
日本沼虾对6种蛋白质原料中蛋白质、氨基酸及磷的离体消化率研究
2023-05-29 03:01:36孙淼张玲葛亚平陈伟亮刘波刘文斌李向飞
南京农业大学学报 2023年3期
孙淼,张玲,葛亚平,陈伟亮,刘波,刘文斌,李向飞*
(1.南京农业大学动物科技学院/江苏省水产动物营养重点实验室,江苏 南京 210095;2.中国水产科学研究院淡水渔业研究中心,江苏 无锡 214081)
鱼粉是水产动物的优质蛋白来源,富含多种必需氨基酸、脂肪酸、矿物质和维生素,具有氨基酸平衡性好、适口性好、抗营养因子少及含有未知的生长因子等优点[1]。随着海洋渔业资源的衰减,鱼粉的供求关系日趋紧张,价格不断上涨,且鱼粉中的磷含量较高,水产动物对其利用率相对较低以及鱼粉在加工过程中产生的废水和废气对环境造成不利影响等[2],种种原因限制了它在水产饲料中的应用。作为另一种动物性蛋白源——血粉,其粗蛋白含量较高(达80%以上)且磷含量低,富含氨基酸(尤其赖氨酸含量较高),但也存在着适口性差、可消化性差、氨基酸组成不平衡等缺陷。江星[3]研究发现,中华绒螯蟹(Eriocheirsinensis)对血粉中蛋白质和能量的表观消化率均较高,表明血粉是蟹的优质动物蛋白源;而Abery等[4]的报道却指出血粉的添加量受鱼种影响而差异较大,且大多数鱼种对血粉的耐受量不超过20%。植物蛋白源中,豆粕粗蛋白含量较高,必需氨基酸组成合理,抗营养因子易消除,且适口性和消化吸收率均较高,是甲壳动物优质植物蛋白来源。此外,棉粕、菜粕和花生粕也具有蛋白含量高的优点,但由于抗营养因子的存在和黄曲霉毒素污染等问题,其在水产饲料中的应用尚无豆粕广泛[5]。白燕等[6]研究了鲫鱼肠道对4种蛋白质饲料的体外消化率,结果发现以花生粕消化率和氨基酸生成效率最高,类似的结果在凡纳滨对虾[7]中也有报道;此外,在对吉富罗非鱼[8]的研究中发现,虽然鱼粉的表观消化率最高,但其也能高效利用豆粕和菜粕中的粗蛋白。由此可见,不同物种对不同蛋白源的利用具有明显差异。
消化率是评价饲料营养价值的重要指标之一[9],研究方法主要有体内和体外消化法2种。前者又分为直接法(收粪法)和间接法(指示剂法),然而这2种方法都需要收集粪便,不可避免地存在粪便中营养成分溶失的弊端;后者是在试管或锥形瓶中模拟动物体内消化的生理条件,以动物消化道酶提取液作为消化酶,对饲料物质进行消化水解,再通过测定消化后饲料物质及消化液中的相关成分来衡量某种营养成分消化率的研究方法。与体内消化率法相比,其具有工作量小、可批量试验、大幅缩短试验周期等优点[10]。由于消化反应的环境条件及消化酶种类等的差异,离体消化率无法完全真实反映体内消化的进程,但仍具有较高准确度及参考价值。例如,Fox等[11]比较研究了凡纳滨对虾对单一蛋白原料的体外消化率和体内消化率,发现二者具有很高的相关性(R2=0.95)。而叶元土[12]也认为在求得二者的相关系数后,就可以利用离体消化率结果推算在体消化率。以上研究表明,离体消化率在水产动物优质蛋白源筛选方面具有重要作用。
