟春玉马金柱崔玉东
黑龙江八一农垦大学学报 2023年3期
刘慧婷,刘学静,宋成祺,陈晶,张帆,高珊,佟春玉,马金柱,崔玉东,
(1.黑龙江八一农垦大学动物科技学院,大庆 163319;2.黑龙江八一农垦大学生命科学技术学院)
奶牛乳腺炎是一种主要由病原菌感染引起的、以乳房红、肿、热、痛为主要临床症状的急性或慢性乳腺炎症,不仅严重危害奶牛健康,也给奶业带来巨经济大损失[1-2]。引起奶牛乳腺炎的病原种类很多,但以革兰氏阳性的链球菌和葡萄球菌为主,包括无乳链球菌(Streptococcus agalactiae)、停乳链球菌(Streptococcus dysgalactiae)、乳房链球菌(Streptococcus uberis)以及金黄色葡萄球菌(Staphylococcus aureus)和表皮葡萄球菌(Staphylococcus epidermidis)[3-4]。奶牛乳腺炎防治一直以抗生素治疗为主,但由于抗生素的广泛持续使用,导致了病原菌耐药性不断增强和抗生素疗效果不佳甚至无效。因此,新型疫苗研发已成为奶牛乳腺炎防治工作重点[5-8]。但由于引起奶牛乳腺炎的病原菌种类繁多,进而可选择作为疫苗的抗原更加繁多,不仅给疫苗研制和使用带来不便,而且研制的疫苗也不具有免疫预防的广谱性。所以寻找奶牛乳腺炎主要病原菌的共同抗原来研制新型疫苗是非常值得探索的课题。
甘油醛-3-磷酸脱氢酶(Glyceraldehyde-3-phosphate dehydrogenase,GAPDH)是一种广泛存在于原核生物和真核生物的参与糖酵解的关键酶[9-12]。甘油醛-3-磷酸脱氢酶C (也常被缩写成GapC)是GAPDH 的一个亚单位,具有高度的保守性,且具有甘油醛-3-磷酸脱氢酶活性、转铁蛋白结合活性、纤维蛋白溶酶活性等[13-16]。以往研究显示,包括S.agalactiae和S.aureus的多种病原菌GapC 都具有良好的免疫原性,能够诱导机体产生良好的免疫应答和免疫保护作用[15,17-19]。进一步研究证明,引起奶牛乳腺炎的三种链球菌GapC 具有交叉免疫原性,且能够诱导良好的交叉免疫保护作用[20-21]。但引起奶牛乳腺炎的链球菌和葡萄球菌的GapC 之间是否存在交叉免疫原性尚缺乏研究。为此,研究对S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC 之间理化特性、同源性、免疫原性及抗原表位进行生物信息学分析,为进一步实验研究其诱导的交叉免疫原性和交叉免疫保护作用提供参考。
