
温度对Mhc>PINK1-RNAi转基因帕金森果蝇模型的影响
2023-06-19 02:22郝生慧贾若凡杜冠华张建琴
山西医科大学学报 2023年5期
郝生慧,贾若凡,尹 菲,杜冠华,张建琴*
(1山西大学中医药现代研究中心,太原 030001;2山西大学化学化工学院;3中国医学科学院北京协和医学院药物研究所;*通讯作者,E-mail:jiangqinzh3@sxu.edu.cn;#共同通讯作者,E-mail:dugh@imm.ac.cn)
帕金森病(Parkinson’s disease, PD)是常见的第二大神经退行性疾病,临床表现为静止性震颤、运动迟缓、肌强直和姿势步态异常等运动障碍,还表现为睡眠质量障碍、嗅觉障碍等非运动障碍[1]。其病理特征主要是黑质中多巴胺能神经元的丢失以及路易小体的形成[2]。目前已提出多种致病因素,包括环境、遗传、衰老等[3-5]。
1893年,Gowers首次提出PD患者有体温调节障碍,在寒冷时患者可能会加剧颤抖[6]。2005年,Meigal等[7]提出体温过低可引起PD患者运动障碍,适当的热条件(热中性或适度加温)可以改善PD患者的身体机能。在一般冷却条件(10 ℃,30 min)下,健康受试者可进行85~100次肘关节屈曲,而PD患者仅能进行36~38次肘关节屈曲[8],该研究表明一般冷却条件下PD患者的肌肉丧失了运动能力。另有研究发现适度加热可提高接受药物治疗的PD患者的肌肉运动能力[9]。升高温度可能会使帕金森病患者已经受损的大脑能量机制超负荷运转[10]。以上研究表明低温会加重PD患者的运动能力下降,适当的加热可以缓解PD患者的运动失调。
目前研究发现与帕金森疾病相关的致病基因有十几种,而应用最多、研究最广的有α-突触核蛋白、PINK1、Parkin、LRKK2、DJ-1等[11]。利用这几种基因构建的模型为PD发病机制的研究提供了较为优良的模型。PINK1突变是常染色体隐性遗传PD的第二大常见病因[12]。黑腹果蝇(Drosophilamelanogaster)是研究神经退行性疾病分子机制的一种有价值的昆虫模型。PINK1-RNAi果蝇模型导致能量消耗,寿命缩短以及选择性间接飞行肌肉和多巴胺能神经元的变性[13],显示出PD的关键特征。该模型常被用于探讨PD的发病机制或药物筛选。
在模式动物果蝇中,UAS-GAL4系统是常用的转基因表达工具。上游激活序列(upstream active sequence,UAS),是酵母中一种类似高等真核生物增强子的序列。GAL4(galactose 4)作为酵母转录激活因子,可结合UAS的特异性位点,并激活其下游基因的表达,从而实现对靶基因表达的操控[14]。在低温状态下,GAL4亦可驱动UAS靶基因表达[15]。果蝇的UAS-GAL4 RNA干扰系统(RNA interference,RNAi)可以实现特定时间特定组织靶标dsRNA的表达以沉默相应基因的表达。利用UAS-GAL4 RNAi系统构建的RNAi果蝇,在较低温度状态下,RNAi效率受到抑制,而在较高温度状态下,RNAi效率提高,且会随着温度的升高而逐渐增强[16,17]。已有研究表明适当的热条件有利于PD患者症状的缓解,但是适当的热条件对UAS-GAL4 RNAi转基因PD果蝇模型的影响还未见报道。为阐明上述问题,本研究以Mhc>PINK1-RNAi转基因PD果蝇模型为研究对象,将w1118作为对照组,通过25 ℃正常培养条件和29 ℃较高温度培养条件,探究不同温度对Mhc>PINK1-RNAi转基因PD果蝇模型中靶基因PINK1表达的影响,并结合行为学和生化指标来评价该模型。本研究结果将揭示温度对RNAi转基因PD果蝇模型的影响,为基于该模型的帕金森疾病的病理机制、药效、药理学研究提供科学依据。
