
蜘蛛的种类和捕食策略对假眼小绿叶蝉捕食效率的影响*
2023-06-01 07:20:30包文杰刘胜杰
中山大学学报(自然科学版)(中英文) 2023年3期
包文杰,刘胜杰
1.中国科学院西双版纳热带植物园,云南 勐腊 666303
2.中国科学院大学,北京 100049
3.中山大学生态学院,广东 广州 510275
在农业生态系统中,害虫的生物防治是一项重要的生态系统服务措施(Naranjo et al.,2015)。但是以往多数关于害虫防治的研究总是将天敌种类的捕食效应简单地归为同一个营养级上的作用强度(Hooper et al.,2005),认为不同捕食性天敌对害虫的抑制作用是相同的,且可以相互取代。但这与生态系统中的实际情况不相符,低估了捕食者多样性对害虫的抑制作用(Griffen,2006;Griffin et al.,2013)。相关研究发现,天敌的捕食策略会显著影响其对猎物的捕获效率(Schmitz,2008;Schmitz,2009;Liu et al.,2015a)。例如,游猎型蜘蛛(actively hunting spider)能够通过直接捕食而显著地减少蚱蜢数量,而结网等待型蜘蛛(sit-andwait spider)并没有直接降低蚱蜢的种群数量,只是间接影响了蚱蜢的取食习惯和活动频率(Schmitz,2008)。我们前期在土壤生态系统中的研究也发现了类似的结论,游猎型蜘蛛能够显著减少弹尾目的密度,从而对凋落物分解产生级联效应,而结网等待型蜘蛛对弹尾目密度没有显著的影响(Liu et al.,2015b)。游猎型和结网等待型捕食者总是共存于农业生态系统中,但是关于这两类捕食者对农业害虫的生物防控效果还不是很清楚。
与此同时,以往的研究经常忽视捕食者之间的相互竞争或协作对害虫种群动态的调节作用(Cardinale et al.,2003;Griffen,2006;Griffin et al.,2013)。Aquilino et al.(2005)的研究发现,与单一种类天敌相比较,两种瓢虫和一种草蛉组合在一起能够显著抑制茶蚜的种群增长,因为这3 种天敌之间存在着生态位的分化,没有强烈的种间或种内竞争。而Finke et al.(2005)发现,较高的天敌多样性反而增加了蜡蝉的种群增长,因为蜡蝉的天敌之间存在很强的相互取食和竞争。较高的天敌多样性和密度,增加了天敌之间的竞争和相互取食,从而有利于蜡蝉的种群增长。廖先骏等(2016)的研究也发现了类似的结论,随着草蛉幼虫密度的增加,种内竞争强度也随之增加,从而导致它们对茶蚜种群的抑制作用降低。由此可见,深入了解捕食性天敌的种内种间竞争,才能更好地预测天敌多样性变化对害虫生物防控效果的影响。
茶园中植物物种组成单一,生态系统结构较为简单,容易造成虫害的大规模爆发。系统性地探究天敌和害虫相互作用机制能够为茶园生态系统的害虫防治提供理论基础(Hazarika et al.,2009)。虫害对我国茶叶年产量造成损失约10%,有些茶园损失严重甚至达到15% ~ 50%(苏红飞等,2019)。假眼小绿叶蝉(Empoasca vitisGothe)是我国茶园主要的害虫之一,在全国茶区均有发生。蜘蛛是茶园生态系统中最主要的捕食性天敌,约占茶园捕食性天敌生物量的80% ~90%,在控制害虫的种群动态方面具有重要作用(戴轩,1992;陈银方,2016)。茶园蜘蛛也是害虫假眼小绿叶蝉的主要天敌,已确认捕食假眼小绿叶蝉的蜘蛛共有80种(戴轩,1999)。然而,关于不同种类和捕食策略的蜘蛛对假眼小绿叶蝉捕获效率的差异目前还没有明确的答案,因此本研究通过野外调查和室内饲养试验相结合的方法,拟回答以下科学问题:(1)茶园生态系统中不同月份蜘蛛的群落动态变化?(2)不同种类的蜘蛛对假眼小绿叶蝉捕获效率是否有差异?(3)不同捕食策略的蜘蛛对假眼小绿叶蝉捕获效率是否有差异?