1 材料与方法
1.1 菌株
所用GapC 氨基酸序列均来自NCBI 上发布菌株的序列(表1)。
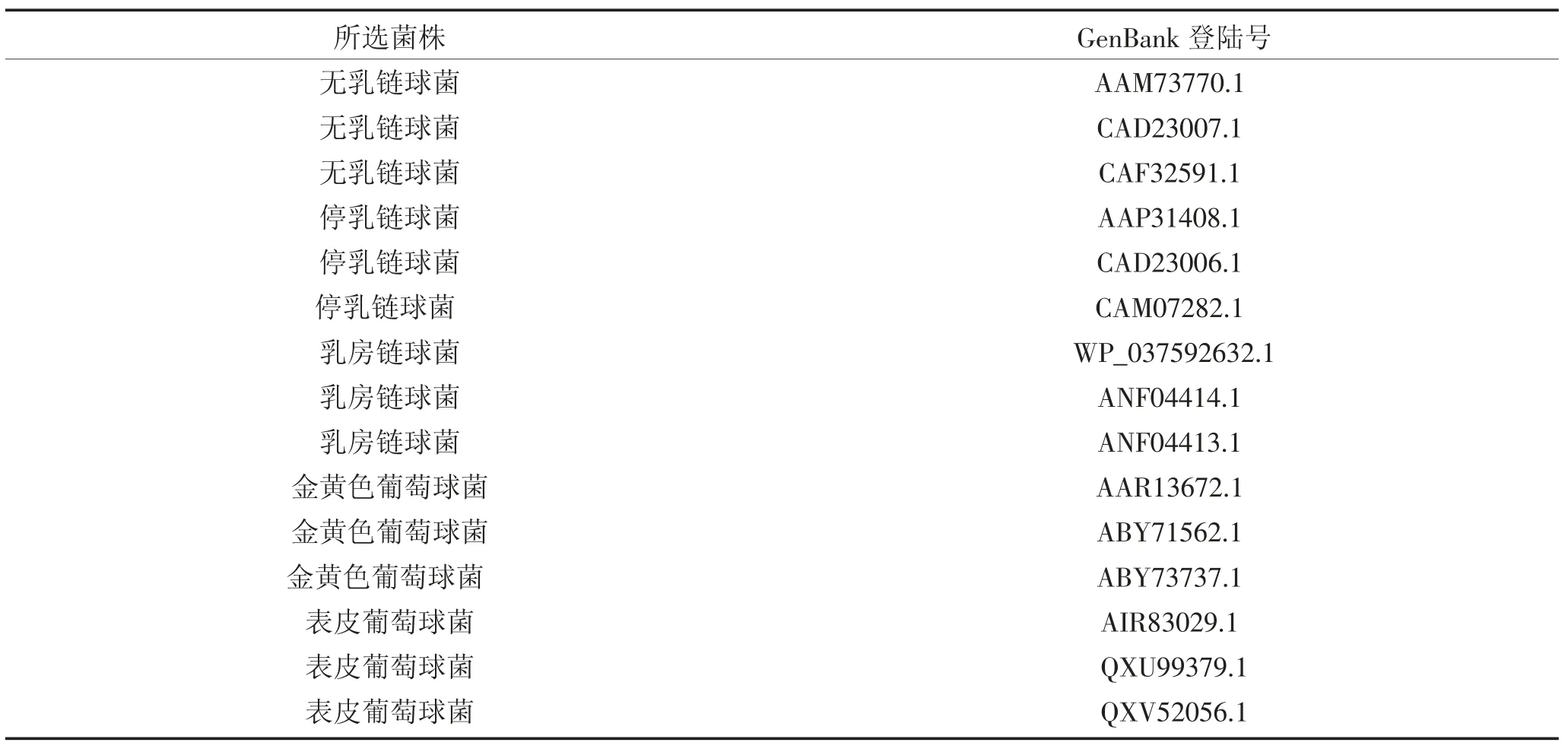
表1 GenBank 上选取相关GapC 氨基酸序列登录号Table 1 Login numbers of relevant GapC amino acid sequences selected from GenBank
1.2 方法
1.2.1 GapC 氨基酸同源性分析及序列比对
选用DNAstar 和NCBI 在线分析网站中的BLAST 功能对各菌株GapC 的氨基酸序列同源性进行分析;同时使用Geneious Prime 软件(网址:https://www.geneious.com/)中Pairwise/Multiple Align 功能对各菌株GapC 的氨基酸序列进行比对。
1.2.2 GapC 二级、三级结构预测
GapC 的二级结构通过在线软件Protein Structure Prediction Server(PSIPRED)数据库(http://bioinf.cs.ucl.ac.uk/psipred/)进行预测;采用Swiss-Model 数据库(http:/swissmodel.expasy.org/)同源性模型进行蛋白质的三级结构预测分析。
1.2.3 GapC 基本理化性质分析
使用ProtParam(http://web.expasy.org/protparam)分析各菌株GapC 的理化性质;使用蛋白质在线分析软件ProtScale(https://web.expasy.org/protscale/)分析不同菌株GapC 蛋白的亲水性。
1.2.4 GapC 的T 细胞和B 细胞线性表位预测分析
使用IEDB 抗原表位数据库(http://www.iedb.org/);其中线性B 细胞表位选择Bepipred Linear Epitope Prediction 2.0 预测方法,阈值设置为0.500,选取阈值大于0.500 的线性B 细胞表位,GapC 的CD4+T 细胞表位选择MHC Ⅱ类分子结合方式、CD8+T 细胞表位择MHC I 类分子结合方式,选取IC50 值小于50 nM的T 细胞表位。
2 结果与分析
2.1 GapC 的基本理化性质及抗原性分析结果
