
外源γ-氨基丁酸对蛇龙珠葡萄果实糖酸代谢的影响
2023-06-12 23:17王宇航韩爱民张立梅李斗金鑫王春恒冯丽丹杨江山
果树学报 2023年4期
关键词:氨基丁酸
王宇航 韩爱民 张立梅 李斗 金鑫 王春恒 冯丽丹 杨江山


摘 要:【目的】探讨外源γ-氨基丁酸(GABA)对葡萄果实糖酸积累及代谢的影响。【方法】以蛇龙珠葡萄为试材,于开 花期、坐果期、膨大期和转色期分别用5 mmol·L(-1 T1)、10 mmol·L(-1 T2)、15 mmol·L(-1 T3)、20 mmol·L(-1 T4)的GABA 溶液进行叶面喷施,以喷施蒸馏水作为对照(CK),研究其对葡萄果实发育过程中糖、酸组分含量以及相关代谢酶活性 的影响。【结果】叶面喷施GABA溶液能显著提高葡萄果实可溶性糖含量,T2处理显著提高蔗糖、葡萄糖和果糖含量, 蔗糖合成酶合成方向(SuSy-s)、蔗糖磷酸合成酶(SPS)、酸性转化酶(AI)及中性转化酶(NI)活性均显著提高。GABA 处理均能显著提高谷氨酸脱羧酶(GAD)活性和果实GABA含量。适宜浓度GABA处理能显著提高葡萄果实草酸、酒 石酸、富马酸、柠檬酸、α-酮戊二酸、苹果酸含量,酸代谢相关酶活性均显著提高。GABA与NI、柠檬酸呈极显著正相 关,与SPS、草酸、线粒体乌头酸酶(Mit-ACO)呈显著正相关。【结论】外源GABA通过提高蔗糖合成相关酶、转化酶活 性和有机酸代谢酶活性,提高了葡萄果实葡萄糖、果糖、苹果酸、酒石酸等有机酸含量,进而改善了葡萄风味品质。
关键词:蛇龙珠葡萄;γ-氨基丁酸;蔗糖代谢;有机酸代谢
中图分类号:S663.1 文献标志码:A 文章编号:1009-9980(2023)04-0699-1
糖和有机酸是影响果实品质的重要代谢物质, 与挥发性物质一起决定果实的风味品质[1] 。酿酒葡 萄适宜的糖和酸含量是决定葡萄酒风味品质的重要 因素,葡萄果实中最主要的可溶性糖是葡萄糖和果 糖,有机酸主要为酒石酸和苹果酸,此外还含有少量 的琥珀酸、草酸和柠檬酸,糖和有机酸的合成与积累 主要受蔗糖代谢相关酶和三羧酸循环相关酶的调 控[2] 。目前生产中主要通过温度、光照、水分、土壤 等环境因素的调控和栽培措施来提高葡萄果实糖分 和有机酸的积累[3] 。
γ-氨基丁酸(GABA)是一种广泛存在于植物体 内的四碳非蛋白氨基酸,GABA支路途径是GABA 通过谷氨酸脱羧酶(GAD)生物合成,并绕过TCA循 环中2种关键酶α-酮戊二酸脱氢酶(α-KGDH)和琥 珀酰辅酶A连接酶(SCS)的反应,最后以琥珀酸的 形式返回三羧酸循环[4] 。研究表明,外源 GABA 处 理可以提高采后苹果果实中草酸、苹果酸、乌头酸、 琥珀酸的水平[5] 。Faraj等[6] 的研究结果表明GABA 處理提高了柑橘叶片中琥珀酸水平,并且提升了 GABA 转 氨 酶(GABA- T)、琥 珀 酸 半 醛 脱 氢 酶 (SSADH)、苹果酸脱氢酶和琥珀酸脱氢酶等的活 性;GABA在碳代谢与氮代谢之间也发挥重要作用, 谷氨酸脱羧酶(GAD)作为GABA合成的关键酶,在 GAD基因敲除的拟南芥中发现,拟南芥种子中可溶 性糖和有机酸含量明显下降[7-8] 。但外源 GABA 在 葡萄果实发育过程中对果实糖酸积累及代谢的影响 鲜有报道。因此,笔者在本研究中以10年生酿酒葡 萄蛇龙珠为试材,对葡萄叶片进行不同浓度GABA 处理,通过测定葡萄果实糖酸组分、糖酸代谢及GA BA 合成相关酶活性,探讨 GABA 对葡萄果实发育 过程中糖酸代谢机制和GABA代谢的影响,以期为 酿酒葡萄优质生产提供理论依据。
1 材料和方法
1.1 试验材料
试验在甘肃农业大学食品科学与工程学院露地 葡萄园进行,供试品种为10年生酿酒葡萄品种蛇龙 珠,株行距为0.75 m×1.5 m,单干双臂Y形整形,南 北走向。
1.2 试验设计
试 验 共 设 4 个 GABA 浓 度 处 理 :5(T1)、10 (T2)、15(T3)、20 mmol·L(-1 T4),以蒸馏水处理为对 照(CK),于盛花期、坐果期、膨大期、转色期对长势 一致、无病虫害的植株进行叶面喷施,以叶片开始滴 液为准,每个处理设3个重复,每个重复5株。果实 采样时间为处理后第3天上午8:00,成熟期再取样1 次,每株随机选取1穗葡萄,冰盒贮运至实验室,液 氮冷冻后放入超低温冰箱备用。
1.3 果实可溶性总糖、糖组分含量及蔗糖代谢相关 酶活性测定