1 材料与方法
1.1 研究地点自然概况
本调查选取了我国重要的茶产地——云南省普洱市一处具有代表性的有机茶园,大尖山茶场(22°49′N,100°57′E),隶属于普洱市龙生茶叶集团。普洱市位于云南省西南部,海拔1 340 m,气候为亚热带山地季风气候,年均温为15.3 ~20.2 ℃,年均降雨量1 600 mm,具有低纬、多雨、湿润、静风的特点。大尖山茶场海拔约1 700 m,茶园面积为43.3 hm2。茶园周围被较大面积的森林围绕,人工除草,使用少量生物农药。冬季封茶园进行修剪。农事管理较好,采摘频繁。
1.2 蜘蛛群落野外调查取样方法
滇南茶园通常11 月下旬进入停采、封园期,翌年1 月下旬或2 月上旬进入开采期,故茶园中蜘蛛群落调查期选定为2~11 月,调查时间为2012 年的8 月、11 月和2013 年的4 月、6 月,这4 个时 间段是蜘蛛群落变化的转折时期。在大尖山茶场设置了3 个100 m × 100 m 的样方。在样方内采用击打法(beating-tray method)调查茶园中蜘蛛群落结构。选取相邻的3棵茶树作为一个取样点,一个样地共有20 个取样点。取样时用力击打每棵茶树约30 下,底部用直径为 50 cm 的扫网接住茶树上掉落的蜘蛛。每两个取样点之间至少间隔20 m。击打所收集的蜘蛛样品放入装有酒精的自封袋中,带回室内鉴定种类和统计数量。由于蜘蛛分类的复杂性和受分类知识所限,因此仅根据蜘蛛的外部形态特征将蜘蛛分为形态种(morphospecies),如眼睛的大小、数目和排列方式,步足,纺器,螯肢等等。所有蜘蛛的鉴定都由同一个人通过相同的方式完成,最后再根据Murphy et al.(2000)的介绍将收集到的蜘蛛分为两个不同的功能群:结网等待型蜘蛛和游猎型蜘蛛。
1.3 蜘蛛对假眼小绿叶蝉日取食量的室内饲养试验
1.3.1 供试蜘蛛的采集和选取本实验所选取的蜘蛛均来自云南省普洱市有机茶园(22°02′~24°50′N,99°09′~102°19′E),根据前期的研究和野外调查结果(Liu et al.,2015a),本实验共选取了8 种常见蜘蛛,分别为蓝翠蛛(Siler cupreus)、白斑猎蛛(Evarcha albaria)、斜纹猫蛛(Oxyopes sertatus)、八斑球腹蛛(Theridion octomaculatum)、温室希蛛(Achaearanea tepidariorum)、机敏异漏斗蛛(Agele‐na difficilis)、八斑鞘腹蛛(Coleosma octomacula‐tum)、白 缘 盖 蛛(Neriene albolimbata)。根 据Schmitz(2008)所提到的蜘蛛捕食策略,我们将这8种蜘蛛分为游猎型蜘蛛和结网等待型蜘蛛两大类。其分类标准为:游猎捕获猎物的蜘蛛为游猎型蜘蛛;结网捕获猎物的蜘蛛为结网等待型蜘蛛。其中蓝翠蛛、白斑猎蛛、斜纹猫蛛属于游猎型蜘蛛,其他5种蜘蛛则归为结网等待型蜘蛛。
1.3.2 室内茶园微环境和蜘蛛对假眼小绿叶蝉捕获效率我们利用透明塑料整理箱(长26 cm,宽18 cm,高15 cm)模拟茶园生境建立微生态系统,即在整理箱的底部中放入约1 cm 厚的茶园土壤,然后放入新鲜茶树枝,并保持微生境的湿度和通风。首先将成熟但还未产卵的雌性蜘蛛分别放入整理箱中(1 只/盒)适应环境或结网,24 h 后再开始试验。每个蜘蛛个体喂食10 只假眼小绿叶蝉,24 h后检查整理箱中剩余的假眼小绿叶蝉数量,从而测定不同种类和捕食策略的蜘蛛对假眼小绿叶蝉的捕获效率。选取10 只假眼小绿叶蝉是根据前期的调查结果模拟了野外假眼小绿叶蝉种群爆发时的虫口密度(Liu et al.,2015a)。每个蜘蛛个体重复测量3次其对假眼小绿叶蝉的日捕食量,计算平均值。每种蜘蛛的个体有10 次重复,最终结果为10次重复的平均值。