S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC 均由336 个氨基酸组成,相对分子质量为35 kDa 左右;等电点分别为5.22、5.34、5.25、4.89、4.83;不稳定系数分别为21.60、17.84、21.80、30.09、25.85,均属于理化性质稳定蛋白,S.dysgalactiae 的GapC 更加稳定;平均亲水性数值分别为-0.088、-0.092、-0.251、-0.125、-0.218、-0.203,均存在大量的亲水基团,属于亲水性蛋白;对抗原性预测分析结果显示,在GapC 中存在多处具有可以形成抗原表位的序列,结果详见图1。以上分析表明5 种病原菌GapC 具有良好的抗原性。
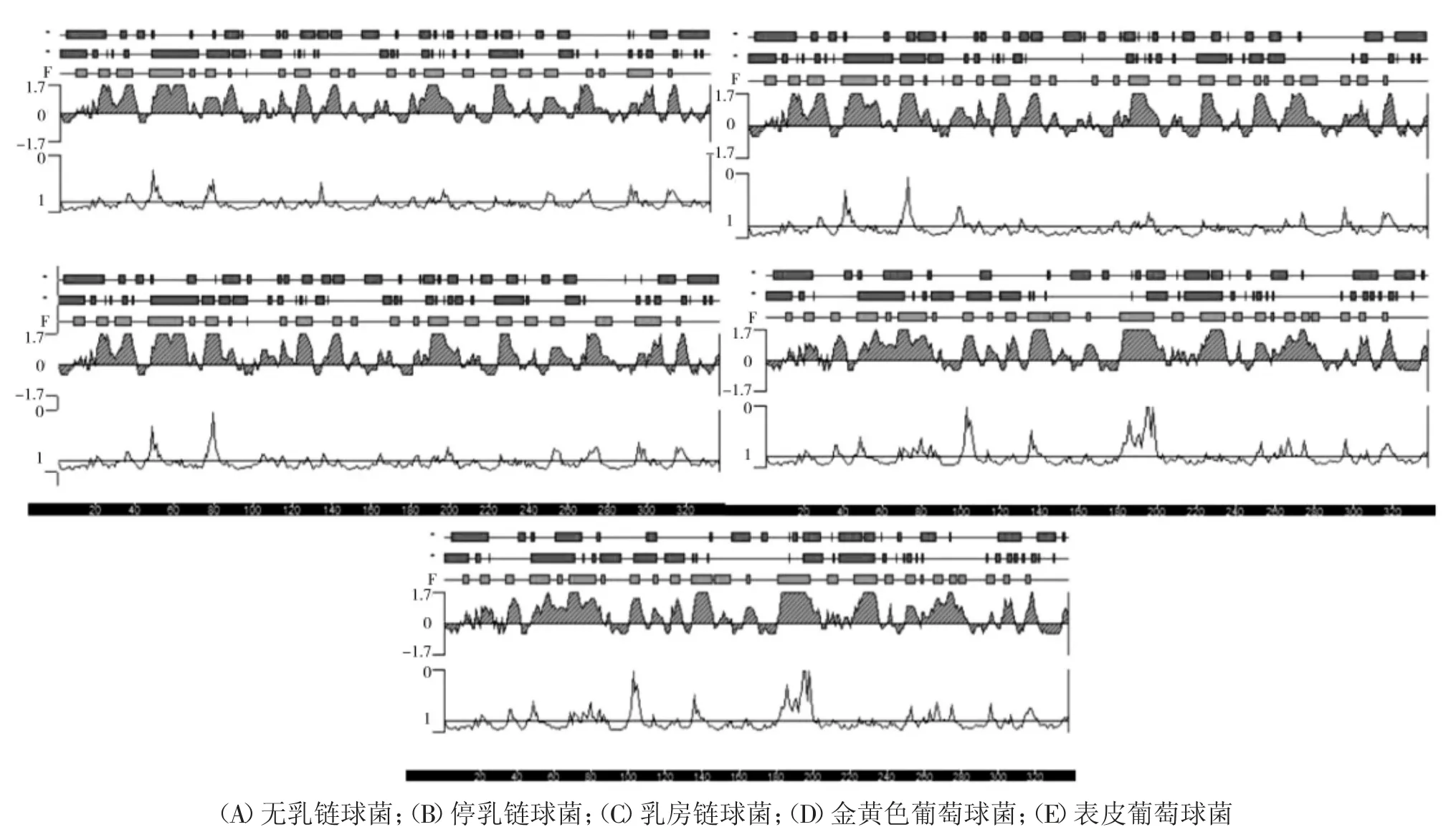
图1 GapC 蛋白抗原性预测结果Fig.1 Prediction of antigenicity of GapC protein
2.2 GapC 蛋白二级、三级结构分析结果
S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC 分别含有11、12、11、12、13 个α-螺旋区和16、18、15、17、18 个β-折叠区;α-螺旋分别占32.44%、27.68%、33.63%、30.06%、30.36%,β-折叠分别占22.62% 、25.89% 、19.94% 、22.92% 、22.92%;无规则卷曲分别占44.94%、46.43%、46.43%、47.02%、46.73%,通过对GapC 的二级结构分析发现各菌株中无规则卷曲占比较多,形成T 细胞或B 细胞表位可能性较大;分别对S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC进行三级结构预测,结果显示,与在线网站中的模板相比,其一致性分别为95.24%、99.70%、93.15%、100%、92.26%,可见五种的GapC 的一致性较高,各GapC 三级结构预测结果如图2 所示,说明,5 种细菌的GapC 的三级结构比较相似。
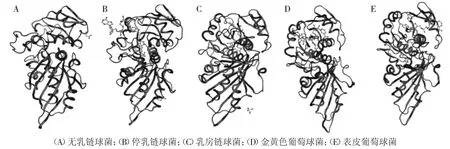
图2 GapC 蛋白三级结构预测Fig.2 Tertiary structure prediction of GapC protein
2.3 GapC 的氨基酸序列同源性分析结果