1 材料和方法
1.1 果蝇的杂交
3个果蝇品系:UAS-PINK1-RNAi(BS31262;y[1]v[1];P{y[+t7.7]v[+t1.8]=TRiP.JF01203}attP);Mhc-GAL4(BS55133;w[*];P{w[+mC]=Mhc-GAL4.K}2/TM3,Sb[1])购于Bloomington Stock Center,黑腹果蝇w1118由山西大学应用生物学研究所提供,25 ℃室温条件下饲养。构建以下杂交组合:Mhc-GAL4品系处女蝇与UAS-PINK1-RNAi雄果蝇杂交,获得子一代Mhc>PINK1-RNAi果蝇。将对照组w1118雄果蝇和实验组Mhc>PINK1-RNAi雄果蝇,分别放置在25 ℃和29 ℃人工气候箱中培养,均在第10,20天收集后用于以下实验。
1.2 RT-qPCR技术检测PINK1基因表达量
PINK1基因表达量下降是Mhc>PINK1-RNAi转基因PD果蝇模型造模成功的首要检测指标。收集10 d和20 d的w1118雄果蝇和Mhc>PINK1-RNAi雄果蝇,每组随机挑选10只果蝇,用Trizol试剂提取总RNA并反转录为cDNA,以ribosomal protein 49(RP49)为内参基因,PINK1为目的基因,采用RT-qPCR技术检测靶基因的mRNA表达水平。各基因引物序列,RP49 F:5′-GACAGTATCTGATGCCCAACA-3′;R:5′-CTTCTTGGAGGAGACGCCGT-3′;PINK1 F:5′-AATCCCAACCCGTCCAAGC-3′;R:5′-AAACACTGCGACCCACCTCC-3′。上述试验重复3次,统计学分析实验结果。
1.3 爬行实验
收集10 d和20 d的w1118雄果蝇和Mhc>PINK1-RNAi雄果蝇,用CO2气体麻醉后,从每组中随机选择20只果蝇,并将它们放在垂直的塑料柱中(长度25 cm,直径1.6 cm)。在室温下放置20 min以适应环境后,将果蝇移至柱底部。记录在10 s内可以爬升或超过6 cm的果蝇数量,并以占果蝇总数的百分比表示[18]。上述实验独立重复5次,并进行统计学分析。
1.4 体视显微镜进行表型观察
翅膀异常是PINK1-RNAi转基因PD果蝇的一种典型的疾病表型。收集10 d和20 d的w1118雄果蝇和Mhc>PINK1-RNAi雄果蝇,各组随机挑选果蝇100只,CO2气体麻醉后,每10只果蝇置于1个果蝇培养管。待果蝇完全麻醉后(约60 min),在体式显微镜下观察果蝇翅膀形态,如果蝇翅膀重叠完好、与果蝇身体水平,则为正常形态;若果蝇翅膀分叉、下垂或竖立,则为异常形态[19],记录其异翅果蝇数,异翅率=异翅果蝇数/果蝇总数×100%。在相同条件下,重复上述试验5次,统计学分析实验结果。