1.3.3 蜘蛛的种内和种间作用对假眼小绿叶蝉捕获效率的影响为了验证蜘蛛的种内和种间相互作用对假眼小绿叶蝉捕获效率的影响,我们选取茶园优势蜘蛛种类蓝翠蛛(游猎型)和温室希蛛(结网等待型)进行这部分的实验。共有3个实验处理:2 只蓝翠蛛、2 只温室希蛛、混合蜘蛛(1 只蓝翠蛛+1 只温室希蛛)。因为第一部分实验整理箱中放置了1 只蜘蛛,这部分实验选择放置了2 只蜘蛛,转换后蜘蛛的密度为43 只蜘蛛/m2,根据前期调查结果,该蜘蛛密度会产生较为明显的种内种间相互作用(Liu et al.,2015a)。具体的实验操作过程为:在整理箱中放入2 个蜘蛛个体,20 只假眼小绿叶蝉,24 h 后检查整理箱中剩余的假眼小绿叶蝉数量,实验重复数为8,最终结果为8 次重复的平均值。
1.4 数据分析
数据的分析和可视化通过R v3.6.3 实现。采用单因素方差分析(one-way ANOVA)检验蜘蛛的捕食策略和种类对假眼小绿叶蝉捕食量的差异。采用非参数Kruskal-Wallis 方法分析蜘蛛的种内和种间相互作用对假眼小绿叶蝉捕食量的影响。种内相互作用的检验方式为,2 只蜘蛛处理对假眼小绿叶蝉的捕获量与1 只蜘蛛捕获量的2 倍进行非参数比较。种间相互作用检验方式类似,混合蜘蛛处理对假眼小绿叶蝉的取食量与各自蜘蛛种类单独取食量相加进行非参数比较。
2 结 果
2.1 蜘蛛群落调查
茶园内蜘蛛群落动态变化的结果表明:游猎型和结网等待型蜘蛛的物种数和多度在4个调查时间点的变化趋势基本相同,在6 月和11 月都呈现上升的趋势。游猎型蜘蛛物种数目的最大值出现在11 月,为8.3 种/样地;结网等待型蜘蛛物种数目的最大值出现在6 月份,为13.7 种/样地(图1A,C)。6 月份游猎型和结网等待型蜘蛛的多度都增加,但不是最高点,分别为51.3 头/样地和145.0头/样地。游猎型和结网等待型蜘蛛多度的最大值出现在11 月份,分别为70.3 头/样地和238.7 头/样地(图1B,D)。蜘蛛群落的总体参数在4 个月份中的变化趋势和结网等待型蜘蛛相似(图1E,F)。

图1 大尖山茶场不同月份蜘蛛物种数(A、C、E)和多度(B、D、F)的变化趋势Fig.1 The trend of species number (A, C, E) and abundance (B, D, F) of spiders in different months in Dajian Mountain Tea Plantation
不同月份茶场中蜘蛛的相对多度结果表明:结网等待型蜘蛛在大尖山茶场中占据优势地位,全年中结网等待型蜘蛛的多度均显著高于游猎型蜘蛛。4 月、6 月、8 月和11 月结网等待型蜘蛛的相对多度分别为68.66%、73.85%、79.12% 和77.24%。8月和11月结网等待型蜘蛛的比例明显高于4月,增加了约10%(图2)。

图2 不同月份结网等待型和游猎型蜘蛛相对多度比例图Fig.2 Relative abundance ratio diagram of sit-and-wait spi‐der and actively hunting spider in different months
2.2 室内饲养试验
通过微生态系统测定不同种类和捕食策略蜘蛛的捕食效率结果显示:游猎型蜘蛛对假眼小绿叶蝉的日取食量要显著高于结网等待型蜘蛛(P<0.001)(图3)。同样,不同种类的蜘蛛对假眼小绿叶蝉的日取食量也有显著的差异,白斑猎蛛和斜纹猫蛛捕食量最高,平均约7.3 头/d,显著高于其他蜘蛛种类(P< 0.05)(图4)。但是在两种捕食策略内,不同的蜘蛛种类之间对假眼小绿叶蝉的捕获效率没有显著差别(图4)。两只结网等待型蜘蛛处理对假眼小绿叶蝉的日取食量要显著高于单个蜘蛛个体的两倍(Kruskal-Wallis,P< 0.05)(图5),该结果表明两只温室希蛛之间存在着较明显的种内协作。混放蜘蛛处理(游猎型+结网等待型蜘蛛)也能显著促进各自蜘蛛个体对假眼小绿叶蝉的捕食(Kruskal-Wallis,P< 0.05)(图5),表明蓝翠蛛和温室希蛛之间存在着种间协作,而蓝翠蛛种内不存在明显的相互作用(Kruskal-Wallis,P>0.05)(图5)。