对S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC 的氨基酸序列进行同源性分析结果显示,氨基酸序列同源性为66.9%~100.0%,其中三种链球菌的GapC 氨基酸序列同源性可达到90.5%~94.0%,金黄色葡萄球菌与表皮葡萄球菌的GapC 氨基酸序列同源性达到92.0%~92.3%;而上述2 种葡萄球菌和3 种链球菌之间的GapC 氨基酸序列同源性67.8%~69.9%之间,结果见表2;并发现GapC 突变较多的位置为:70~81、100~110、163~172、186~194、247~258、300~306,结果见图3。
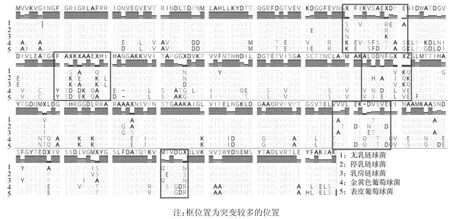
图3 GapC 蛋白氨基酸序列比对Fig.3 Amino acid sequence alignment of GapC protein
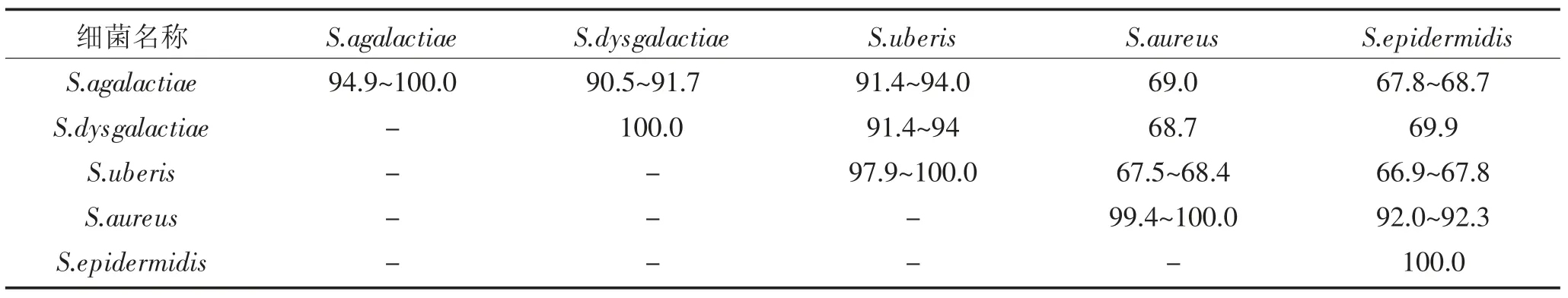
表2 GapC 氨基酸序列同源性分析(%)Table 2 Homology analysis of amino acid sequence of GapC from the pathogens(%)
2.4 GapC 的线性B 细胞表位和T 细胞表位预测分析结果
对S.agalactiae、S.dysgalactiae、S.uberis、S.aureus、S.epidermidis的GapC 的氨基酸序列进行线性B 细胞表位预测,结果显示存在共同B 细胞表位有4 个,详见表3。CD4+T 细胞表位预测结果显示,它存在共同CD4+T 细胞表位9 个,详见表4。CD8+T 细胞表位预测结果显示,共同CD8+T 细胞表位9 个,结果详见表5。其中,有4 个CD4+T 细胞表位与CD8+T 细胞表位氨基酸序列存在重叠。

表3 GapC 的线性B 细胞表位预测结果Table 3 Prediction results of linear B-cell epitopes on GapC
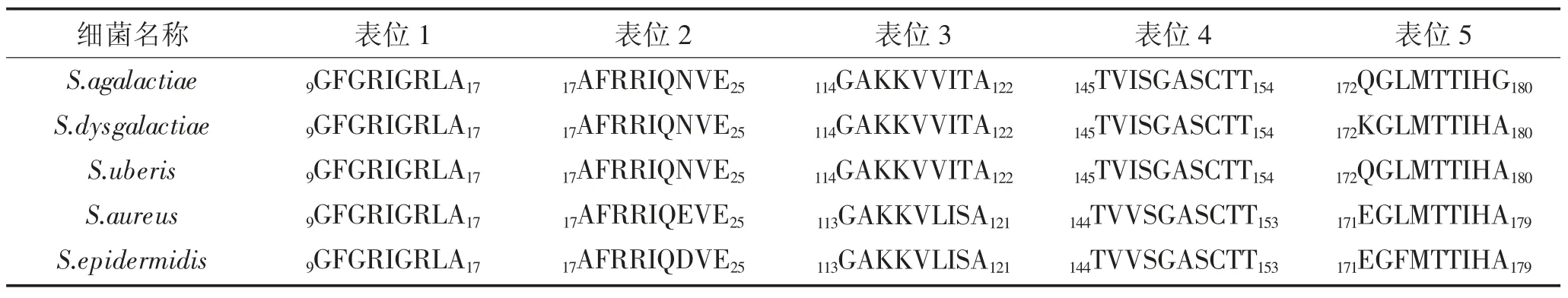
表4 GapC 的CD4+T 细胞表位核心序列预测结果Table 4 Prediction results of the core sequence of CD4+T cell epitope in GapC

表5 GapC 的CD8+T 细胞核心表位预测结果Table 5 Prediction results of the core sequence of CD8+T cell epitope in GapC
3 讨论
