
BCL11A在神经细胞中稳定低表达对智能障碍风险基因的转录调控机制*
2023-06-27 05:56唐小涵陈金蓉苏炳银李淑蓉解学敏
成都医学院学报 2023年3期
唐小涵,陈金蓉,苏炳银,李淑蓉△,解学敏△
1.成都医学院 发育与再生四川省重点实验室组织胚胎学教研室(成都 610500);2.成都医学院 病理学与病理生理学教研室(成都 610500)
BCL11A基因座位于人类染色体2p16.1,编码krüppel样锌指蛋白[1]。BCL11A在血液和神经组织中呈高表达,可调节血红蛋白的转化并介导B细胞恶性肿瘤的发生[2]。BCL11A是皮质发育所必需的关键因子[3],在皮质发育过程中,BCL11A调控神经元从多极形态到双极形态的转换,从而参与深层皮质中的投射神经元形成[4-5]。BCL11A缺陷的神经元表现为辐射迁移受损和Semac3c的过表达,进而参与调节智能障碍(intellectual disability,ID)关联基因TBR1 在皮质中的表达,导致ID[6]。
ID的临床表现为进行性的认知能力受损,伴随适应社会环境能力的严重缺陷,其全球发病率高达2%~3%[7],病因多源于遗传因素。遗传性ID受蛋白表达、DNA碱基修饰、染色质共价和非共价变化等多种转录以及表观遗传调控[8]。临床报道[9]显示,BCL11A基因座的缺失会介导2p15-p16.1 缺失型ID发生,并伴有持续性胎儿血红蛋白升高。2p15-p16.1 微缺失综合征是一种特殊类型的遗传性ID,其临床表现为中至重度ID、自闭症、小头畸形以及包括皮质在内的大脑结构异常等特征[10]。然而,BCL11A在神经发育中的调控方式及参与2p15-p16.1 微缺失综合征的具体机制尚需进一步阐释。
前期研究[11]证实,BCL11A主要在小鼠胚胎13.5 d的大脑皮质表达,并通过设计siRNA瞬时敲降BCL11A,致使细胞转录活化水平整体下降,初步证明BCL11A在胚胎皮质发育过程中具有转录激活作用。为了进一步探索BCL11A如何参与神经组织的表观遗传调控,本课题组在此基础上,首先通过慢病毒载体构建能长期、稳定敲降BCL11A的细胞株,并利用实时荧光定 量PCR(reverse-transcription quantitative PCR,RT-qPCR)和蛋白质印迹技术,探索BCL11A对皮质发育关键转录因子的表达影响,继而转录组测序分析BCL11A敲降后差异基因的富集情况,最终通过染色质免疫共沉淀(chromatin immunoprecipitation,ChIP)验证差异基因启动子的染色质活性变化,从而初步揭示BCL11A在神经细胞中对ID风险基因的转录调控机制。
1 材料与方法
1.1 实验细胞
293T细胞系(人肾上皮细胞系)来源于人胚肾细胞、SH-SY5Y细胞系(人神经母细胞瘤细胞系)来源于人神经母细胞,均为本实验室冻存细胞。
1.2 试剂与仪器