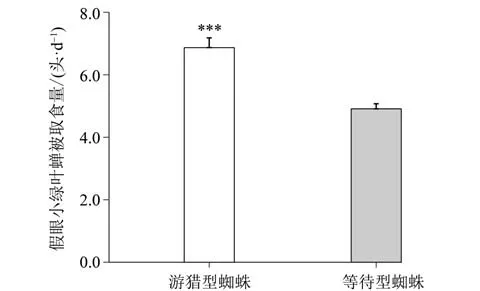
图3 不同捕食策略的蜘蛛对假眼小绿叶蝉捕获效率的比较Fig.3 Comparison of the predation efficiency of spiders with different foraging strategies to the green leafhopper

图4 不同种类的蜘蛛对假眼小绿叶蝉捕获效率的比较Fig.4 Comparison of the predation efficiency of different spider species to the green leafhopper

图5 蜘蛛的种内和种间作用对假眼小绿叶蝉捕获效率的比较Fig.5 Comparison of intraspecific and interspecific effects of spiders on their predation efficiency to the green leafhopper
3 讨 论
云南地区茶园一年中假眼小绿叶蝉的发生具有2个高峰期,第1次为5~6月,第2次为10~11月(Liu et al.,2015a;苏红飞等,2019),在我们所调查的4 个时间点中,6 月和11 月蜘蛛的物种数和多度相较于4 月和8 月都有一个明显的增加,表明随着茶园中假眼小绿叶蝉种群增长,蜘蛛的物种数和多度也会相应地增加。蜘蛛、假眼小绿叶蝉和茶树在茶园中组成了一个较为单一的食物链,负反馈是自然界中一项重要的自我调节机制,能够稳定食物网的动态(Mougi et al.,2016)。相关研究结果表明,较高的捕食者多样性能够显著抑制害虫种群数量的增长和爆发,起到很好的生物防控 效 果(Cardinale et al.,2003;Aquilino et al.,2005;Finke et al.,2005)。4月和8月蜘蛛多度下降可能是由于茶园进入开采期,假眼小绿叶蝉把卵产在嫩叶中,茶农通过采摘鲜叶能够带走大部分虫卵,由于假眼小绿叶蝉种群的减少,相应地也会导致蜘蛛多度的减少。到11 月份,由于进入停采、封园期,不受采摘的影响,假眼小绿叶蝉的种群数量迅速增长,蜘蛛多度又呈现一个快速上升的趋势。
捕食者的很多特性都会影响它们对猎物的捕获效率,捕食策略是其中一个重要的方面(Abrams,2008;Nakazawa et al.,2013;Miller et al.,2014)。Ross et al.(2015)通过构建捕食者和猎物的行为生态学模型发现:当捕食者的移动需要消耗大量的能量时,此类捕食者倾向于采取坐等捕食的策略;当捕食者速度过快而容易引起猎物的警觉时,此类捕食者更多采取的是埋伏追击的捕食策略。Huey et al.(1981)研究发现沙漠中游猎觅食的蜥蜴比静坐等待的蜥蜴吃掉更多聚集的白蚁或不活跃的蝎子。同样,Miller et al.(2014)发现与游猎型蜘蛛相比,织网等待的蜘蛛捕食范围较小,捕食的蚱蜢数量也更少。捕食者能够通过致死效应(直接捕食猎物)或特征效应(引起猎物行为的变化以避免被捕食)影响猎物种群(Cresswell et al.,2010)。