
月季花形态建成及花期调控的研究进展
2023-05-17 06:47熊智颖于蕊胡子祎张玉满刘青林
中国农业科技导报 2023年2期
熊智颖, 于蕊, 胡子祎, 张玉满, 刘青林*
(1.中国农业大学园艺学院,北京100193; 2.中国科学院微生物研究所,北京100101)
中国古老月季的优秀品种在18 世纪先后传入英国和法国,通过复合杂交和多次回交,1876年育成了连续开花的现代月季 ,成为世界现代月季的里程碑和与古老月季的分水岭。1998—2018年间我国月季育种机构共计30 余个,育成月季新品种共计168 个,月季育种目标已经从最初只集中在外部形态(观赏性)上,如花型、花色、株型以及花香等,转变为除外部形态外,更多地关注月季的栽培品质,包括其耐寒性、耐热性、抗病虫害的能力以及瓶插寿命[2]。
开花是植物从营养生长向生殖生长转化的枢纽,在植物生长和发育中起核心作用。植物成花同时受内部生理状态和外部环境因素的双重影响,具有复杂的基因调控网络[3]。花朵的数目、形状、颜色及其开花习性是观赏植物的重要性状,决定了其观赏价值和经济收益。现代月季有着丰富的花型及花色,花型包括平开状、杯状、高芯状、高芯翘脚状、球状、莲座状;花色可以分为红色系、粉色系、黄色系、白色系、橙色系、黑色系、紫蓝色系、绿色系、复色系和表里双色系。月季还具有连续开花的性状,此性状源自我国的月季花(Rosa chinensis)和香水月季(Rosa×odorata),作为少有的能够连续开花的木本花卉,月季的花期调控途径以及花形态建成受到国内外学者的普遍关注[4-6]。本文对近几年月季花期调控与花形态建成的分子机制的研究进展进行了综述,以期为月季分子生物学研究和分子育种提供参考。
1 月季花发育研究进展
植物花发育是一个内部生理状态和外部形态发生复杂变化的调控过程,可以分为以下5 个发育阶段:①成花诱导(flower induction),植物从营养生长向生殖生长转变的过程;②花发端(flower evocation),花原基开始分化的过程;③花器官发育(floral organ development),花分生组织最后形成花器官;④花的成熟;⑤开花。
1.1 成花诱导途径
月季开花同时受环境因素与自身状态的影响,目前已知的成花诱导(花期调控)途径有光周期途径、春化途径、温敏途径、赤霉素途径、自主途径和年龄途径共6 条[5]。其中,光周期途径、春化途径和温敏途径受外界环境因素的影响,赤霉素途径、自主途径和年龄途径受自身状态控制,这些途径形成了彼此独立又相互交联的精密的成花调控网络(图1)。即使环境因子调控的开花途径,植物也有响应环境的受体、信号传导和靶基因等一系列内在的调控网络,本质上也受内在的生理和遗传控制。

图1 月季开花调控途径Fig. 1 Schematic diagram of flowering regulation pathway of rose
1.1.1 光周期途径 光周期途径是指植物依赖一定的日长或夜长而开花的调控途径。植物特异的光受体会感受不同波长的光,继而将光信号传递给生物钟,生物钟进一步将光信号输出给CO(COSTANS)。CO是FT(FLOWERING LOCUS T)和SOC1(SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1)的上游基因。FT被CO激活后,进而激活AP1(APETALA 1)、LFY(LEAFY)等花分生组织特异性基因,最终启动成花进程[7-8]。月季作为一种日中性植物,在长日照和短日照下均可开花。月季的日中性是受COL4(CONSTANS-LIKE 4)和CO分别在长日照和短日照条件下综合调控FT所获得的。在连续开花的‘月月粉Old Blush'中,RoCO在长日照条件下高表达,促进RoFT的表达,使月季在长日照下可以开花;RoCOL4则在短日照下高表达,通过与RoCO 蛋白的互作,促进RoCO与RoFT启动子CORE 元件的绑定,进而促进RoFT在短日照下表达,使得月季在短日照条件下也能开花,表现出日中性[9]。月季开花也会受光照强度的影响,表现为强光促进开花,弱光抑制开花。在弱光条件下,光信号通路的核心转录因子RoPIF1 (PHYTOCHROME INTERACTING FACTOR 1)、RoPIF3 和RoPIF4 与RoCO 互作,形成RoCO-RoPIF 复合体,抑制CO 与FT 启动子结合,从而抑制其开花;在强光下,则不会形成复合体,CO 会促进FT的表达,促进开花[10]。但也有研究表明月季并不是严格的日中性植物,月季品种‘Cara Mia'在长日照或连续光照下能促进营养生长,短日照可促进开花,属于数量型短日照植物[11]。