1)主要试剂:蛋白质印迹技术整套试剂(上海碧云天)、蛋白酶抑制剂(北京擎科)、RIPA裂解液(上海碧云天)、anti-Ctip1 抗体(上海Abcam)、anti-β-actin抗体(上海Abcam)、anti-GAPDH抗体(上海Abcam)、anti-BCL11A 抗体(上海Abcam)、pPolⅡ(上海Abcam)、H3K9ac(上海Abcam);opti-MEM 培养基和达尔贝科极限必需培养液(Dulbecco minimum essential medium,DMEM)/F12(美国Gibco)、DMEM培养基(美国Gibco)、0.25%胰酶及胎牛血清(美国Gibco);抗生素-抗真菌(美国Thermo)、聚凝胺(北京Solarbio)、TRIzol(美国Thermo)、逆转录RT-qPCR试剂盒(北京Trans Gen Biotech)。2)主要仪器:Olympus SZX2-ILLT荧光显微镜(日本Olympus)、PCR核酸扩增仪(美国Bio-Rad)、CO2培养箱(美国Thermo)、倒置荧光显微镜(日本Nikon)、台式低温离心机(美国Thermo)、RT-qPCR仪及CFX96TMReal-time system(美国Bio-Rad)。
1.3 实验方法
1.3.1 细胞的培养与传代 细胞复苏后,在37 ℃、5% CO2条件下,使用完全培养基(10% Gibico胎牛血清+1%抗真菌抗生素)培养HEK293T、SH-SY5Y细胞,每1~2 d采用1×PBS轻柔清洗后,更换1 次培养基,细胞汇合度达80%左右时进行传代。
1.3.2 shRNA慢病毒质粒的构建及病毒收集 在NCBI网站上下载人源BCL11A基因的全长序列,设计5组慢病毒序列(表1)。PCR扩增目的片段后,引物退火连接目标片段,待感受态细胞为冰水混合状态时进行转化连接产物,然后在37 ℃培养箱中过夜培养。第2天挑选克隆于37 ℃培养箱中,220 r/min摇晃过夜培养后,提取质粒测序鉴定。

表1 慢病毒引物序列
1.3.3 慢病毒介导稳定敲降BCL11A的SHSY5Y细胞稳定株的构建 本研究部分设计并根据测序结果,筛选出测序结果正确的针对BCL11A的shRNA质粒: sh1a、sh2c、sh3d、sh4b。利用载体质粒pLKO.1-shRNA1a/2c/3d/4b以及HIV慢病毒包装质粒psPAX2、pMD2.G(质量比例为10∶7.5∶3.5 )进行共转染。以eGFP为对照,收集48、72、96 h的含有shRNA病毒上清,选择72 h病毒感染SH-SY5Y细胞。细胞生长密度为80%时进行感染,病毒感染48 h后,每隔48 h向完全培养基里加入嘌呤霉素10 g/L Polybrene筛选,1 周后筛选出目的细胞株。
1.3.4 RT-qPCR 通过TRIzol法提取各组细胞的总RNA,然后使用超微量分光光度计检测RNA 的浓度和纯度,按说明书取2 μg总RNA进行逆转录,参照试剂盒TransScript All-in-One First-Stand cDNA Synthesis SuperMix for RT-qPCR说明书步骤操作,得到cDNA,以cDNA为模版进行RT-qPCR检测目的基因表达情况。