可溶性总糖含量采用蒽酮硫酸法测定[9] ,使用高 效液相色谱仪(美国Waters Acquity Arc)参照贺雅娟 等[10] 的方法进行果实蔗糖、葡萄糖和果糖含量的测 定。葡萄果肉加液氮研磨后称取 0.5 g,加入 5 mL 80%乙醇,35 ℃超声提取20 min,12 000 r·min-1 离心 15 min,重复提取 2 次,每次加 80%乙醇 2 mL,合并 上清液,定容至10 mL,真空离心浓缩仪旋转蒸发至 全干(60 ℃),用 1 mL 超纯水 1 mL 乙腈复溶,用 0.22 μm 有机相微孔滤膜过滤后加入样品瓶中待 测。色谱条件:XBridge BEH Amide色谱柱(4.6 mm× 150 mm,2.5 μm),柱温 40 ℃,流动相为 75%乙腈、 0.2%乙胺以及 24.8%超纯水,流速 0.8 mL·min-1 ,进 样量 10 μL,检测波长为 254 nm。蔗糖、葡萄糖和 果糖的标准曲线分别为y=110 269x-9 598.4,R2 = 0.999 3、y=110 132x-9 849.6,R2 =0.999 4、y=104 844x-10 395,R2 =0.999 2。
SuSy-s(蔗糖合酶合成方向)、SuSy-c(蔗糖合酶 分解方向)、SPS(蔗糖磷酸合成酶)、AI(酸性转化 酶)、NI(中性转化酶)酶液提取参考文献[11]的方 法,酶活性测定参考文献[12]的方法。以鲜质量计。
1.4 果实GAD、GABA、有机酸组分含量及有机酸 代谢相关酶活性测定
GAD 酶液提取及酶活性测定采用文献[13]的 方法,GABA 含量测定采用文献[14]的方法。果实 草酸、酒石酸、莽草酸、富马酸、柠檬酸、α-酮戊二 酸、苹果酸含量测定参考文献[15]的方法,葡萄果 肉加液氮研磨后称取 1.5 g,加入 7.5 mL 超纯水, 4 ℃、10 000 r·min-1 离心 10 min,用 0.22 μm 水相微 孔濾膜过滤,将滤液加入样品瓶中待测,使用美国 Waters Acquity Arc高效液相色谱仪,亚特兰蒂斯T3 柱(4.6 mm×150 mm,3 μm),流动相为 20 mmol·L-1 NaH2PO4 溶液(用 H3PO4 将 pH 调至 2.7),流速为 0.50 mL·min-1 ,柱温为 30 ℃,检测波长为 210 nm, 进样量为 20 μL。草酸、酒石酸、莽草酸、富马酸、柠檬酸、α-酮戊二酸、苹果酸的标准曲线分别为y=28 071 704.260 7x-2 742.293 2,R2 =0.999 4、y= 4 250 792.452 8x-3 184.606 5 ,R2 =0.999 1 、y= 102 070x-30 412,R2 =0.997 9、y=174 769.779 9x+ 13 427.704 9 ,R2 =0.999 8 、y=1 877 841.095 9x- 2 096.561 6,R2 =0.998 6、y=18 989x-43 769,R2 =0.999 5、y=1 721 625.744 2x+5 796.673 9,R2 =0.999 9。
CS(柠檬酸合酶)、Cyt-ACO(细胞质乌头酸 酶)、Mit-ACO(线粒体乌头酸酶)、NAD-IDH(NAD- 异柠檬酸脱氢酶)、PEPC(磷酸烯醇式丙酮酸羧化 酶)、NAD-MDH(NAD-苹果酸脱氢酶)、NADP-ME (NADP-苹果酸酶)酶液提取及活性测定参考文献 [16-17]的方法,以鲜质量计。
1.5 数据统计分析
采用 SPSS 23.0 进行数据处理,方差分析采用 Duncans多重比较。采用Excel进行图表绘制。
2 结果与分析
2.1 GABA对葡萄果实可溶性总糖及糖组分含量的 影响
如图1-A所示,在果实发育过程中,膨大期和转 色期可溶性总糖含量迅速增加,成熟期增加缓慢;膨 大期GABA处理葡萄可溶性总糖含量较CK提高效 果最明显,其中T3处理较CK提高了0.81倍;成熟期 GABA 处理均显著提高了可溶性总糖含量(p< 0.05),其中 T2 处理较 CK 提高了 0.09 倍,说明外源GABA处理有利于果实可溶性总糖的积累。 坐果期和膨大期蔗糖(图 1-B)、葡萄糖(图 1- C)、果糖(图 1-D)含量缓慢增加,随着葡萄果实成 熟,葡萄糖、蔗糖、果糖含量迅速增加。成熟期GA BA 处理均显著提高了葡萄糖含量(p<0.05),其中 T2处理效果最好,较CK提高了0.33倍;成熟期T2、 T3和T4处理蔗糖含量较CK均显著提高(p<0.05), 其中T2处理提高效果最为显著,较CK提高了0.63 倍;成熟期 T1 和 T2 处理果糖含量提高显著(p< 0.05),分别较CK提高了0.32倍、0.48倍。
2.2 GABA对葡萄果实蔗糖代谢酶活性的影响
在果实发育过程中CK处理SuSy-s活性呈先升 高后降低的变化趋势,葡萄转色期达到最高(图 2- A)。经过GABA处理后,各处理SuSy-s活性随着时间的推移,均显著高于CK,其中T2处理效果最为显 著(p<0.05),较CK坐果期、膨大期、转色期和成熟 期分别提高了1.79、0.59、0.45、1.11倍。