1.5 酶联免疫技术测定多巴胺含量
收集10 d和20 d的w1118雄果蝇和Mhc>PINK1-RNAi雄果蝇,各组随机挑选20只果蝇,加180 μl生理盐水,研磨,按照小鼠DA酶联ELISA试剂盒(Bioswamp)说明书开展实验,形成抗体-抗原-酶标抗体复合物,洗涤显示转化为终止液,得到多巴胺组织液,使用多功能酶标仪进行检测,上述实验重复3次,并进行统计学分析。
1.6 数据处理
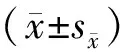
2 结果
2.1 不同温度对Mhc>PINK1-RNAi转基因PD果蝇PINK1基因表达水平的影响
结果显示,在常温25 ℃培养条件下,与w1118相比,转基因PD果蝇PINK1基因的表达在10 d和20 d分别下降了50.7%和84.6%,差异有统计学意义(P<0.01)。在29 ℃培养条件下,与w1118相比,转基因PD果蝇PINK1基因的表达在10 d和20 d分别上升了29.1%和12.9%(P<0.01,见图1)。以上结果表明,在25 ℃培养条件下,Mhc>PINK1-RNAi转基因PD果蝇体内可有效沉默PINK1基因的表达。在29 ℃培养条件下,PINK1基因没有被沉默。
A.25 ℃培养条件下果蝇中PINK1基因相对表达B.29 ℃培养条件下果蝇中PINK1基因相对表达
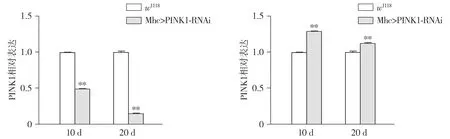
2.2 不同温度对Mhc>PINK1-RNAi转基因PD果蝇运动能力的影响
在25 ℃培养条件下,与w1118相比,10 d和20 d Mhc>PINK1-RNAi转基因PD果蝇爬行指数分别降低了14%和19%(P<0.01,见图2A),运动能力显著降低。在29 ℃培养条件下,与w1118相比,Mhc>PINK1-RNAi转基因PD果蝇爬行指数分别降低24%和31%(P<0.01,见图2B),运动能力显著降低。与25 ℃培养条件相比,在29 ℃培养条件下,Mhc>PINK1-RNAi转基因果蝇运动能力较低。
A.25 ℃培养条件下果蝇的爬行实验结果B.29 ℃培养条件下果蝇的爬行实验结果
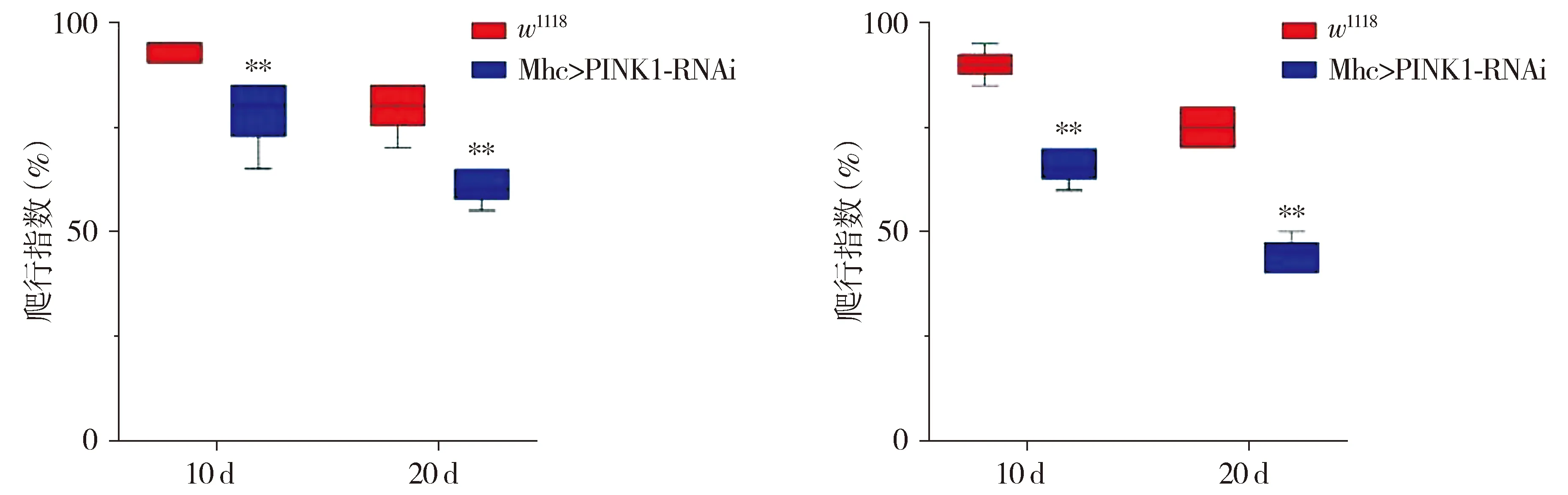
2.3 不同温度对Mhc>PINK1-RNAi转基因PD果蝇表型的影响
与总是保持翅膀与体轴平行的对照果蝇相比,异常翅膀表现出向上或向下的位置(见图3A)。25 ℃培养条件下,10 d和20 d Mhc>PINK1-RNAi转基因PD果蝇异翅率分别为37.7%和67.7%(P<0.01,见图3B)。29 ℃培养条件下,10 d和20 d Mhc>PINK1-RNAi转基因PD果蝇异翅率分别为4.6%和8.2%(P<0.05,见图3C)。与25 ℃培养条件相比,29 ℃培养条件下,Mhc>PINK1-RNAi转基因果蝇异翅率较低。
B.25 ℃培养条件下雄果蝇异翅率C.29 ℃培养条件下雄果蝇异翅率
2.4 不同温度对Mhc>PINK1-RNAi转基因PD果蝇多巴胺含量的影响
25 ℃培养条件下,与w1118比,10 d和20 d Mhc>PINK1-RNAi转基因PD果蝇体内多巴胺含量均显著降低(P<0.01,见图4A),而29 ℃培养下,10 d和20 d Mhc>PINK1-RNAi转基因PD果蝇与w1118果蝇体内多巴胺含量表达量均相当,无统计学意义(P>0.05,见图4B)。