日本沼虾(Macrobrachiumnipponense)俗称青虾,是我国传统淡水养殖的经济虾种之一。与其他虾类相比,其个体较小,养殖产量相对偏低,营养需求与原料消化率等相关基础数据较为缺乏,严重制约了其配合饲料的研发[13]。目前,张南南[14]比较了青虾对鱼粉和4种常用植物蛋白源中的干物质和粗蛋白离体消化率,但氨基酸及磷的消化率相关研究尚为空白。基于此,本试验探究了日本沼虾对6种蛋白原料中蛋白质、氨基酸及磷的离体消化率,以期为日本沼虾高效配合饲料研发提供技术指导。
1 材料与方法
1.1 试验虾及饲料原料
试验虾为杂交日本沼虾太湖1号,运回后暂养于室内的循环系统中,饥饿24 h以排空消化道内容物。
试验原料包括进口鱼粉、血粉、豆粕、菜粕、花生粕和棉粕等6种常规蛋白质原料,均购自江苏海普瑞饲料有限公司,产地分别为秘鲁以及中国的山东、黑龙江、四川、河北、新疆。原料经粉碎后过178 μm孔径筛,于-20 ℃冰箱保存。饲料原料的概略养分含量见表1。
1.2 粗酶液的制备
粗酶液的提取参照叶元土等[15]的方法。选取约2 g的健康个体,快速剖取出虾的胃、肝胰腺和肠道。用超纯水清洗去除组织内容物后,吸干表面水分。然后,分别称取肝胰腺和胃肠道质量并分别加入其质量9倍体积的磷酸盐缓冲液(PBS,0.2 mol·L-1,pH7.4),用玻璃匀浆器在冰浴中匀浆。最后,匀浆液在4 ℃、5 000 r·min-1下离心10 min,上清液即为粗酶提取液,于-80 ℃冰箱中冷冻保存备用。

表1 饲料原料的概略养分含量Table 1 The proximate composition of feed ingredients %
1.3 干物质、粗蛋白及总磷消化率的测定
称量过180 μm 筛的饲料原料样品各0.50 g,放入100 mL具塞锥形瓶中,加入0.2 mol·L-1、pH7.4的PBS 47.5 mL,各消化道部位的粗酶提取液2.5 mL。每组原料酶解时设3个重复,并以消化道各部位的煮沸粗酶提取液为对照组。参照王子淑[16]的方法加入双抗(青霉素和硫酸链霉素)各150 IU·mL-1,置于30 ℃恒温水浴振荡器中反应8 h,振荡频率为50次·min-1,间隔4 h补充1次,以防止外界微生物的干扰。
酶解反应结束,反应残渣用定量滤纸过滤,用超纯水冲洗3~4次后,在105 ℃下烘至恒重并称重。采用凯氏定氮法测定消化后残渣的粗蛋白含量,残渣中总磷含量参照《分光光度法:GB/T 6437—2002》测定。饲料原料中干物质、粗蛋白及总磷体外消化率计算公式如下:
干物质消化率(%)=(饲料样品初重-残渣样品质量)/饲料样品初重×100;
粗蛋白消化率(%)=(饲料样品初重×样品中粗蛋白的含量-残渣样品质量×滤渣样品中粗蛋白的含量)/(饲料样品初重×样品中粗蛋白的含量)×100;
总磷消化率(%)=(饲料样品初重×样品中总磷的含量-残渣样品质量×滤渣样品中总磷的含量)/(饲料样品初重×样品中总磷的含量)×100。
1.4 氨基酸生成量的测定
采用叶元土[17]的茚三酮法测定酶解液中的氨基酸含量。按照前述方法对饲料进行酶解,分别于1、2、4、6、8 h取各组的水解液0.2 mL,加入等体积的100 g·L-1三氯乙酸沉淀蛋白后,于6 000 r·min-1离心20 min。取上清液0.2 mL,加入0.2 mol·L-1的醋酸盐缓冲液(pH5.5)1.8 mL,再加入20 g·L-1茚三酮溶液2 mL显色。置于沸水浴20 min后冷却至室温。加入50%乙醇7 mL,以不加样品为空白对照组,用分光光度计在570 nm处测定吸光值(A570)。用亮氨酸标准溶液制作标准曲线,以此来测定酶解液中氨基酸总量。