如图2-B所示,CK处理SPS活性在果实发育过 程中呈波动型变化趋势,在膨大期缓慢上升,在转色 期到达最低点,随后又开始上升。坐果期GABA处 理均显著提高了 SPS 活性(p<0.05),分别是 CK 的 0.83、1.37、1.76、1.71倍;从膨大期至成熟期,T2处理 和T3处理SPS活性较CK提高显著且波动较小,转 色期效果最为显著(p<0.05),分别是 CK 的 1.25、 1.01 倍。成熟期 GABA 处理 SPS 活性均显著高于 CK(p<0.05),但不同浓度GABA处理差异不明显。 SuSy-c活性(图2-E)在坐果期最高,随着果实的 发育呈迅速下降-缓慢升高-迅速下降趋势,坐果期 GABA处理均显著增强了SuSy-c的活性(p<0.05), 其中,T1处理效果最好,较CK提升了0.55倍;但从 膨大期开始至成熟期,GABA处理表现出明显的抑 制作用,膨大期和转色期T4处理的抑制作用表现最 强,成熟期T3处理抑制作用最显著(p<0.05),较CK 降低了17%。说明T2处理能够提高SuSy-s、SPS活 性,降低SuSy-c活性,有利于葡萄果实蔗糖大量合成 和积累,而T3处理抑制蔗糖分解作用最强。
AI(图2-C)和NI(图2-D)活性随着果实发育均 逐渐升高,成熟期AI活性迅速降低,而NI活性降低 缓慢。膨大期、转色期GABA处理AI活性均显著提 升,其中膨大期 T3 处理效果最显著(p<0.05),较 CK 提高了 0.95 倍 ,转色期 T2 处理最显著(p< 0.05),较CK提高了0.40倍,成熟期各处理间差异均 不显著。膨大期和转色期各处理NI活性均没有表 现出显著差异,成熟期T3处理效果显著(p<0.05), 较 CK 提高了 0.22 倍。AI 和 NI 活性的提高提升了 蔗糖转化为葡萄糖和果糖的速率,从而促进了葡萄 糖和果糖的积累。
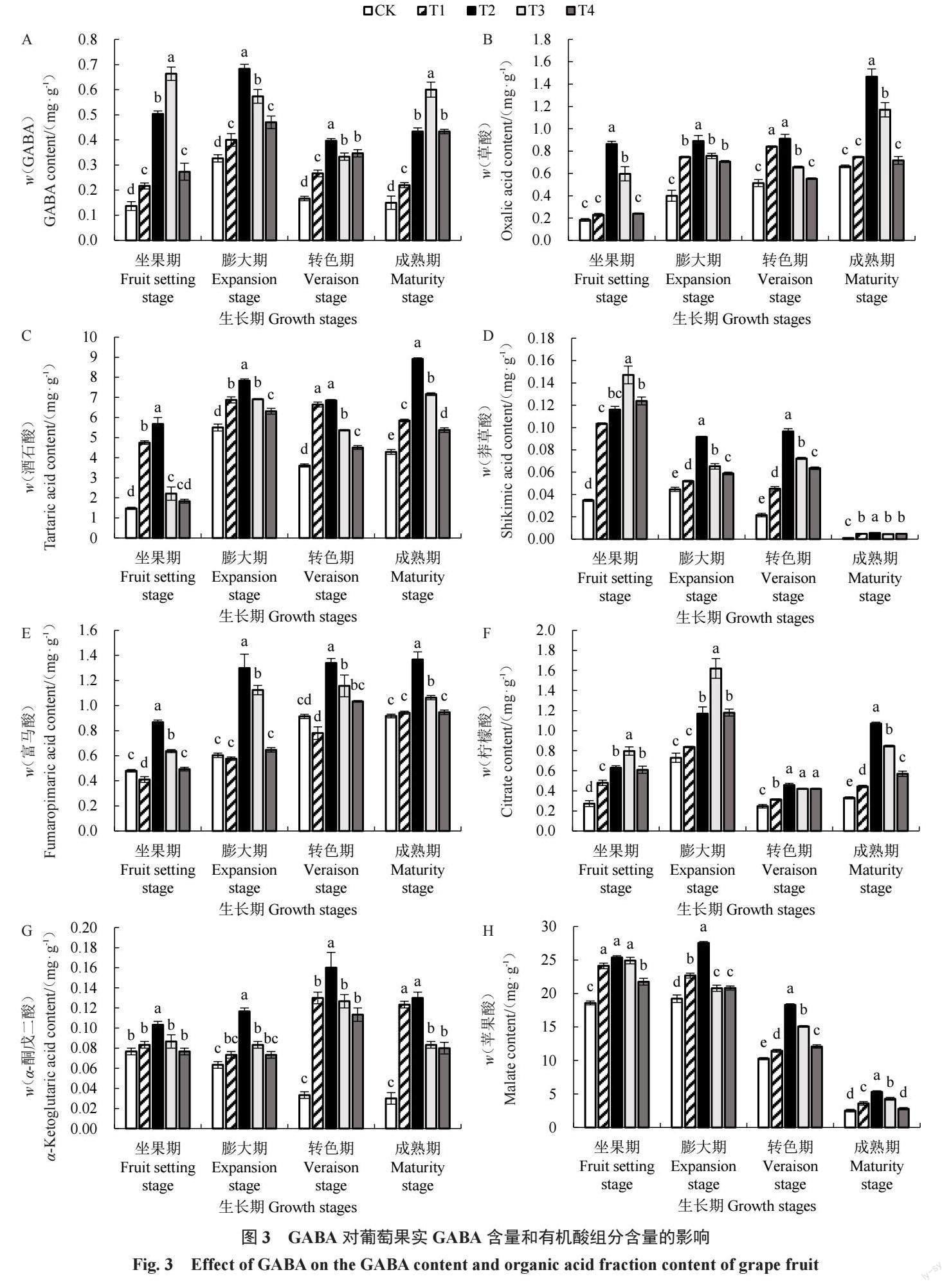
2.3 GABA对葡萄果实GABA含量和有机酸组分 含量的影响
如图3-A所示,CK处理葡萄GABA含量在果实 发育过程中呈先升高后降低的趋势,经过GABA处 理后,各处理 GABA 含量随着时间的推移,均显著 高于对照(p<0.05),其中坐果期 T3 处理对 GABA 含量提升的效果最显著,是CK的3.91倍,说明外源 GABA处理能提高葡萄果實GABA含量。
草酸含量随着果实成熟逐渐升高(图 3-B),而 酒石酸含量在果实发育过程中呈现出先升高后略微 降低再升高的趋势(图 3-C)。经过对 GABA 处理 后 ,成熟期草酸及酒石酸含量均显著提高(p< 0.05),其中T2处理的效果最为显著,分别是CK的 1.21、1.08倍,说明不同浓度GABA处理均能提高草 酸及酒石酸含量。
莽草酸(图 3-D)含量呈先升高后降低的趋势, 坐果期各处理莽草酸含量达到峰值,其中T3处理莽 草酸含量最高,是 CK 的 3.23 倍,但随着果实的成 熟,各处理莽草酸含量均较低且差异较小;而富马酸 含量(图 3-E)呈缓慢升高的趋势,转色期和成熟期 变化较小,成熟期GABA处理显著提高了富马酸含 量(p<0.05),以T2处理效果最显著,富马酸含量较 CK 提升了 0.49 倍;说明适宜浓度 GABA 处理有利 于莽草酸及富马酸的积累。
柠檬酸含量(图3-F)随着果实的发育呈先迅速 升高、略微降低后再升高的趋势,在膨大期达到峰 值,成熟期 GABA 处理均显著提高了柠檬酸含量 (p<0.05),其中T2处理效果最好,是CK的2.26倍, GABA处理能促进果实柠檬酸的积累。
在果实发育过程中,CK 处理α-酮戊二酸含量 (图3-G)呈逐渐降低的趋势,但GABA处理呈先上 升后下降的趋势,转色期达到峰值,其中T2处理效 果最为显著(p<0.05),转色期T2处理是CK的3.74 倍,成熟期T2处理是CK的3.41倍。
如图3-H所示,CK处理苹果酸含量随着果实发 育呈逐渐降低的趋势,但T2处理呈先上升后下降的 趋势,与CK相比,T2处理均显著提高了苹果酸含量 (p<0.05),其中膨大期T2处理苹果酸含量最高,随 着果实成熟迅速降低,成熟期T2处理是CK的1.11 倍。
2.4 GABA对葡萄果实GAD及有机酸代谢相关酶 活性的影响
GAD可以通过谷氨酸合成GABA。如图4-A所 示,在果实发育过程中,CK处理葡萄GAD活性缓慢 降低,经过GABA处理后,各处理GAD活性随着时 间的推移,均显著高于CK(p<0.05),且降低幅度较 小,其中,成熟期T4处理提升效果最为显著,较CK 提高了0.50倍。