1.3.5 蛋白质印迹技术验证BCL11A蛋白表达水平收集细胞,用PBS缓冲液清洗2~3 次,用RIPA裂解液冰上裂解60 min后,将裂解液经12000 r/min,离心半径9.6 cm,4 ℃,离心30 min。经浓度12%SDS-PAGE分离胶、5% SDS-PAGE浓缩胶、电泳电压60~120 V垂直电泳、300 mA恒流120 min转膜。PVDF膜用含5%脱脂奶粉的封闭液室温封闭2 h后,分别与BCL11A一抗(1∶1000)和GAPDH一抗(1∶1000)在4 ℃摇床孵育过夜。含聚山梨酯-20 的Tris盐酸缓冲液(Tris buffered saline with tween®20,TBST)清洗3 次后,再与辣根过氧化物酶标记的二抗(1∶5000)室温孵育2 h,洗涤3 次后,将化学发光试剂(electrochemiluminescence,ECL)A 液和B 液1∶1比例混合,覆盖到膜上,置于 Bio-Rad 成像系统进行拍摄,使用ImageJ软件进行分析。
1.3.6 ChIP实验分析神经元特异性因子的启动子活性SH-SY5Y-sh细胞用终浓度1%甲醛固定,加入甘氨酸终止反应,PBS清洗后,离心收集固定好的细胞,Lysis buffer重悬细胞放冰上10 min,离心后用PBS漂洗。NCP bufferⅠ重悬,冰上放置10 min,ChIP Lysis buffer重悬,冰上放30 min 裂解细胞核膜,超声仪使染色质碎裂,用等超声体积的酚氯仿提取超声样品,取上清液,超声样品分为3 组,即抗体IP组、IgG组和input组。每个样品加入IgG,再加入磁珠Invitrogen dynabeads protein A磁珠,4 ℃摇床慢速摇2.5 h,去除非特异性结合的杂质,磁架上静置1 min。将管中清亮液体吸取到新EP管,加入dynabeads protein A,4 ℃摇床过夜。依次使用Wash bufferⅠ、Ⅱ、Ⅲ分别4 ℃摇床慢速清洗磁珠,弃去液体,向珠子中加入新鲜制备的提取缓冲液,置于磁架上提取染色质复合物。样本提取液和input样品中,加入RNase、NaCl、蛋白酶K,混匀,65 ℃水浴裂解过夜,第2天抽提DNA后,RT-qPCR定量分析。
1.4 统计学方法
采用ImageJ及Graphpad Prism 8.0 软件进行统计学分析,定量资料采用()描述,两组间比较采用t检验,3 组及以上表达水平差异采用ANOVA分析。检验水准α除特别说明外均设定为0.05。
2 结果
2.1 成功构建稳定敲降BCL11A 的SH-SY5Y 细胞株
成功构建4 株稳定敲降BCL11A的SH-SY5Y-sh细胞株,不同慢病毒细胞株的细胞生长速率存在明显差异,SH-SY5Y-sh1a、SH-SY5Y-4b的生长密度较SH-SY5Y-2c、SH-SY5Y3d低(图1)。

图1 嘌呤霉素筛选后感染成功的SH-SY5Y细胞
2.2 SH-SY5Y-sh 细胞株中BCL11A 的mRNA 表达水平