1.5 酶解液中无机磷含量的测定
采用钼黄比色法测定酶解液中无机磷含量。分别取1、2、4、6和8 h酶解液2 mL于50 mL容量瓶中,加入钒钼酸铵显色剂10 mL,加水稀释至刻度,摇匀,放置10 min后置于10 mm比色皿,用分光光度计测得试样酶解液的吸光值A400,再根据磷标准曲线查得酶解液的含磷量。其中,血粉、菜粕、棉粕和花生粕因在显色反应过程中易产生沉淀而出现浑浊现象而影响样品的吸光值,需在4 000 r·min-1条件下离心15 min后再测定吸光值。
1.6 数据统计与分析

2 结果与分析
2.1 日本沼虾肝胰腺和胃肠道粗酶液对原料中干物质、粗蛋白及总磷离体消化率的影响
如表2所示,日本沼虾肝胰腺粗酶提取液对豆粕中干物质的消化率最高,显著高于血粉、菜粕和花生粕(P<0.05),但与棉粕和鱼粉间无显著差异(P>0.05)。棉粕中粗蛋白消化率最高,其与豆粕间无显著差异(P>0.05),但显著高于其他原料(P<0.05)。此外,花生粕中总磷消化率最高,其与棉粕和血粉间的差异不显著(P>0.05),但显著高于其他原料(P<0.05)。
如表3所示,日本沼虾胃肠道粗酶提取液对菜粕中的干物质消化率最高,其显著高于棉粕和血粉(P<0.05),但与豆粕、花生粕与鱼粉间无显著差异(P>0.05)。粗蛋白消化率和总磷的消化率均以棉粕最高,其中,棉粕粗蛋白消化率显著高于鱼粉、血粉和花生粕(P<0.05),而与其他2种原料间无显著差异(P>0.05);总磷消化率则显著高于其他各组原料(P<0.05)。

表2 日本沼虾肝胰腺粗酶提取液对原料中干物质、粗蛋白及总磷离体消化率的影响Table 2 Effect of the hepatopancreas enzyme extract of Macrobrachium nipponense on the in vitrodigestibility of dry matter,crude protein and total phosphorus in feed ingredients %

表3 日本沼虾胃肠道粗酶提取液对原料中干物质、粗蛋白及总磷离体消化率的影响Table 3 Effect of the gastrointestinal tract enzyme extract of M. nipponense on the in vitro digestibility ofdry matter,crude protein and total phosphorus in feed ingredients %
2.2 日本沼虾肝胰腺和胃肠道粗酶液对6种饲料氨基酸生成量的影响
由图1可知:原料类别、酶解时间及其交互作用均极显著影响日本沼虾消化道粗酶提取液酶解原料蛋白生成的氨基酸总量(P<0.001)。其中,氨基酸的生成量随消化道粗酶提取液酶解时间的延长而显著升高(P<0.05)。在肝胰腺酶解过程中(图1-A),鱼粉生成的氨基酸总量最高,且显著高于其他原料(P<0.05);4种植物蛋白源中以棉粕生成的氨基酸生成量最高。胃肠道酶解生成的氨基酸总量(图1-B)以血粉最高,其显著高于其他原料(P<0.05)。4种植物蛋白源中仍以棉粕的氨基酸生成量最高,且显著高于其他植物原料(P<0.05)。