如图4-B所示,CK处理CS活性呈缓慢降低-迅 速升高-迅速降低的趋势,转色期活性最高,成熟期 迅速降低。坐果期、膨大期和转色期GABA处理CS 活性均显著高于CK(p<0.05),其中膨大期T3处理 的提升效果最显著,CS活性是CK的3.95倍。
ACO催化柠檬酸产生水和顺乌头酸,Cyt-ACO 和Mit-ACO是植物中的2种同工酶。如图4-C、D所 示,Cyt-ACO活性在坐果期最高,随着果实发育逐渐 降低,各处理Mit-ACO活性在果实发育过程中均呈 迅速降低-缓慢上升的变化趋势,转色期 Mit-ACO 活性最低。坐果期 GABA 处理均显著提高了 Cyt ACO和Mit-ACO活性(p<0.05),其中T3处理效果 最为显著,分别是CK的2.90和1.67倍。ACO活性 的提高可加快柠檬酸的利用,但ACO活性较CS活 性低,有利于柠檬酸积累,NAD-IDH也是柠檬酸降 解的控制因子,NAD-IDH活性(图4-E)在果实发育 过程中呈先缓慢降低后迅速升高的变化趋势,成熟 期 NAD-IDH 活性最高。NAD-IDH 活性成熟期 T4 处理提升效果最为显著(p<0.05),是CK的1.58倍。
如图 4-F 所示,PEPC 活性随果实成熟逐渐上 升,成熟期酶活性最高且各处理均差异显著(p< 0.05),其中,T2处理提升效果最为显著,较CK提高 了0.86倍。NAD-MDH活性在果实发育过程中先迅 速升高后缓慢升高(图4-G);成熟期T2、T3和T4处 理 NAD-MDH 活性均与 CK 差异显著(p<0.05),其T4 处理提升效果最显著,较 CK 提高了 0.41 倍。 CK处理NADP-ME活性在果实发育过程中逐渐升 高(图 4-H)。GABA 处理与 CK 相比均显著提升了 NADP-ME 活性(p<0.05),以膨大期提升效果最显著,分别是CK的1.44、1.14、1.27、1.51倍,但不同浓 度GABA处理的效果不显著。
2.5 葡萄果实发育过程中GABA代谢与糖组分含 量及糖代谢相关酶活性的相关性
利用SPSS软件对葡萄果实GABA含量、GAD、 糖组分含量及糖代谢相关酶活性进行相关性分析, 获得相关系数矩阵(表 1)。结果表明,GABA 与 GAD、NI呈极显著正相关(p<0.01),与SPS呈显著 正相关(p<0.05)。蔗糖与可溶性糖、葡萄糖、果糖、 SuSy-s 呈极显著正相关(p<0.01),与 SuSy-c、NI 呈 极显著负相关(p<0.01)。葡萄糖和果糖与可溶性 糖 、蔗糖 、SuSy- s 呈极显著正相关(p<0.01),与SuSy-c、NI 呈极显著负相关(p<0.01),与 AI 呈显 著负相关(p<0.05)。
2.6 不同浓度 GABA 处理对葡萄果实发育过程 中GABA代谢与有机酸组分含量及有机酸代谢相 关酶活性的相关性
利用SPSS软件对葡萄果实GABA含量、GAD 活性、有机酸组分含量及有机酸代谢相关酶活性 进行相关性分析,获得相关系数矩阵(表2)。结果 表明,GABA与GAD、柠檬酸呈极显著正相关(p< 0.01),与 草 酸 、Mit- ACO 呈 显 著 正 相 关(p< 0.01)。草酸與富马酸、酒石酸、IDH、MDH、ME、 PEPC呈极显著正相关(p<0.01),与GABA、CS呈 显著正相关(p<0.05)。富马酸与草酸、酒石酸、α- 酮戊二酸、CS、MDH、ME、PEPC 呈极显著正相关 (p<0.01)。酒石酸与草酸、富马酸、ME呈极显著 正相关(p<0.01),与柠檬酸、MDH呈显著正相关 (p<0.05)。莽草酸与 GAD、苹果酸、Cyt-ACO、 Mit- ACO 呈极显著正相关(p<0.01),与 MDH、 PEPC 呈显著负相关(p<0.05)。柠檬酸与 GABA 呈极显著正相关(p<0.01),与酒石酸呈显著正相关 (p<0.05)。苹果酸与 GAD、莽草酸、MDH、PEPC 呈 极显著正相关(p<0.01),与 Mit-ACO 呈显著正相关 (p<0.05),与CS、IDH、ME呈显著负相关(p<0.05)。α-酮戊二酸与富马酸呈极显著正相关(p<0.01),与 GAD、草酸、CS呈显著正相关(p<0.05)。
3 讨 论
糖、有机酸是构成果实风味物质的重要组成部 分,而在酿酒葡萄中,糖和有机酸含量是影响葡萄酒 品质的重要因素。研究表明,酿酒葡萄中的糖类大部 分转化为酒精,约10%转化为脂类和酚酸类物质,蔗 糖、葡萄糖及果糖含量的提高可以增加葡萄酒的风味 强度,而有机酸种类及含量的变化不仅对葡萄酒的口 感、色泽及稳定性都有影响,还具有调节酸碱平衡的 作用[18] ;张扬等[19] 研究表明酒石酸等有机酸含量的提 高可以增强葡萄酒的色泽和香气。在蛇龙珠葡萄发 育过程中,各种糖类物质含量逐渐上升,有机酸含量 逐渐降低,在果实成熟期,蛇龙珠葡萄中葡萄糖含量略高于果糖含量,蔗糖含量较低,有机酸中酒石酸和 苹果酸含量最高,10 mmol·L-1 GABA处理后,蔗糖、 葡萄糖及果糖含量均显著高于对照,且果糖含量略 高于葡萄糖含量,酒石酸、苹果酸、柠檬酸等有机酸 含量也显著提高。因此,外源GABA处理提高了酿 酒葡萄糖和有机酸含量,使酿酒葡萄果实达到适宜 的糖酸含量,可能是提高葡萄酒品质的重要措施。