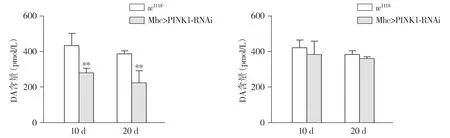
A.25 ℃培养条件下多巴胺含量 B.29 ℃培养条件下多巴胺含量
3 讨论
UAS-GAL4系统作为研究基因功能的强大工具,可以在特定部位控制靶基因的表达。因此,通过干扰果蝇体内与人类PD致病基因同源基因的表达可构建PD果蝇模型。已有研究表明RNAi效率对温度较敏感[16,17]。Fortier等[17]研究表明,利用GAL4依赖性启动子驱动Transformer-2(性别分化基因)基因的表达时,在22 ℃时果蝇发育为可育雌性,在29 ℃时果蝇发育为性中间体(雌雄同体),只有在29 ℃时果蝇出现功能丧失突变表型。本研究发现,Mhc-GAL4品系和UAS-PINK1-RNAi品系杂交的子一代RNAi果蝇在29 ℃培养条件下,PINK1基因没有沉默,而在常温25 ℃培养时,PINK1基因得到有效沉默。导致这一结果可能是在较高温度(29 ℃)培养条件下,转基因PD果蝇体内PD疾病与RNAi同时存在拮抗作用,温度对PD疾病的回调作用显著。
PD的流行病学研究表明,与夏季相比,冬季和春季出生的新生儿后期患PD的概率会增加[20]。人大脑是身体对热最敏感的器官[21],且PD患者表现出低于对照组的脑室内温度[22]。在大鼠模型中,温度对致密部黑质多巴胺能神经元有直接影响,致密部黑质多巴胺能神经元升温导致放电频率增加、输入电阻降低、内向电流和(Ca2+)增加[23]。PINKl突变引起多巴胺代谢异常或者多巴胺神经元功能异常从而导致PD。贾焕珍等[24]将MN9D细胞进行免疫荧光双标记,发现PINKl和酪氨酸羟化酶(TH)有共定位,表明二者之间可能存在相互作用。因此,在较高温度状态下(29 ℃),果蝇体内PINK1和多巴胺神经元可能存在相互作用,进而导致多巴胺含量的升高和PINK1基因干扰效率的下降。本研究发现29 ℃培养条件对PD果蝇症状有一定的缓解作用,即温度较高时,与w1118相比,Mhc>PINK1-RNAi转基因PD果蝇靶标基因PINK1未被沉默,异翅率降低,多巴胺含量回调且两组差异无统计学意义。
结论:本研究发现采用UAS-GAL4系统构建的Mhc>PINK1-RNAi转基因PD果蝇在常温25 ℃培养条件下,PD果蝇模型成立,表现出典型的PD症状:PINK1基因表达、多巴胺含量和攀爬能力显著下降和异翅率显著提高。因此,建议采用该模型开展科学研究时,应在25 ℃条件下培养。本研究结果将为基于该模型的帕金森疾病的病理机制、药效、药理学研究提供科学依据。
猜你喜欢
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
读者(2022年9期)2022-04-22
学苑创造·A版(2022年3期)2022-03-29
老年医学研究(2021年5期)2022-01-19
烟台果树(2021年2期)2021-07-21
疯狂英语·新读写(2020年3期)2020-06-06
时代英语·高一(2019年5期)2019-09-03
学苑创造·A版(2019年6期)2019-07-11
测控技术(2018年7期)2018-12-09