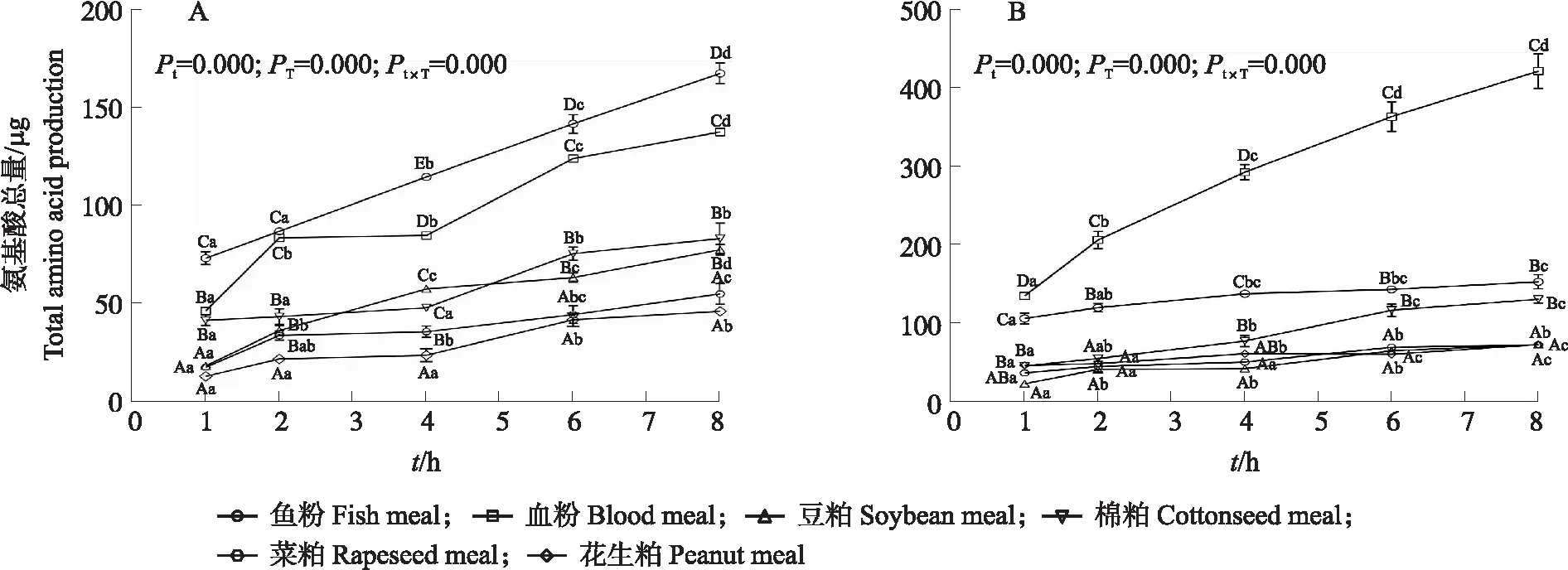
图1 日本沼虾肝胰腺(A)和胃肠道(B)粗酶提取液酶解6种饲料蛋白生成的氨基酸总量变化Fig.1 Total amino acid production of six feed ingredients by the hepatopancreas(A)and gastrointestinal tract(B)enzyme extract of M. nipponensePt:酶解时间P值;PT:原料种类P值;Pt×T:交互作用P值。小写字母不同表示同一饲料不同时间点间差异显著(P<0.05),大写字母不同表示同一时间点不同饲料间差异显著(P<0.05)。下同。Pt:P-value for enzymatic time;PT:P-value for feed type;Pt×T:P-value for interaction effect. Different lower-case letters indicate significant differences(P<0.05)at different time points within each feed,whereas different capital letters indicate significant differences(P<0.05)at the same time point among different feed. The same below.
2.3 6种蛋白饲料在日本沼虾肝胰腺和胃肠道粗酶液中无机磷含量的变化
由图2-A可知:原料种类、酶解时间及其交互作用对日本沼虾肝胰腺粗酶液中无机磷含量均无显著影响(P>0.05)。
由图2-B可知:原料类别、酶解时间及其交互作用均极显著影响胃肠道酶解液中无机磷含量(P<0.01)。随着酶解时间的延长,鱼粉和血粉组胃肠道酶解液中磷含量均在6 h达到峰值,随后血粉组显著下降(P<0.05)。花生粕组变化趋势与之相似,区别在于其峰值在4 h处出现。豆粕和菜粕均在2 h达到最大值,而棉粕组在整个反应期间的磷含量均无显著差异(P>0.05)。同时,在反应进行到8 h时,鱼粉组的磷含量显著高于其他各组(P<0.05)。
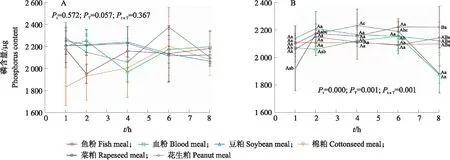
图2 6种饲料在日本沼虾肝胰腺(A)和胃肠道(B)粗酶提取液中的磷含量变化Fig.2 Phosphorus contents of six feed ingredients in the hepatopancreas(A)and gastrointestinal tract(B)crude enzyme extract of M. nipponense
3 讨论
3.1 日本沼虾肝胰腺和胃肠道粗酶液对原料中干物质、粗蛋白及总磷离体消化率的影响