研究表明,外源喷施GABA通过提高正常条件 和低温条件下番茄叶片叶绿素含量,提高抗氧化酶 活性和叶片净光合速率,从而提高可溶性糖、还原糖 及非还原糖含量[20] 。GAD 作为 GABA 支路中 GA BA合成的关键酶,拟南芥GAD突变体种子表现出 部分糖和有机酸含量降低[8] 。在本研究中,通过喷 施不同浓度的GABA,各处理均表现出可溶性糖、葡 萄糖、果糖、蔗糖含量增加。
在植物中,蔗糖、葡萄糖及果糖的积累主要由 SuSy、SPS、AI 及 NI 共同调控 ,蔗糖的合成通过 SuSy和SPS催化,而葡萄糖和果糖的积累则来自蔗 糖的分解,由转化酶和SuSy催化[21] 。在蛇龙珠葡萄 果实发育过程中,在果实发育初期SPS活性较高,在 果实发育后期活性下降,SuSy-s活性则在果实发育 的中后期迅速升高,而果实中的蔗糖含量随着果实 的成熟逐渐升高,因此,SuSy可能是决定果实蔗糖 积累的重要因素。通过相关性分析发现,蔗糖含量 与SuSy-s活性呈极显著正相关,与SuSy-c活性、NI 活性呈极显著负相关,说明SuSy可能是调控果实蔗 糖含量的关键酶,而NI可能是果实成熟过程中调控 蔗糖分解的关键酶;这与寇单单等[22] 的结果一致。 通过分析发现,GABA含量与SPS活性呈显著正相 关,GABA可能通过影响SPS活性影响蔗糖含量,但 其调控机制仍需进一步研究。外源 GABA 处理显 著提高了葡萄糖和果糖的水平和转化酶活性,而 GABA处理在果实发育中后期,SuSy-c活性在果实 发育过程中呈缓慢降低的趋势,且酶活性均较低, AI活性在果实发育过程中逐渐升高,且活性较高, AI可能在调控葡萄糖和果糖积累过程中发挥关键 作用,与文献[23-24]结论一致。通过相关分析发现, GABA含量与NI活性呈极显著正相关,说明GABA 也可能通过提高NI活性,从而影响蔗糖代谢。研究 表明,蔗糖可作为信号分子调控基因的表达,从而影 响酶催化活性[25] 。蔗糖水平的提高可能提高了转化 酶活性,进而促进了葡萄糖和果糖的积累[26] 。本研 究结果显示外源GABA处理提高了蔗糖合成相关酶 的活性,降低SuSy-c活性,促进了蔗糖的积累,通过 提高转化酶活性提高了葡萄糖和果糖含量。但其调 控机制仍需进一步研究。
谷氨酸脱羧酶(GAD)是谷氨酸合成GABA的限 速酶,外源 GABA 通过提高 GAD、GABA-T 活性和CmGAD基因表达量,促进GABA的生物合成,进而增 强了GABA支路的效率[27- 28] 。这与本研究的结果一 致,外源GABA条件下,GAD活性和GABA含量显著 提高,相关性分析表明,GABA含量与GAD活性呈极 显著正相关,说明GAD可能是GABA合成的关键酶。 GABA通过GABA支路连接三羧酸循环,而三 羧酸循环是果实中碳水化合物、有机酸和氨基酸代 谢的桥梁[29] 。果实中各种有机酸的积累取决于有机 酸的合成、降解和利用的平衡[30] 。酒石酸、苹果酸、 琥珀酸、草酸和柠檬酸是影响葡萄果实风味的主要 有 机 酸 ,受 PEPC、NADP- ME、NADP- IDH、Cyt ACO、NAD-MDH和CS直接或间接调节。PEPC将 磷酸烯醇丙酮酸催化为草酰乙酸,随后在 NAD MDH 存在下降解为苹果酸或通过 CS 降解为柠檬 酸,而苹果酸可通过 NADP-ME 进一步降解为丙酮 酸,柠檬酸盐可通过ACO和NAD-IDH分解为α-酮 戊二酸和二氧化碳[31] 。喷施使用同位素标记的GA BA 发现,GABA 能快速转化为琥珀酸并进入 TCA 循环[32] 。有学者发现外源 GABA 能提高 TCA 循环 的速率,增强酸代谢相关酶活性,从而促进了有机酸 的积累[6,33] 。Shoukun[5] 认为GABA处理的果实可加 速苹果酸的生物合成和抑制其分解 ,通过上调MdGAD、MdGABA-T和MdSSADH,促进GABA分流 的活性,这可能导致琥珀酸和GABA的积累。笔者 在本研究中发现外源GABA增加了果实中CS、Cyt ACO、Mit- ACO、NAD- IDH、PEPC、NAD- MDH、 NADP-ME的活性,果实中草酸、酒石酸、莽草酸、富 马酸、柠檬酸、α-酮戊二酸和苹果酸含量也显著提 高,通过相关性分析发现,GABA含量与柠檬酸含量 呈极显著正相关,与草酸含量呈显著相关,说明GA BA处理能促进柠檬酸和草酸的积累。