
玉米大斑病菌醇脱氢酶基因家族的鉴定和生物信息学分析
2023-06-09 03:28:56张淑红张运峰武秋颖高凤菊李亚子纪景欣范永山
浙江农业学报 2023年5期
张淑红,张运峰,武秋颖,高凤菊,李亚子,纪景欣,许 可,范永山
(唐山师范学院 生命科学系,唐山市农业病原真菌与毒素重点实验室,河北 唐山 063000)
醇脱氢酶(alcohol dehydrogenase,ADH)广泛存在于人和哺乳动物的肝脏、植物组织及微生物细胞中,是生物体内重要的氧化还原催化剂之一,其为具有广泛专一性的含锌金属酶,以NAD+为辅酶,参与糖酵解末端反应[1]。在各种代谢物的生物合成过程中,ADH催化醇和醛或酮之间的相互转化,并且在酒精、烷烃、糖、脂质代谢以及外源性酒精和醛类防御方面都有着极其重要的作用[2-5]。
ADH具有广泛的底物特异性。在人和哺乳动物体内,ADH是乙醇代谢的关键酶,在酒精性肝损伤的诊断、治疗和解酒药物研究中发挥重要作用[6-8]。在植物组织中,ADH是无氧呼吸时能量生成和NAD+再生的关键酶[9]。来源于微生物的ADH结构差异较大,有的为二聚体,有的为四聚体,单体的折叠结构与哺乳动物相似,主要区别是活性位点与锌离子的协调作用不同[10]。酵母ADH是145 ku 的四聚体,每条单体结合1个 NAD+和1个 Zn2+。酵母ADH主要有3种依赖Zn2+的同工酶——厌氧表达的YADH1、细胞质组成成分YADH2 和线粒体中的YADH3,其中YADH1是酵母ADH活性的主要部分[11]。
ADH参与植物病原真菌致病性的研究较少。有报道[12]指出,灰霉菌(Botrytiscinerea)BcADH1、BcADH2和BcADH3等3个ADH基因在病菌侵染寄主早期转录水平急剧提升,参与调控该菌的生长发育、逆境适应和致病过程,是必不可少的致病相关因子。玉米大斑病菌(Setosphaeriaturcica)是威胁玉米生产的一种重要病原真菌,ADH基因在病菌侵染过程中的功能尚未明确。本文在玉米大斑病菌ADH基因(StADH)家族的鉴定和生物信息学分析的基础上,进一步分析它们在病菌侵染过程中的表达规律,为明确ADH基因在致病过程中的功能奠定基础。
1 材料与方法
利用乙醇脱氢酶(ADH)的EC.1.1.1.1检索玉米大斑病菌Et28A菌株基因组的KEGG数据库(https://mycocosm.jgi.doe.gov/Settu1/Settu1.home.html),获得玉米大斑病菌ADH家族成员的基因和蛋白质序列,利用在线分析工具MG2C(http://mg2c.iask.in/mg2c_v2.1)进行染色体定位,利用ProtParam(https://web.expasy.org/protparam)进行理化性质分析,利用WOLF PSORT(https://wolfpsort.hgc.jp)进行亚细胞定位,利用NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和InterPro(http://www.ebi.ac.uk/interpro/)进行结构域分析,利用MEME(https://meme-suite.org/meme)进行保守基序分析。
通过UniProt数据库(https://www.uniprot.org)获得酿酒酵母(Saccharomycescerevisiae)ADH氨基酸序列,利用MEGA7.0软件采用邻接法(neighbor-joining method, NJ)构建系统发育树。基于本课题组已有的玉米大斑病菌接种抗病自交系B73Ht1(R)和感病自交系B73(S)玉米叶片3、5、7、10 d后的转录组数据,以β-肌动蛋白为内参,获得各基因的相对表达量,利用TBtools[13]和GraphPad prism软件(GraphPad Software, San Diego, California USA, www.graphpad.com)进行作图分析。