SH -SY5Y- sh细胞株的RT-qPCR 实验表明,SH-SY5Y-sh1a、SH-SY5Y-sh2c、SH-SY5Y-sh3d细胞株中BCL11A的mRNA干扰效果明显,其中,SH-SY5Y-sh1a细胞株BCL11A的mRNA水平下调为对照组的73.5%(P<0.01),SH-SY5Y-sh2c的mRNA水平下调为43.0 %(P<0.001),SH-SY5Y-sh3d细胞株BCL11A的mRNA水平下调为39.0%(P<0.001)(图2)。

图2 RT-qPCR检测SH-SY5Y-sh细胞中BCL11A的干扰效率(,n=3)
2.3 SH-SY5Y-sh 细胞株中BCL11A 蛋白表达水平改变
通过蛋白质印迹技术检测BCL11A蛋白的表达情况,可见在构建成功的4 组慢病毒细胞株中,SH-SY5Y-Sh1a、SH-SY5Y-Sh2c、SH-SY5Y-Sh4b细 胞中BCL11A蛋白表达量下降>80.0%(P<0.001),而SH-SY5Y-Sh3d细胞BCL11A蛋白表达量下调约20.0%(P<0.01);将慢病毒细胞株的蛋白表达量灰度值统计分析显示,SH-SY5Y-Sh1a、SH-SY5Y-Sh2c 细胞的BCL11A蛋白表达量最低(图3)。

图3 SH-SY5Y-sh细胞中BCL11A蛋白表达水平(,n=3)
2.4 BCL11A 敲降后调控皮质神经元发育关键基因TCF12、MEF2D 的表达
TCF12 参与调控RNA聚合酶Ⅱ启动子、免疫应答、肌发育、调控转录等重要生物学过程[12];MEF2 是转录调节因子MADS-box家族成员之一,在脊椎动物中参与转录、调控RNA 聚合酶Ⅱ启动子、肌发育等过程[13]。
RT-qPCR实验显示,BCL11A敲降后,SH-SY5Y-sh1a细胞株中皮质发育关键转录因子TCF12的mRNA表达上调至1.25倍(P<0.001)、MEF2D的mRNA表达量上调至3倍(P<0.001)。SH-SY5Y-sh2c细胞株中皮质发育关键转录因子TCF12的mRNA表达上调至1300倍(P<0.001),MEF2D的mRNA 表达量上调至3.8倍(P<0.001)。由此提示,BCL11A对于皮质神经元关键转录因子的表达具有负向调控作用,BCL11A对TCF12、MEF2D的抑制作用呈线性负相关(图4)。
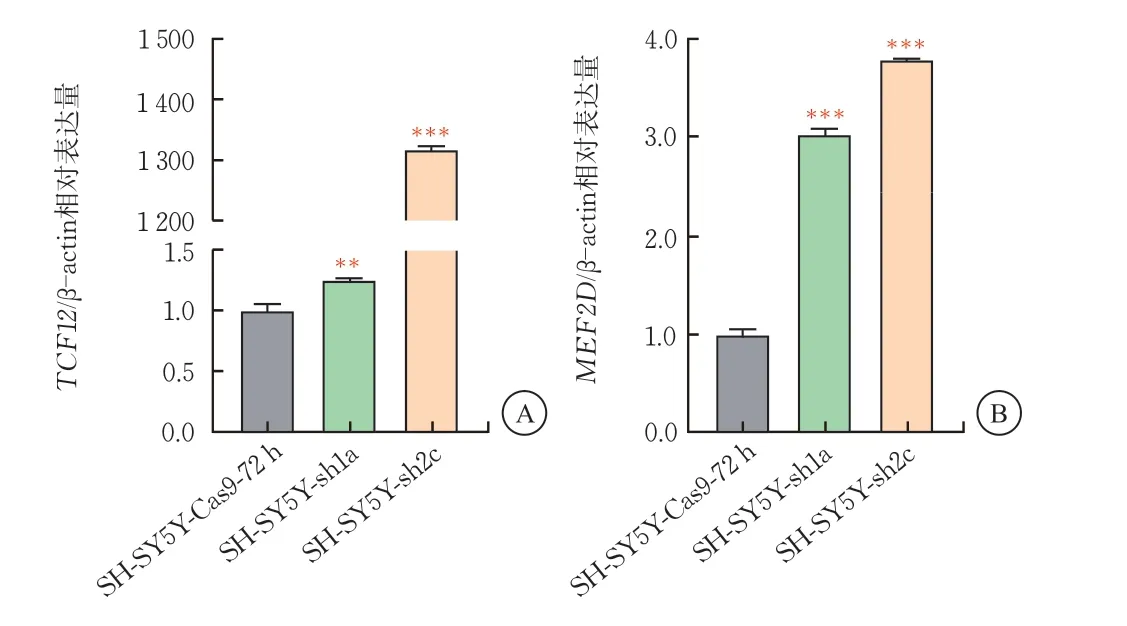
图4 SH-SY5Y-sh细胞中皮质发育关键转录因子TCF12、MEF2D的mRNA表达水平(,n=3)
2.5 BCL11A 敲降后关键基因BCL11B、CTCF、ADCYAP1R1、NeuroD 1/2、PAX6、SATB 1/2 的表达水平
选取BCL11A蛋白水平敲降效果明显的SH-SY5Y-sh1a、SH-SY5Y-sh2c细胞株进行RT-qPCR实验,观察皮质神经元发育关键转录因子的表达变化。转录因子BCL11B为参与神经系统及免疫系统发育的核内转录因子[14];CTCF广泛存在于真核生物中,可阻断增强子和启动子的相互作用[15];ADCYAP1R1 在焦虑、恐惧认知和应激反应中发挥重要作用[16];NeuroD1/2 参与调控RNA聚合酶Ⅱ启动子,在神经系统发育以及细胞形态分化中起重要作用[17];在胚胎发育过程中,PAX6 对调节神经系统起重要作用[18];SATB1/2 参与调节神经系统的分化和成熟[19]。