可能是外源 喷施 GABA 后,能使果实中 GABA 含量迅速升高, GABA 通过 GABA 支路转化为琥珀酸进入 TCA 循 环,琥珀酸含量的增加通过负反馈调节使α-酮戊二 酸含量增加,从而降低柠檬酸的分解速率,促进了柠 檬酸的积累;而琥珀酸含量的增加也可提高苹果酸合成的底物水平,从而促进苹果酸的积累;TCA 循 环速率的提高为其他中间代谢产物提供了较高水平 的底物,促进了草酸、酒石酸、莽草酸、富马酸的积 累。有研究表明,GABA调节ALMT(铝活化阴离子 蛋白)的活性来引发跨膜电位差的变化,从而影响三 羧酸循环的中间代谢产物[34] 。Alexis[35] 的研究表明VvALMT9 能够介导向内整流苹果酸盐和酒石酸盐 电流,促进这些二羧酸在葡萄浆果液泡中的积累。
4 结 论
研究结果表明,外源GABA通过提高SuSy-s和 SPS活性,抑制SuSy-c活性来促进蔗糖的积累,通过 提高转化酶活性来促进葡萄糖和果糖的积累。外源 喷施GABA通过提高GAD活性来促进GABA的生 物合成,从而提高TCA循环速率,柠檬酸合酶、细胞 质乌头酸酶和线粒体乌头酸酶、NAD-异柠檬酸脱氢 酶、磷酸烯醇式丙酮酸羧化酶、NAD-苹果酸脱氢酶 和 NADP-苹果酸酶活性均显著提高,促进了草酸、 酒石酸、莽草酸、富马酸、柠檬酸、α-酮戊二酸和苹果 酸的积累,进而改善了葡萄风味品质,以叶面喷施 10 mmol·L-1 GABA效果最好。
参考文献 References:
[1] JULIA B,CLAUDIO O B,LUC?A P,MARTIN A L, VER?NICA A L,RICARDO M,CARLOS S A,MAR?A F D, MAR?A V L. Carbon metabolism of peach fruit after harvest: changes in enzymes involved in organic acid and sugar level modifications[J]. Journal of Experimental Botany,2009,60(6): 1823-1837.
[2] CRYSTAL S,LAURENT G D,GRANT R C,CHRISTOPHER M F,KATHLEEN L S. Regulation of malate metabolism in grape berry and other developing fruits[J]. Phytochemistry, 2009,70(11):1329-1344.
[3] 蘇静,祝令成,刘茜,彭云静,马百全,马锋旺,李明军 . 果实糖 代谢与含量调控的研究进展[J]. 果树学报,2022,39(2):266- 279.
SU Jing,ZHU Lingcheng,LIU Xi,PENG Yunjing,MA Bai quan,MA Fengwang,LI Mingjun. Research progress on sugar metabolism and concentration regulation in fruit[J]. Journal of Fruit Science,2022,39(2):266-279.
[4] ALAN W B,BARRY J S. Does the GABA shunt regulate cyto solic GABA?[J]. Trends in Plant Science,2020,25(5):422-424.
[5] SHOUKUN H,YUYU N,WEI Q,YIHENG H,QIUYAN B, YANRONG L,JINGPING R. Exogenous γ- aminobutyric acid treatment that contributes to regulation of malate metabolism and ethylene synthesis in apple fruit during storage[J]. Journal of Agricultural and Food Chemistry,2018,66(51):13473- 13482.
[6] FARAJ H,NABIL K. Exogenous GABA is quickly metabolized to succinic acid and fed into the plant tca cycle[J]. Plant Signal ing & Behavior,2019,14(3):e1573096.