2 结果与分析
2.1 玉米大斑病菌ADH基因家族的染色体定位
利用玉米大斑病菌基因组的KEGG数据库共获得22个ADH基因家族成员,按照基因定位分别命名为StADH1-StADH22(图1)。基因定位结果表明,22个StADH家族成员分布在12条scaffold上,其中以scaffold2最多,分布有6个基因。而其他395条scaffold上未发现有玉米大斑病菌ADH基因家族成员分布。

图1 玉米大斑病菌ADH基因染色体定位Fig.1 Chromosome locations of ADH genes in Setosphaeria turcica
2.2 玉米大斑病菌ADH基因家族成员的序列及理化性质分析
22个StADH基因长度变化较大,CDS序列全长696~1 485 bp,编码232~495个氨基酸残基,分子量31 538.78~53 590.22,理论等电点5.65~9.22。除了StADH5、StADH15、StADH18和StADH22显碱性外,其他18个ADH基因编码蛋白均为酸性。疏水性平均值显示,StADH6、StADH9、StADH11、StADH15、StADH17及 StADH21呈现疏水性,其他16个基因家族成员编码蛋白均显示亲水性。亚细胞定位结果表明,StADH7、 StADH15、StADH20和StADH21定位于线粒体,StADH4、StADH22定位于过氧化物酶体,其余家族成员均定位于细胞质中。
2.3 玉米大斑病菌ADH蛋白系统发育树及结构分析
利用UniProt数据库(https://www.uniprot.org)获得酿酒酵母(Saccharomycescerevisiae)7个ADH基因家族成员(YADH1-YADH7),与玉米大斑病菌22个ADH基因家族成员(StADH1-StADH22)共同构建系统发育树,利用InterPro和MEME分析其保守结构域和保守基序(图2)。结果表明,玉米大斑病菌ADH家族蛋白分为6类:第Ⅰ类包含StADH2、StADH4、StADH7、StADH10、StADH12-13、StADH19-20等8种,与酵母YADH4亲缘关系较近,其结构中含有0~2个保守基序,其中StADH4和StADH13分别含有乙醇脱氢酶的保守结构域ADH_N和ADH_zinc_N[9,14],StADH2和StADH12含有保守结构域Aldo_ket_red,属于醛酮还原酶家族[15-16],其余6个基因与YADH4具有相同的保守结构域Fe-ADH,属于含铁醇脱氢酶家族[17];第Ⅱ类包括StADH1、StADH5、StADH9、StADH14、StADH16、StADH22等6种,含有2~4个保守基序,仅StADH22的结构域为ADH_zinc_N,其余均为ADH_N;第Ⅲ类为StADH3和StADH8,分别含有4、5个保守基序,均含有ADH_zinc_N结构域序列;第Ⅳ类为StADH21和StADH6,与YADH1、YADH2、YADH3、YADH5亲缘关系较近,其中StADH6与YADH均含有5个保守基序,典型结构域为ADH_N,而StADH21不含Motif3,结构域为ADH_zinc_N;第Ⅴ类为StADH11、StADH17和StADH18,均含有5个保守基序,典型结构域为ADH_zinc_N;第Ⅵ类为StADH15,与YADH6、YADH7亲缘关系较近,都含有5个保守基序,结构域组成也相同,均为ADH_N。
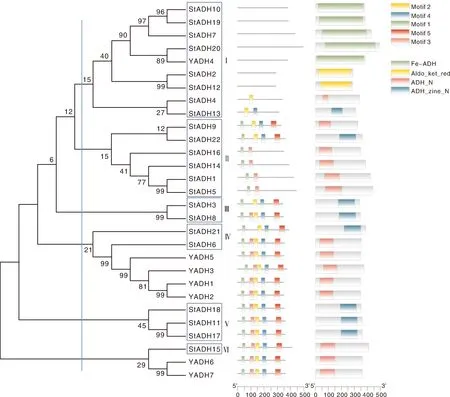
图2 玉米大斑病菌ADH的系统发育树Fig.2 Phylogenetic tree of the ADH of Setosphaeria turcica