先前的研究发现游猎型蜘蛛对猎物种群有强烈的致死效应,很少引起猎物行为的改变;相反,结网型蜘蛛通常引起猎物栖息地的转变而不 存 在 显 著 的 致 死 效 应(Schmitz et al.,2001;Schmitz,2008)。这与我们的研究结果相一致,游猎型蜘蛛具备更加积极主动的捕食策略,而结网型蜘蛛采取的是一种“守株待兔”的捕食策略,所以游猎型蜘蛛对假眼小绿叶蝉的捕获效率要显著高于结网等待型蜘蛛。我们还发现不同种类的蜘蛛对假眼小绿叶蝉的日捕食量也有显著不同,但在游猎型蜘蛛和结网等待型蜘蛛内,不同种类蜘蛛对假眼小绿叶蝉的捕获量并没有显著差异,这进一步说明蜘蛛的捕食策略确实是影响其对猎物捕获效率的一个重要功能特征。
早期关于捕食者-猎物相互作用的大部分研究和理论都是基于只有单一捕食者而言(Soluk,1993;Cardinale et al.,2003)。然而,在自然界中捕食者并不是单独存在的,总是有不同种类和不同个体的捕食者共存于同一个生态系统中。当同一种猎物面对多个或者多种捕食者时,多个捕食者对同一猎物的捕获量并不等于单一捕食者捕获量的简单叠加。当捕食者之间存在协作效应时,多个捕食者对猎物的捕获量将高于每个捕食者捕获量的叠加之和;当捕食者之间存在着相互竞争或相互捕食时,多个捕食者对猎物的捕获量将低于每个捕食者捕获量的叠加之和;若捕食者之间不发生相互作用,那么多个捕食者的捕获量则等于每个捕食者捕获量的叠加之和(Sih et al.,1998;Vance-Chalcraft et al.,2004;Vance-Chalcraft et al.,2005;Griffen,2006)。我们的研究发现,两只温室希蛛个体之间存在着较为明显的种内协作和互惠关系。多个结网等待型蜘蛛个体在茶树周围同时织网能够缩小假眼小绿叶蝉的活动范围,从而更有效地对其进行捕食,增加各自对假眼小绿叶蝉的捕获量。由于游猎型蜘蛛不断移动对假眼小绿叶蝉进行捕食,会将其赶往结网等待型蜘蛛布置的“陷阱”中,因此蓝翠蛛和温室希蛛的种间相互作用也会提高各自对假眼小绿叶蝉的捕获效率。但是由于游猎型蜘蛛习惯于在更加广阔的范围内寻找更多的猎物,个体之间相遇的概率较小,种内协作和竞争都不明显,因此蓝翠蛛的种内相互作用并未影响各自对假眼小绿叶蝉的捕获效率。
4 结 论
茶园生态系统中,蜘蛛群落变化与假眼小绿叶蝉的种群动态变化相关,这表明蜘蛛物种数和多度的增加对茶园主要害虫假眼小绿叶蝉种群具有抑制作用。室内饲养试验结果表明,蜘蛛的种类和捕食策略会显著影响其对假眼小绿叶蝉的捕获效率,且温室希蛛种内和蓝翠蛛与温室希蛛种间存在互惠协作关系,能够提高各自对于假眼小绿叶蝉的捕食量。因此,深入了解捕食性天敌的特性及其种内和种间相互作用能够更好地预测茶园生态系统中害虫生物防治的效果。然而,本文中关于蜘蛛捕食效率的试验只是室内研究工作的初步结果,在实际应用中捕食者的多样性和相互作用往往比室内茶园微环境更加复杂,因此今后的研究需要增加更多的蜘蛛种类以及进一步到田间进行试验来验证该结论。
猜你喜欢
领导文萃(2023年13期)2023-07-10 12:00:26
公民与法治(2022年1期)2022-07-26 05:58:06
云南大学学报(自然科学版)(2021年1期)2021-02-05 08:04:04
太原师范学院学报(自然科学版)(2018年2期)2018-08-17 12:21:50
教学考试(高考化学)(2018年3期)2018-08-07 02:05:26
东华大学学报(自然科学版)(2018年1期)2018-06-29 03:35:24
莫愁(2017年22期)2017-08-28 09:42:26
中外文摘(2016年13期)2016-08-29 08:53:27
蚕桑茶叶通讯(2015年1期)2015-02-22 15:13:21
安徽文学(2014年5期)2014-05-30 05:49:59