[7] LI L,DOU N,ZHANG H,WU C X. The versatile GABA in plants[J]. Plant Signaling & Behavior,2021,16(3):1862565.
[8] AARON F,ADRIANO N N,RUTHIE A,MARTIN L,PHU ONG A P,LUHUA S,RICHARD P H,JOHNATHAN A N, GAD G,ALISDAIR R F. Targeted enhancement of glutamate-to-γ-aminobutyrate conversion inArabidopsis seeds affects carbon nitrogen balance and storage reserves in a development- depen dent manner[J]. Plant Physiology,2011,157(3):1026-1042.
[9] 刘晓涵,陈永刚,林励,庄满贤,方晓娟 . 蒽酮硫酸法与苯酚硫 酸法测定枸杞子中多糖含量的比较[J]. 食品科技,2009,34 (9):270-272.
LIU Xiaohan,CHEN Yonggang,LIN Li,ZHUANG Manxian, FANG Xiaojuan. Comparison of methods in determination of polysaccharide inLycium barbarum L. [J]. Food Science and Technology,2009,34(9):270-272.
[10] 贺雅娟,马宗桓,韦霞霞,李玉梅,李彦彪,马维峰,丁孙磊,毛 娟,陈佰鸿 . 黄土高原旱塬区不同品种苹果果实糖及有机酸 含量比较分析[J]. 食品工业科技,2021,42(10):248-254.
HE Yajuan,MA Zonghuan,WEI Xiaxia,LI Yumei,LI Yanbiao, MA Weifeng,DING Sunlei,MAO Juan,CHEN Baihong. Com parative analysis of sugar and organic acid contents of different apple cultivars in dryland of loess plateau[J]. Science and Tech nology of Food Industry,2021,42(10):248-254.
[11] 张弦 . 不同施钾水平对‘嘎拉苹果果实糖、酸生理代谢的影 响[D]. 杨凌:西北农林科技大学,2016.
ZHANG Xian. Effects of different potassium level on sugar and acid metabolism in Gala apple fruit[D]. Yangling:Northwest A&F University,2016.
[12] 潘俨 . 库尔勒香梨果实发育及采后糖代谢与呼吸代谢关系的 研究[D]. 乌鲁木齐:新疆农业大学,2016.
PAN Yan. The relationship between sugar metabolism and respi ratory metabolism throughout fruit development and postharvest of Korla Fragrant pear (Pyrus Sinkiangensis Yu) [D]. Urumqi: Xinjiang Agricultural University,2016.
[13] PHAIWAN P,PARITA T,TIPAWAN T,PANATDA J,FENG C,SUDARAT J. Glutamate decarboxylase (gad) extracted from germinated rice: enzymatic properties and its application in soy milk[J]. Journal of Nutritional Science and Vitaminology,2019, 65:S166-S170.
[14] PHUONG H L,LIESA V,THIEN T L,YANNICK V, KATLEEN R. Implementation of hplc analysis for γ-aminobutyr ic acid (GABA) in fermented food matrices[J]. Food Analytical Methods,2020,13(5):1190-1201.
[15] 李彥彪,马维峰,贾进,牟德生,李生保,毛娟 . 河西走廊不同产 地‘赤霞珠酿酒葡萄果实品质评价[J]. 西北植物学报,2021, 41(5):817-827.
LI Yanbiao,MA Weifeng,JIA Jin,MU Desheng,LI Shengbao, MAO Juan. Evaluation on fruit quality of Cabernet Sauvignon wine grapes from different producing areas in hexi corridor[J]. Acta Botanica Boreali Occidentalia Sinica,2021,41(5):817- 827.
[16] 罗安才 . 柑橘果实有机酸代谢生理和奉节脐橙芽变株系的 AFLP 分析研究[D]. 重庆:西南农业大学,2003.
LUO Ancai. Research on the organic acids metabolism in citrus fruits and the AFLP analysis of the fengjie navel orange mu tants[D]. Chongqing: Southwest Agricultural University,2003.
[17] 刘丽媛 . 山葡萄糖酸积累规律及代谢调控机理研究[D]. 杨凌: 西北农林科技大学,2016.
LIU Liyuan. The physiological study on the sugar and acid accu mulations and metabolic regulation mechanisms ofVitis amuren sis Rupr. grape[D]. Yangling:Northwest A & F University, 2016.
[18] 曹炜玉,路文鹏,舒楠,杨义明,范书田 . 葡萄酒风味物质及其 影响因素研究进展[J]. 中国酿造,2022,41(5):1-7.
CAO Weiyu,LU Wenpeng,SHU Nan,YANG Yiming,FAN Shutian. Research progress on wine flavor substances and their influencing factors[J]. China Brewing,2022,41(5):1-7.
[19] 张扬,彭晶晶,李坤一,杨洁,郭安鹊 . 发酵前添加有机酸对 ‘西拉红葡萄酒颜色和感官质量的影响[J/OL]. 食品与发酵 工 业 :[2022- 08- 20]. https://kns.cnki.net/kcms/detail/11.1802. TS.20220712.1408.009. html.
ZHANG Yang,PENG Jingjing,LI Kunyi,YANG Jie,GUO Anque. Effects of organic acid added before fermentation on col or and sensory quality of Syrah red wine[J/OL]. Food and Fer mentation Industries:[2022- 08- 20]. https://kns.cnki.net/kcms/ detail/11.1802.TS.20220712. 1408.009.html.
[20] OLA H AE,AMR E,GNIEWKO N,REHAM F,TOMASZ W, SOUMYA M,AYMAN F A,HUSSIEN M E,AHMED A E,HA NY G A E,EHAB A,ADIL A G,NIHAL E N,AHMED M E, AHMED B,MOHAMED F M I. Protective effect of γ-aminobu tyric acid against chilling stress during reproductive stage in to mato plants through modulation of sugar metabolism, chloro plast integrity, and antioxidative defense systems[J]. Frontiers in Plant Science,2021,12:663750.