2.4 玉米大斑病菌ADH基因在病菌侵染过程中的表达分析
分析玉米大斑病菌ADH基因接种到抗病自交系B73Ht1(R)和感病自交系B73(S)玉米叶片3、5、7、10 d的表达规律,结果如图3所示,StADH1基因平均相对表达量最高,为1.98;其次为StSDH6和StADH12,基因平均相对表达量分别为1.69和1.60;StADH4的相对表达量中等,为1.02;其余18个StADH的相对表达量较低,在0.01~0.67。因此,StADH1、StADH4、StADH6和StADH12在病菌侵染过程中可能发挥着较重要的作用。

图3 玉米大斑病菌ADH基因在接种后不同时间的相对表达量分析Fig.3 Analysis of the relative expression of ADH gene in Setosphaeria turcica at different time after inoculation
分析StADH1、StADH4、StADH6和StADH12在病菌侵染后的表达规律(图4-A、B),发现接种玉米叶片3、5、7、10 d后,StADH6的表达量比较稳定,接种期间差异不显著;StADH1、StADH4和StADH12在接种3、5、7 d 时表达量很低,10 d 时表达量均显著升高(P<0.05),其中StADH1和StADH4差异达到极显著水平(P<0.01)。比较接种抗病玉米(R)和感病玉米(S)10 d时的基因表达差异(图4-C),结果发现,接种感病玉米StADH1基因的表达量显著高于接种抗病玉米(P<0.05),StADH4、StADH6和StADH12则差异不显著。

A,接种抗病自交系(R);B,接种感病自交系(S);C,接种10 d的基因表达差异;*表示差异显著(P<0.05)。A,Inoculated disease-resistant inbred line (R); B,Inoculated susceptible inbred line (S); C,Difference of gene expression after 10 days of inoculation; * Indicating the significant difference(P<0.05).图4 玉米大斑病菌ADH基因家庭成员在病菌侵染后的表达规律Fig.4 Expression of family members of ADH gene in Setosphaeria turcica after infection
以上研究表明,StADH1、StADH4、StADH12在病菌接种10 d时发挥重要作用,是关键StADH基因,它们主要在病害侵染后期起作用,其中StADH1还可能与病菌的致病特异性有关。
2.5 玉米大斑病菌ADH基因家族关键基因的功能分析
NCBI CDD分析结果显示,StADH1、StADH4及StADH6均包含MDR superfamily(依赖锌/中链醇脱氢酶超家族)(图5)。StADH1的典型结构域FDH_like_1为谷胱甘肽依赖性甲醛脱氢酶,由Motif1和Motif3组成(图2),FDH可将甲醛和NAD(P)转化为甲酸和NAD(P)H。StADH4的典型结构域enoyl_reductase_like为类烯酰还原酶,由Motif2组成(图2),催化脂肪酸合成中反式烯酰载体蛋白/辅酶A(ACP/CoA)依赖于NADPH转化为酰基(ACP/CoA)。此活性在热带假丝酵母中被认为是维持线粒体呼吸功能所必需的。StADH6的结构域CAD3为肉桂醇脱氢酶,由Motif1、Motif2、Motif3、Motif4、Motif5组成(图2),其主要功能是依赖NAD(P)(H)催化醇和醛或酮之间的相互转化。StADH12的结构与其他3个StADH不同,在分子中没有发现保守基序,其结构域为AKR_AKR1-5-like,是一类可溶性NAD (P)(H)氧化还原酶,属于AKR_SF superfamily(图5),其主要功能是将醛和酮还原为仲醇。