奶牛乳腺炎是严重危害奶牛健康和影响奶业发展的重要疾病之一。引起奶牛乳腺炎的病原菌种类繁多,但以链球菌(S.agalactiae,S.dysgalactiae,S.uberis)和葡萄球菌(S.aureus,S.epidermidis)感染为主。当病原菌侵入机体后,抗原提呈细胞对病原菌进行吞噬处理,通过MHC 将抗原决定簇即抗原表位呈递给CD4+T 细胞,进而使CD4+T 细胞活化、增值,分化成辅助性T(Th)细胞和记忆性T 细胞。Th 细胞调节B细胞和CD8+T 细胞活化,并能调控和增强巨噬细胞等功能活性,从而发挥Th 细胞对固有免疫和适应性免疫的调节作用。在这一过程中,真正发动抗感染免疫应答的有效抗原部分是抗原表位,即B 细胞表位、CD4+T 细胞表位和CD8+T 细胞表位[22-23]。不同抗原是否存在共同表位能够反映抗原间的交叉免疫原性。在金黄色葡萄球菌和链球菌抗感染免疫过程中,B 细胞介导的体液免疫应答和T 细胞特别是CD4+T细胞介导的细胞免疫应答发挥着重要作用[25]。因此,研究在对葡萄球菌和链球菌GapC 交叉免疫原性分析时,着重分析了这些抗原表位。如果葡萄球菌和链球菌GapC 之间存在共同抗原表位,就说明存在着交叉免疫原性和交叉免疫保护作用。
通过研究分析,上述5 种细菌的GapC 均由336个氨基酸组成,相对分子质量为35 kDa 左右,是具有亲水性的非经典分泌蛋白,且无规则卷曲占比较高,说明具有良好的抗原性。3 种链球菌GapC 非常保守,氨基酸序列同源性90.5%~94.0%,金黄色葡萄球菌和表皮葡萄球菌的GapC 也非常保守,氨基酸序列同源性92.0%~92.3%;而3 种链球菌与2 种葡萄球菌之间GapC 的氨基酸序列同源性67.8%~69.9%。5 种细菌共存在4 个共同线性B 细胞表位。3 种链球菌的4 个线性B 细胞表位高度保守,其中表位1 完全一致;表位3 的187 位V 突变为I,而I 与V 的性质十分相似、分子量相相差较小,可能不会影响表位的性质和功能;表位4 的271 位Y 变异为F,而Y 与F同属芳香族氨基酸,也是性质十分相似、分子量相差较小,可能对表位的性质和功能影响不大;表位2 的102 位S 突变为K、104 位A 突变为E,如果这两个位点的氨基酸是表位关键氨基酸,可能会影响表位之间的交叉免疫原性和交叉反应。此外,有研究证实这3 种链球菌间还存在链球菌特有的其他共同表位[26],即3 种链球菌GapC 之间肯定存在明显的体液交叉免疫原性和交叉反应性。