[21] MUHAMMAD J U,LUQMAN B S,HAILESLASSIE G, SHENGJIE Z,PINGLI Y,HONGJU Z M O K,MUHAMMAD A,XUQIANG L,NAN H,CHENGSHENG G,WENGE L. Iden tification of key gene networks controlling organic acid and sug ar metabolism during watermelon fruit development by integrat ing metabolic phenotypes and gene expression profiles[J]. Horti culture Research,2020,7(1):193.
[22] 寇單单,张叶,王朋飞,李东东,张学英,陈海江 .‘仓方早生 桃及其早熟芽变果实蔗糖和苹果酸积累与相关基因表达[J]. 园艺学报,2019,46(12):2286-2298.
KOU Dandan,ZHANG Ye,WANG Pengfei,LI Dongdong, ZHANG Xueying,CHEN Haijiang. Differences in sucrose and malic acid accumulation and the related gene expression in ‘Kurakato Wasepeach and its early- ripening mutant[J]. Acta Horticulturae Sinica,2019,46(12):2286-2298.
[23] 龚荣高,张光伦,吕秀兰,曾秀丽,罗楠,胡强 . 脐橙在不同生 境下果实蔗糖代谢相关酶的研究[J]. 园艺学报,2004,31(6): 719-722. GONG Ronggao,ZHANG Guanglun,L? Xiulan,ZENG Xiuli,
LUO Nan,HU Qiang. Studies on the sucrose-metabolizing en zymes in navel orange fruit from different habitats[J]. Acta Hor ticulturae Sinica,2004,31(6):719-722.
[24] 高彦婷,张芮,李红霞,魏鹏程 . 水分胁迫对葡萄糖分及其蔗糖 代谢酶活性的影响[J]. 干旱区研究,2021,38(6):1713-1721.
GAO Yanting,ZHANG Rui,LI Hongxia,WEI Pengcheng. Ef fect of water stress on sugar accumulation and sucrose metabo lism enzyme activities of greenhouse grape fruit[J]. Arid Zone Research,2021,38(6):1713-1721.
[25] JULIA W,SJEF S,JOHANNES H. Sucrose: metabolite and sig naling molecule[J]. Phytochemistry,2010,71(14):1610-1614.
[26] BERNARD F,PHILIPPE J,MARC C,COLETTE G,OLIVIER H,DOMINIQUE R,DIDIER M. Acid invertase as a serious can didate to control the balance sucrose versus(glucose + fructose) of banana fruit during ripening[J]. Scientia Horticulturae,2011, 129(2):197-206.
[27] YUXING L,BOYANG L,YUXIAO P,CHENLU L,XIUZHI Z,ZHIJUN Z,WEI L,FENGWANG M,CUIYING L. Exoge nous GABA alleviates alkaline stress in malus hupehensis by regulating the accumulation of organic acids[J]. Scientia Horti culturae,2020,261:108982.
[28] 梁靜宜,郭凡,赵科,王鸿飞,许凤 . 外源 γ-氨基丁酸对鲜切南 瓜品质和γ-氨基丁酸代谢的影响[J]. 食品工业科技,2022,43 (19):385-392.
LIANG Jingyi,GUO Fan,ZHAO Ke,WANG Hongfei,XU Feng. Effect of exogenous quality and γ- aminobutyric acid on GABA metabolism in fresh-cut pumpkins[J]. Science and Tech nology of Food Industry,2022,43(19):385-392.
[29] ETIENNE A,G?NARD M,LOBIT P,MBEGUI?- A MB?GUI? D,BUGAUD C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells[J]. Journal of Experimental Botany,2013,64(6):1451-1469.
[30] YANG C,CHEN T,SHEN B R,SUN S X,SONG H Y,CHEN D,XI W P. Citric acid treatment reduces decay and maintains the postharvest quality of peach (Prunus persica L. )fruit[J]. Food Science & Nutrition,2019,7(11):3635-3643.
[31] FAMIANI F,BONGHI C,CHEN Z H,DRINCOVICH M F, FARINELLI D,LARA M V,PROIETTI S,ROSATI A,VIZ ZOTTO G,WALKER R P. Stone fruits:Growth and nitrogen and organic acid metabolism in the fruits and seeds:A review[J]. Frontiers in Plant Science,2020,572601.
[32] FARAJ H,NABIL K. The use of deuterium-labeled gamma-ami nobutyric (d6-GABA) to study uptake, translocation, and metab olism of exogenous GABA in plants[J]. Plant Methods,2020,16 (1):24.
[33] ZHOU L,JINGJIN Y,YAN P,BINGRU H. Metabolic pathways regulated by γ- aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera) [J]. Scien tific Reports,2016,6(1):30338.
[34] MATTHEW G,STEPHEN D T. Linking metabolism to mem brane signaling: the GABA- malate connection[J]. Trends in Plant Science,2016,21(4):295-301. [35] ALEXIS D A,ULRIKE B,RITA F,JINGBO Z,MARIA M C, ANA R. The vacuolar channel vvalmt9 mediates malate and tar trate accumulation in berries ofVitis vinifera[J]. Planta,2013, 238(2):283-291
猜你喜欢
中国食品(2021年17期)2021-09-27
现代畜牧科技(2021年6期)2021-07-16
现代食品(2021年2期)2021-03-11
食品工业(2020年10期)2020-11-02
中国粮油学报(2019年4期)2019-07-12
现代盐化工(2018年1期)2018-03-29
食品安全导刊(2017年24期)2017-09-16
磁共振成像(2015年5期)2015-12-23
饲料博览(2015年2期)2015-04-04
饲料博览(2014年9期)2014-04-05