RT-qPCR 结果显示,构建的SH-SY5Y-sh1a、SH-SY5Y-sh2c细胞株中,BCL11A的mRNA水平降至44.0%~70.0%(P<0.001),此时细胞株内皮质神经元发育关键基因BCL11B、CTCF、NeuroD1、NeuroD2、SATB1、ADCYAP1R1、PAX6、SATB2 的mRNA表 达降低。以上结果提示,BCL11A可正向调控以上皮质神经元关键转录因子的表达,且对胚胎发育关键转录因子ADCYAP1R1、PAX6、SATB2 的降低程度更明显(图5)。
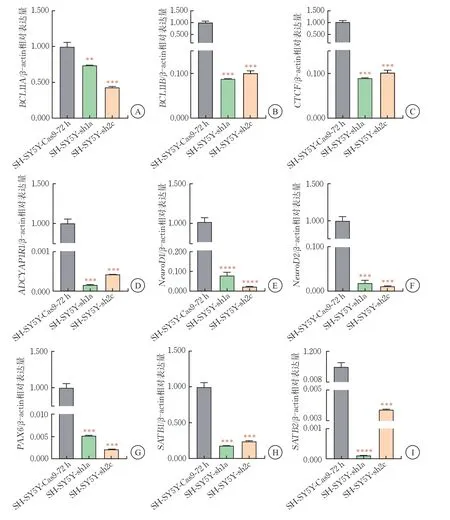
图5 SH-SY5Y-sh细胞中皮质发育关键转录因子BCL11A、BCL11B、CTCF、ADCYAP1R1、NeuroD1/2、PAX6、SATB1/2的mRNA表达水平(,n=3)
2.6 转录组测序分析BCL11A 在神经细胞中参与调控的靶基因及相关启动子染色质水平变化
转录组测序样显示,SH-SY5Y-sh细胞株组差异表达的246个基因进行基因本体论(gene ontology,GO)富集分析显示,193个基因富集到人类疾病相关因素的GO条目(P<0.05),有约200个基因富集到生物系统及环境信息处理、细胞过程、代谢、遗传信息处理等方面皆有参与(图6A~B)。通过京都基因和基因组百科全书数据库(kyoto encyclopedia of genes and genomes,KEGG)对BCL11A敲降后差异基因参与的基因组、化学和系统功能信息进行分析,结果显示,BCL11A代谢通路分为多个分支,其中在组织系统异常富集数量排名第3 的为神经系统异常(图6C)。在生物学功能中,BCL11A与免疫系统、内分泌系统以及神经系统高度相关,KEGG疾病富集柱状图分析结果显示,BCL11A在神经系统疾病有高度富集(图6D)。

图6 BCL11A参与的重要生物学过程
RNA测序结果显示,SH-SY5Y-sh细胞株组差异表达基因共824 个差异有统计学意义(P<0.05),其中有649 个基因上调,175 个基因下调;与神经发育异常相关的差异表达基因有两个,分别是下调基因GRM7 与上调基因GRIN1,都是ID风险基因(图7)。GRM7 是一种抑制性异源三聚体G蛋白偶联受体,它通过河马信号通路影响脑发育,可调节哺乳动物中枢神经系统突触前末端的神经递质释放和突触可塑性,为神经发育障碍风险基因[20]。GRIN1 基因突变可影响NMDA受体的生理功能,引起神经元功能异常,最终导致大脑到肠道的各种神经、精神症状[21]。
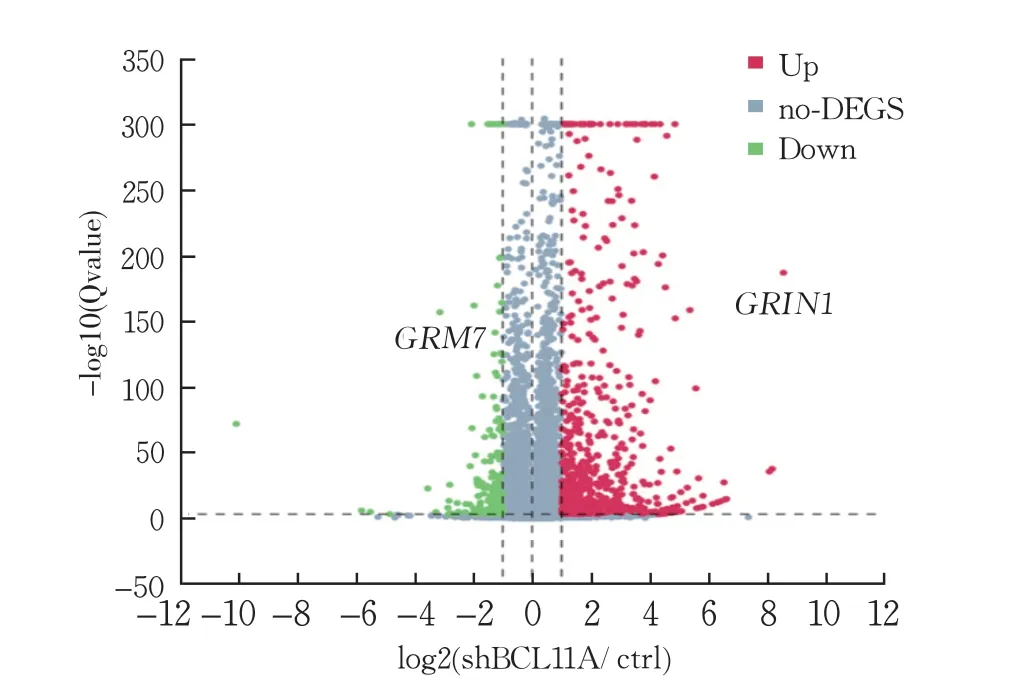
图7 SH-SY5Y细胞差异基因表达水平的分布情况
2.7 BCL11A 敲降后神经元中GRM7 启动子染色质富集情况