2 种葡萄球菌的4 个表位也高度保守,表位1、表位3 氨基酸序列完全一致;表位2 的104 位D 突变为E,二者均为酸性氨基酸,性质相近,E 仅比D 多出一个-CH2-,可能不会影响表位的性质和功能;表位4 的268 位D(天冬氨酸)突变为N(天冬酰胺),而二者分子量相近,仅一个化学基团差别,可能二者的性质和功能差别也不大,所以2 种葡萄球菌GapC 之间肯定存在明显的体液交叉免疫原性和交叉反应性。在上述葡萄球菌和链球菌的4个表位之间,表位1 存在50 位T 与M 的突变,已有研究证明该表位T 是关键氨基酸,其突变就导致了不再出现交叉反应[26];而其他3 个表位都存在多个氨基酸变异,很可能会影响链球菌与葡萄球菌GapC 之间的体液交叉免疫原性和交叉反应性。但该判断有待于实验进一步证实。据报道[27],在抗原B 细胞表位中,构象性B 细胞表位具有更重要的价值,但研究未对构象B 细胞表位进行分析。
对CD4+T 细胞表位分析显示,5 种菌存在着9个共同CD4+T 细胞表位,而且表位都相当保守,菌株间的表位很少存在变异,即便存在变异如N 变异为E 或D、V 变异为L 或I、Q 变异为K 或E 等,它们突变氨基酸之间的分子量、性质等相近,对CD4+T细胞表位的识别和应答可能不会有明显影响,进而不会明显影响CD4+T 细胞交叉免疫原性,由此认为CD4+T 细胞表位的交叉免疫原性较强。而有关CD8+T细胞活化对奶牛乳腺炎链球菌和葡萄球菌的抗感染免疫作用未见明确报道,所以对CD8+T 细胞表位分析结果在此不进行深入讨论。
通过生物信息学分析表明,上述5 种病原菌的GapC 同源性较高,且免疫原性良好,3 种链球菌与2种葡萄球菌GapC 间存在一定的交叉免疫原性,为进一步研究GapC 作为潜在的预防链球菌、葡萄球菌感染的基因工程亚单位疫苗抗原提供参考。
猜你喜欢
遵义医科大学学报(2023年4期)2023-05-05 05:05:16
皮肤病与性病(2021年3期)2021-07-30 08:07:30
上海农业学报(2017年4期)2017-04-10 12:40:34
中国免疫学杂志(2017年1期)2017-01-17 04:53:25
特产研究(2016年3期)2016-04-12 07:16:15
中国医药生物技术(2015年4期)2015-12-26 08:26:36
华南农业大学学报(2015年5期)2015-12-04 03:04:38
畜牧兽医学报(2015年3期)2015-07-05 08:22:53
医学研究杂志(2015年6期)2015-07-01 17:41:11
微生物与感染(2015年1期)2015-02-28 17:42:38