干物质的消化率可反映动物对饲料总体的消化水平[18]。本试验中,日本沼虾肝胰腺粗酶液对豆粕中干物质的消化率最高,其次为棉粕和鱼粉;而胃肠道粗酶液对菜粕的干物质消化率最高,豆粕和鱼粉次之。上述结果表明,日本沼虾对植物蛋白源中干物质的消化率高于动物蛋白源。张南南[14]的研究结果表明,日本沼虾对5种饲料原料干物质的消化率分别为鱼粉11.24%、豆粕11.47%、棉粕12.33%、菜粕12.32%、花生粕8.73%,本研究结果与之相符。此外,在凡纳滨对虾(Litopenaeusvannamei)[7]离体消化率相关研究中也发现了类似结果。究其原因,可能是因为鱼粉中灰分含量较高,影响了虾体对其中干物质的消化率[16]。基于上述结果,在鱼粉资源短缺、饲料成本高昂的情况下,4种植物饼粕均可在一定程度上替代日本沼虾饲料中的鱼粉。
蛋白质是动物生长所需的关键营养物质。因此,在饲料营养价值评定中,动物对原料中蛋白质的消化率显得尤为重要[19-20]。综合来看,日本沼虾对棉粕粗蛋白的消化率最高,豆粕次之,二者均高于鱼粉,该结果与在鲫鱼(Carassiusauratus)[6]和凡纳滨对虾[7]的研究结果类似,推测原因,可能与日本沼虾的食性相关,一般情况下,与肉食性动物如卵形鲳鲹(Trachinotusovatus)[21]和兰州鲇(Siluruslanzhouensis)[22]等相比,杂食性动物对动物性原料中蛋白的利用率相对植物蛋白偏低。此外,由于饲料原料氨基酸组成的差异,消化酶对蛋白水解的位点及数量也不同,同一条件下的消化酶便会产生不同的酶解速度和程度[18],同时,也可能是由于鱼粉中蛋白质含量较高,超出了日本沼虾蛋白酶作用的最大底物浓度,继而造成其消化率相对偏低。相较于鱼粉,棉粕在水产动物饲料中的添加量较少,主要原因在于棉酚的存在、赖氨酸不足及粗纤维含量较高导致动物对其利用率低。但本文的结果却显示日本沼虾对棉粕具有较高的离体消化率。究其原因可能在于鱼虾相对低等,对棉粕中有害物质的耐受能力比家禽等恒温动物强[23],类似结果同样见于异育银鲫[24]。这进一步表明,豆粕和棉粕等植物饼粕可能是日本沼虾优质蛋白来源。
磷是构成软骨骼等的重要成分,是维持甲壳动物生长发育的必需矿物质之一。在饲料营养价值评定中,动物对原料中磷的消化也尤为重要。大量研究表明,植物原料中的磷大部分都以植酸磷形式存在,且其易螯合钙、镁、锌等金属元素,难以被动物肠道吸收利用。而在本研究中,日本沼虾肝胰腺和胃肠道粗酶液对菜粕中磷离体消化率较高,而对鱼粉中磷的离体消化率较低。究其原因,可能在于日本沼虾体内碱性磷酸酶活性相对较高,在钙、磷的消化吸收等过程中发挥重要作用[25],而碱性磷酸酶具有水解植酸磷的作用[26]。鲍蕾[27]的研究表明,适宜的日粮磷水平可显著提高日本沼虾肠道蛋白酶活性。由此可以推测,鱼粉中高含量的磷对日本沼虾肠道消化酶活性产生了抑制作用,影响了其对粗蛋白的消化率,符合上述结果。此外,鱼粉中的磷多为羟基磷石灰和磷酸钙形式,需要在酸性条件下才能被充分分解[28],因此胃肠道粗酶液对鱼粉中磷的离体消化率略高于肝胰腺。
3.2 日本沼虾肝胰腺和胃肠道粗酶液酶解饲料蛋白生成氨基酸总量的变化