为探索神经细胞中BCL11A的上游差异表达基因GRM7的启动子染色质活性,课题组针对GRM7的启动子进行了ChIP-qPCR实验,结果显示,在SH-SY5Y-sh细胞中,ID基因GRM7 启动子位点的转录活性标志pPolⅡ(抗Phospho-Rpb1 CTD)以及染色质活性修饰H3K9ac(抗H3K9ac)的富集都明显降低;其中,mRNA水平和蛋白表达水平下调效果明显的SH-SY5Y-sh1a、SH-SY5Y-sh2c细胞中pPolⅡ的富集度下调95%以上(P<0.001),较IgG下调92%以上(P<0.001)。GRM7启动子染色质活性修饰H3K9ac的富集度降低50%以上(P<0.001),SH-SY5Y-sh1a细胞中H3K9ac的富集较对照组降低约25%(P<0.01),SH-SY5Y-sh2c细胞中H3K9ac的富集较对照组降低约90%(P<0.001)。以上结果表明,GRM7的启动子染色质活性及染色质修饰活性在BCL11A敲降后明显降低,且在SH-SY5Y-sh2c细胞中富集度降低明显,提示神经发育异常相关基因GRM7 可能受到BCL11A的正向调控(图8)。

图8 SH-SY5Y-sh1a/2c/3d/4b细胞中GRM7 启动子染色质富集情况(,n=3)
2.8 BCL11A 敲降后神经元中GRIN1 启动子染色质富集情况
RNA测序结果显示,ID风险基因GRIN1 表达上调。课题组针对GRIN1 的启动子进行了ChIP-qPCR实验,以探索神经细胞BCL11A敲降后GRIN1 的启动子染色质活性。SH-SY5Y-sh2c、SH-SY5Y-sh3d细胞中GRIN1 启动子处pPolⅡ的富集度上调为140 倍,SH-SY5Y-sh4b的富集度上调为243 倍。GRIN1 启动子处SH-SY5Y-sh1a细胞的pPolⅡ富集度为对照组的4.5倍,SH-SY5Y-sh2c细胞的pPolⅡ富集度为对照组的749倍,SH-SY5Y-sh3d细胞的pPolⅡ富集度为IgG的7.7 倍,SH-SY5Y-sh4b细胞的pPolⅡ富集度为IgG的2800 倍。
SH-SY5Y-sh2c细胞中GRIN1 启动子染色质活性修饰H3K9ac的富集度上调为4倍,SH-SY5Y-sh3d的富集度上调为1.3倍。GRIN1启动子处SH-SY5Y-sh1a细胞的H3K9ac富集度为对照组的11 倍,SH-SY5Y-sh2c细胞的H3K9ac富集度为对照组的21倍,SH-SY5Y-sh3d细胞的H3K9ac集度为对照组的7.8倍,SH-SY5Y-sh4b细胞的H3K9ac富集度为对照组的1.4 倍(图9)。

图9 SH-SY5Y-sh1a/2c/3d/4b细胞中GRIN1 启动子染色质富集情况(,n=3)
以上实验结果表明,转录活性标志pPolⅡ以及染色质活性修饰H3K9ac在GRIN1启动子的富集呈现升高趋势,且在SH-SY5Y-sh2c细胞中的富集差异有统计学意义(P<0.05),提示ID风险基因GRIN1可能受到BCL11A的负向调控。
3 讨论