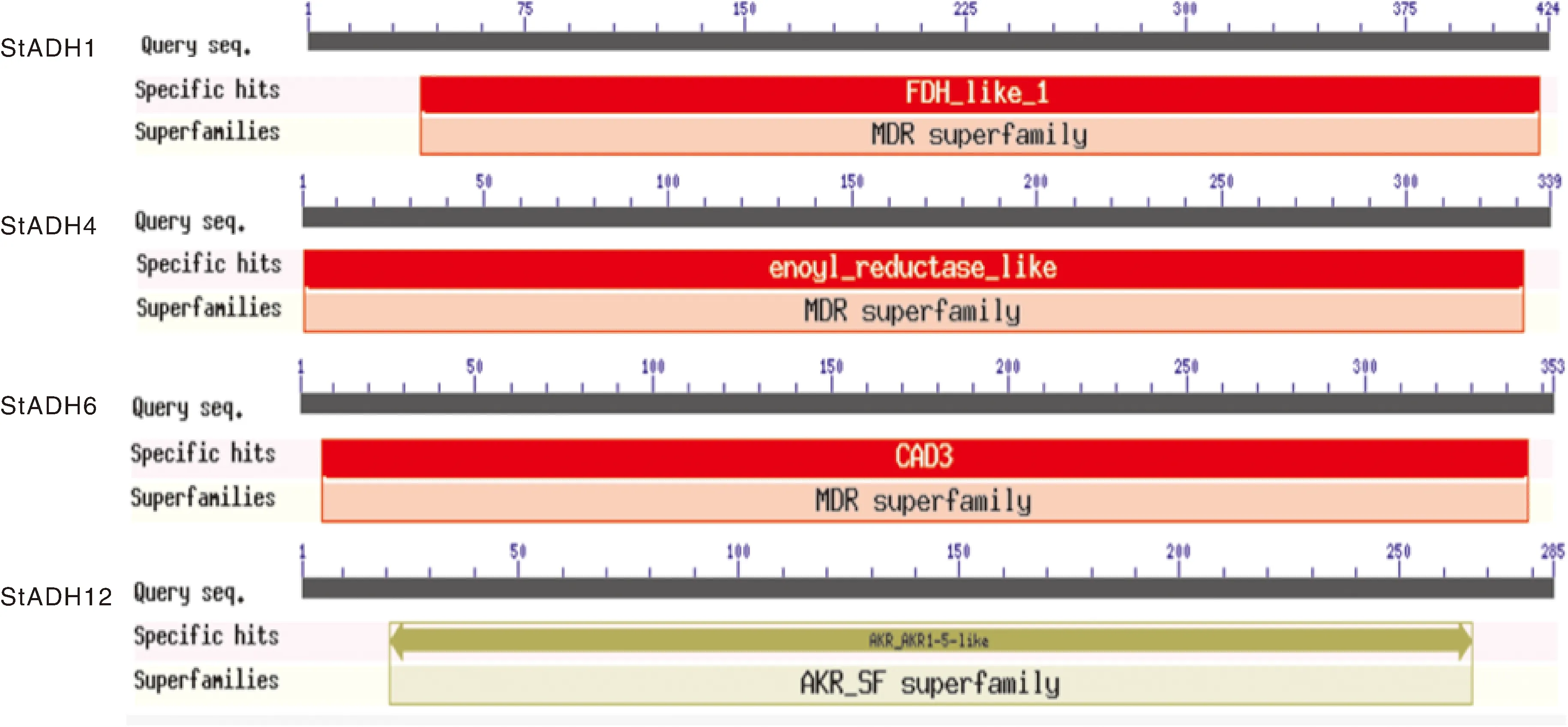
图5 玉米大斑病菌ADH基因家族关键基因的结构域分析Fig.5 Domain analysis of key genes of ADH gene family in Setosphaeria turcica
3 讨论
目前,在微生物、动物、植物细胞中都存在醇脱氢酶,且在微生物中最广泛[18]。醇脱氢酶(ADH)是生物体内重要的醇类代谢酶,具有广泛的基因多态性。酿酒酵母有7个ADH编码基因(YADH1-YADH7)。YADH1是催化乙醛生成乙醇的关键酶[19],将NADH再生为NAD+,当以葡萄糖为发酵底物时,该基因组成型表达,以非葡萄糖如乙醇、甘油为唯一碳源时,YADH1基因表达受抑制。YADH2催化乙醇生成乙醛,高浓度葡萄糖抑制其基因表达,主要参与 NADH 氧化还原平衡,氨基酸代谢和杂醇类的生成[20]。YADH3存在于线粒体基质中,表达受葡萄糖阻遏抑制,主要功能是维持线粒体内氧化还原平衡和参与乙酸-乙醛还原穿梭途径[21]。YADH4在葡萄糖发酵过程中将乙醛降解为乙醇,当锌含量受到限制时,可能会结合铁,为依赖Fe2+的乙醇脱氢酶[22]。YADH5催化乙醇生成乙醛,可在细胞核内起作用[23]。YADH6和YADH7的辅酶都为NADP+。在玉米大斑病菌基因组内发现的22个ADH家族成员中,与酵母聚到相同类群的有11个基因,其中StADH4、StADH12等8个基因与YADH4聚到同一类群,StADH6和StADH21与YADH1、YADH2、YADH3、YADH5聚到同一类群,StADH15与YADH6、YADH7聚到同一类群。另外11个玉米大斑病菌ADH家族成员又分为3类,其中StADH1等6个基因为同一类群,StADH3、StADH8及StADH11、StADH17、StADH18分别聚为同一类群。以上研究表明,玉米大斑病菌存在与酵母不同的ADH家族成员。
在玉米大斑病菌ADH家族成员中,仅有StADH1、StADH4、StADH6和StADH12在病菌侵染过程中起主要作用。其中StADH6在侵染过程中表达量稳定,可能为组成型表达;StADH1、StADH4、StADH12在接种10 d时表达量均显著增高,表明它们可能在糖酵解无氧氧化过程中发挥着关键作用,对于病菌侵入寄主体内后期的生存非常重要。StADH4和StADH12在接种抗病品种和感病品种间表达量差异不显著,因此与寄主专化性无关,而StADH1接种感病品种的表达量显著高于接种抗病品种,说明该基因是玉米大斑病菌的重要致病基因,由于其含有FDH_like_1结构域,可通过催化产生甲醛或甲酸破坏寄主细胞,在病菌的成功侵染、定殖和病害发生过程中都可发挥非常重要的作用。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
幼儿100(2023年18期)2023-05-29 08:34:36
课堂内外·小学版(低年级)(2022年4期)2022-06-24 09:47:10
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
孩子(2019年8期)2019-08-26 05:43:32
心肺血管病杂志(2018年11期)2018-12-18 01:51:40
中国医药生物技术(2015年4期)2015-12-26 08:26:36
聪明泉·少儿版(2014年2期)2014-09-22 09:54:50
河南医学研究(2014年3期)2014-02-27 14:51:59