1.1.2 春化途径和自主途径 春化途径和自主途径是相对的,无需春化(和光周期)即可开花,即为自主途径。对不同开花习性的月季春化处理的结果表明,一季开花的月季必须经过春化处理才能开花;而连续开花月季‘Meijikatar'不经过春化处理也可以开花,但春化处理可以增加每个植株上花的数量[12]。本团队将连续开花月季‘月月粉'(Rosa chinensiscv. Old Blush)、‘梅朗口红'(Rosacv. Rouge Meilland)和一季开花月季‘变色粉团蔷薇'(Rosa multifloracv.Discoloring Cluster)、单瓣月季花(Rosa chinensisvar.spontanea)进行春化处理,也得到类似的结果,因此推测一季开花月季存在春化途径,而连续开花月季不需要春化,可能是自主途径。
1.1.3 赤霉素途径 赤霉素(gibberellins,GA)对调控植物开花具有重要的作用,虽然在许多草本植物中GA 可解除开花抑制,促进成花;但在月季中外施GA 会抑制其成花。Randoux 等[13]在不同季节分别对月季喷施外源GA 和GA 抑制剂多效唑(paclobutarzol, PCB),发现对一季开花月季喷施GA 会使其不能在春季开花;但在夏季对其喷施PCB,并不能促进一季开花月季在夏季开花。研究发现,月季中存在FT的同源基因RoTFL1c(RoKSN),该基因的启动子区域存在GA相应的cis元件,当对一季开花月季外施高水平的GA 时,会导致RoTFL1c大量表达,抑制下游AP1以及LFY等开花基因的表达,从而抑制开花;RoTFL1c是已被证实的控制连续开花性状的基因之一,在一季开花月季中能正常的积累与表达,而在连续开花的月季中由于RoTFL1c第2 个内含子中插入了1个9 kb的逆转录转座子,导致RoTFL1c的mRNA不能正常积累,使月季在夏秋季也可开花,获得连续开花习性[14-15]。但RoTFL1c的杂合度也会决定其对连续开花性状的影响程度,如‘紫枝玫瑰'(Rosa rugosa‘Purple branch')连续开花习性的获得是由于RoTFL1c杂合,表达量减半导致的[16-17]。事实上,蔷薇属连续开花的性状是受多基因控制的数量型质量性状,有一季开花的野蔷薇、一季半开花的玫瑰和部分藤本月季品种、多季和连续开花的现代月季[18]。
1.1.4 温敏途径 环境温度对于成花的影响表现为在一定温度范围内,高温促进成花而低温抑制成花,这一调控方式称为温敏途径[5]。温室切花月季的最低生长温度为16 ℃,在≤10 ℃时生长缓慢,冻害(霜、雪、冰冻)和≤10 ℃的天气过多会使“盲枝”率大幅提高;在≥30 ℃时生长不良。当生长期温度在18~30 ℃时,月季从修剪到开花的时间随温度升高呈线性缩短;低夜温和低日温会延迟花芽形成和开花时间[19]。显然,切花月季的成花受温敏途径的调控,但相关的基因和调控机理尚未见报道。
1.1.5 年龄途径 年龄途径是一条与年龄增长密切相关的开花调控途径。SPL(SQUAMOSA promoter-binding protein-like) 家 族 和 miR156(microRNA 156)、miR172(microRNA 172)是年龄途径的重要调控基因[20-21]。与月季花品种‘月月粉'相比,‘绿萼'(R. chinensiscv. Viridiflora)中的miR172在花瓣、雌蕊、雄蕊中表达均显著降低,预测其靶基因RcAP2的表达量显著升高,同时也预测了miR156的靶基因为SPL[22]。对月季基因组内SPL家族成员进行检索分析表明,月季中共含15个SPL基因,转录组分析发现,RoSPLa可能参与调控成花转变,将RoSPLa转入拟南芥中过表达,可使转基因植株开花时间提前[23]。月季杂交育种的实践表明,如果杂交后代在当年开花,则为四季开花的月季;反之,如果杂交后代在第2 年才开花,则为一季开花的蔷薇。月季的童期只有4~5周,而蔷薇的童期超过1年,这种童期的巨大差异是年龄途径相关基因表达的变化还是越冬的春化作用,有待进一步研究。