BCL11A在脑组织中高表达,它参与调节大脑中的轴突分支[22],其基因座缺失可致2p15-p16.1 微缺失综合征发生。前期研究[11]通过设计siRNA瞬时敲降BCL11A,初步证明BCL11A在神经元发育过程中具有转录激活作用。在此基础上,课题组构建了可稳定敲降BCL11A的SH-SY5Y-sh细胞株作为工具来进一步探索BCL11A对神经系统的影响机制。本研究发现,BCL11A正向调控部分胚胎发育关键转录因子,其中BCL11A对焦虑症状、恐惧应激因子ADCYAP1R1、神经系统发育障碍因子PAX6 的降低程度更明显。同时,在神经系统中,BCL11A负向调控神经元转录关键因子TCF12、MEF2D,且抑制作用呈线性负相关,由此提示,BCL11A除在胚胎皮质发育时期发挥重要作用外,还与ID及相关自闭症谱系障碍疾病的发生有着紧密联系。
GRM7 调节整个中枢神经系统的神经传递和突触可塑性,其与抑制性G蛋白偶联主要定位在突触前活性区[22-23]。在发育退化的雷诺氏综合征人类模型[24]中显示,患者运动皮层的mGlu7 蛋白表达水平降低。GRM7 在神经元发育早期突变可导致轴突生长严重受损,进而引起成熟神经元突触前末端数量缺陷[25]。在早期发育期间,使用mGlu7 激动剂可恢复诱导的轴突生长和突触前末端发育缺陷[26],表明GRM7 在神经发育障碍发病机制中的稳定表达具有关键作用。GRIN1相关神经发育障碍(GRIN1-NDD)的所有受影响个体表现为轻度至重度发育迟缓及ID,其皮质发育畸形个体的杂合致病变异位于结构域S2 和M3 中,且可能存在基因型-表型相关性[27]。NMDAR是人类大脑学习与记忆的分子开关,在人类大脑皮质中广泛分布,参与大脑发育及神经元活动[28]。GRIN1 编码GluN1 亚基,交替剪接产生8 种亚型,GRIN1 通过影响NMDAR通道,在大脑发育、学习、记忆和其他高级认知功能中起着重要作用。
本研究构建的SH-SY5Y-sh细胞株转录组测序分析发现,BCL11A的缺失与人类神经系统疾病高度相关,BCL11A缺失显示下游有两个神经发育迟滞风险差异基因GRM7 与GRIN1,且GRM7 可能受到BCL11A的正向调控,GRIN1 可能受到BCL11A的负向调控,通过ChIP验证其增强子活性同样受BCL11A的表达影响。由此可以推论,BCL11A和其他神经元关键因子共同作用于皮质发育的重要环节,并与ID密切联系,但相关的分子机制还有待进一步实验证实。
综上所述,BCL11A在神经元中功能重要且复杂,它参与多种神经系统发育关键因子的表达调控。BCL11A对胚胎神经元发育的功能及作为转录因子参与皮质发育的具体分子机制仍有待进一步研究,完善靶向转录因子BCL11A在皮质发育中的组织特异性,对研究和诊治2p15-p16.1 微缺失综合征至关重要。
猜你喜欢
畜牧兽医学报(2022年3期)2022-03-30
中国畜牧兽医(2022年1期)2022-02-15
成都医学院学报(2021年2期)2021-07-19
小资CHIC!ELEGANCE(2021年46期)2021-01-11
睿士(2020年11期)2020-11-16
现代泌尿外科杂志(2019年10期)2019-10-31
生物学通报(2019年2期)2019-06-15
医学研究杂志(2015年11期)2015-06-10
中国当代医药(2015年16期)2015-03-01
中国医药导报(2015年27期)2015-02-28