蛋白质在被摄食后,需经消化生成氨基酸才能被动物吸收。因此,易于酶解的蛋白质才更易被动物利用。基于此,蛋白质酶解后氨基酸的生成率也可以用来评估蛋白质的营养价值。本研究结果表明,在经日本沼虾肝胰腺和胃肠道粗酶提取液酵解后,鱼粉和血粉的氨基酸生成量相对较高。究其原因,可能在于,与植物原料相比,动物原料中蛋白质的溶解度相对较高及氨基酸组成相对平衡。研究表明,饲料被酶解的速度除了与其组织结构和化学组成有关,还与饲料蛋白的溶解度及其氨基酸组成密切相关[29]。另外,鱼粉和血粉酶解后的氨基酸生成量较高,而其粗蛋白消化率较低。结果看似矛盾,实则可能是由于两者在酶解之前的游离氨基酸含量就远高于其他蛋白饲料,而过高水平的氨基酸抑制了虾体对粗蛋白的消化[6]。此外,棉粕和豆粕酶解后产生的氨基酸也相对较高,表明二者在日本沼虾配合饲料的开发中具有广泛的应用前景。粗酶液酶解后,血粉也具有较高的氨基酸生产量,但由于其干物质和粗蛋白消化率均较低,可通过补充限制性氨基酸及改善加工工艺(如喷雾干燥和日晒)等来提高其利用率[30]。
3.3 日本沼虾肝胰腺和胃肠道粗酶液中无机磷含量的变化
动物消化道内的总磷由采食后进入消化道的外源磷、消化液和肠壁细胞分泌的磷及消化道脱落细胞中的内源磷等组成。其中,无机磷在消化道内溶解后可直接被吸收利用,而有机磷则需经过水解酶的作用释放出无机磷后才可被吸收[31]。本试验中,各种原料经日本沼虾肝胰腺粗酶液处理后,无机磷含量均无显著差异。究其原因,可能在于磷的主要消化部位为肠道[32]。胃肠道粗酶液酶解1 h后,6种原料在粗酶液中的磷含量均呈上升趋势,可能是磷酸酶将有机化合物中的磷分离出来而逐渐溶解至粗酶液中所致[33];在酶解反应后期即6~8 h,除鱼粉外的其他饲料原料中磷含量大致呈现下降的趋势。究其原因,可能是随着反应时间的延长,酶浓度逐渐降低,无法进一步消化原料中的磷而导致磷含量下降;而鱼粉则由于其磷存在形式的影响,阻碍了酶解液对其消化而导致酶解液中磷含量不断攀升,这与上述磷的体外消化率结果一致。另外,不同原料酶解液中的无机磷含量达到峰值的时间点也有差异,可能与不同原料中钙水平及钙磷比有关[34]。
综上所述,4种植物性蛋白饲料中,日本沼虾消化器官酶解液对棉粕中粗蛋白和总磷的离体消化率较高,同时其氨基酸的生成量也较高。因此,在配合饲料制作过程中,可用棉粕替代部分鱼粉来降低饲料成本。另外,粗酶液酶解后,血粉氨基酸生成量较高,但干物质和粗蛋白消化率较低,可通过改善加工工艺等措施来提高其利用率。
猜你喜欢
畜禽业(2023年10期)2023-10-27 12:17:04
当代水产(2022年8期)2022-09-20 06:46:06
当代水产(2022年7期)2022-09-20 06:23:00
养殖与饲料(2022年7期)2022-07-14 06:53:44
当代水产(2022年6期)2022-06-29 01:13:12
当代水产(2022年6期)2022-06-29 01:12:02
当代水产(2021年8期)2021-11-04 08:49:10
饲料工业(2020年11期)2020-06-19 08:41:54
当代水产(2019年6期)2019-07-25 07:52:08
当代水产(2019年1期)2019-05-16 02:41:50