通过媒体传播是塑造城市文化形象的基本手段。影视媒介研究表明,采用系列影视片能够对受众在传递信息和影响人们观点、意见方面有着明显的效果。因此,本地区主流媒体更应该深入厂区、发掘历史、深挖典型人物、典型事件、结合当前党中央、国务院大力振兴东北老工业基地精神,制作出人民群众所喜闻乐见的影视纪录片,擦亮齐齐哈尔“国家工业明珠”这张亮丽的城市文化形象名片。
1.2 花芽分化
1.2.1 花芽分化的时期 植物茎尖分生组织接收到成花诱导信号后就进入了花发端阶段,开始进行花芽分化。花芽分化一般分为生理分化和形态分化2 个过程。芽内生长点在生理状态上向花芽转化的过程,称为生理分化,生理分化为花芽分化提供生理环境和物质基础。此后开始的花芽发育的形态变化过程,称为形态分化,形态分化能更直接的确定花芽分化及发育的关键时期[24]。环境因子会影响花芽分化进程,如不同品种月季在不同温度下花芽分化持续时间不尽相同,在一定温度范围内,温度越高花芽分化所需时间越短[25];遮荫可以在一定程度上延缓‘梅郎口红'(Rosacv.Rouge Meilland)花芽分化进程[26]。
目前以石蜡切片作为观察花芽分化的主要手段。胡祖坤[25]制作并观察了月季花芽的石蜡切片,将月季花芽从分化前期到开花前期分为了16 个时期。蔷薇属植物的成花转变一般可划分为3 个生长期:①营养生长期-腋芽的茎尖分生组织出现三角形小突起;②营养生长锥转化期-分生组织迅速扩张,由营养生长组织转变为生殖生长组织;③生殖生长锥分化初期-分生组织向上突起逐渐加宽伸长呈宽圆锥形[27-28]。对‘梅郎口红'的花芽用体视显微镜结合石蜡切片观察的方法,可将营养生长锥转化期和生殖生长锥分化初期细分为:①萼片原基分化期-萼片原基分生组织细胞基本形成;②花瓣原基分化期-萼片原基伸长、变宽且向内弯曲,花瓣原基分生组织朝着各个方向分裂;③雄蕊原基分化期-杯状花托上部形成多轮雄蕊原基,雄蕊群轮状分布,多层分化;④雌蕊原基分化期-雌蕊原基边缘的细胞体积小、数量大且分裂旺盛,随着发育的进行而形成更多雌蕊原基[26]。通过花芽大小也可大致区分花芽分化阶段,通过对‘月月粉'花芽进行石蜡切片发现腋芽长度8 mm 是重要节点之一,当腋芽长度大于8 mm 时,月季才普遍开始花芽分化[29]。
1.2.2 开花整合子 虽然存在多种成花诱导调控途径,每条调控途径都能独立完成从营养生长到生殖生长的转变,但是又彼此关联,多条途径最终作用于LFY、FT、SOC1等基因,促进植物花芽分化,这类基因被称为开花整合子。RoLFY作为花分生组织特性基因,促进花序分生组织向花分生组织的转变,维持花分生组织的正常功能与成花启动,上游受开花抑制子TFL1基因的调控,下游可促进开花整合子AP1的表达,从而促进植物成花。不同开花习性月季LFY的表达量存在显著差异,在花芽形成至开花这一发育阶段,在连续开花月季中LFY的表达量一直在降低,在一季开花月季中LFY表达量先升高后降低[30]。FT 可与LFY共同促进AP1的表达从而促进成花,FT 蛋白可从韧皮部运输至花分生组织,激活花分生组织特性基因的表达[31]。在油用玫瑰‘Bao White'中转入梅花的FT基因,其开花时间明显提前[32]。SOC1属于MADS-box家族成员,在月季各器官中均有表达,但在顶芽中表达量最高。在拟南芥中过表达RoSOC1,可使转基因植株开花时间提前[33]。因此,多条成花诱导途径共同作用于开花整合子,形成一个彼此关联、错综复杂的成花调控网络。
1.3 花形态建成分子生物学进展
1.3.1 花器官发育 决定月季花形态建成的基因是花器官特性基因(MADS-box同源异型基因),包括A、B、C、D 和E 类功能基因,构成ABCDE 模型。对ABCDE 模型可简要解释为,A+E 调控萼片发育,A+B+E 调控花瓣发育,B+C+E 调控雄蕊发育,C+E 调控心皮发育,C+D+E 调控胚珠发育,A 和C相互拮抗[34](图2)。在月季中,部分控制花器官特性的MADS-box基因已被克隆并进行功能验证,如A 类 基 因cAP1、RhFUL、RcAP2L[35-36],B 类 基 因RhPI、RhAP3、MASAKO B3、MASAKO BP[37],C 类基因RhAG、MASAKO C1等[38-39]。通过转录组也检测到了月季D 类基因RhSTK(SEEDSTICK)、RhSHP(SHATTERPROOF),E 类 基 因MASAKO S1、MASAKO S3[40]。对于月季花形态建成的研究,主要集中在重瓣性与绿萼这2种特殊的观赏性状上。
1.3.2 重瓣性是A、C 类基因表达边界位移的结果 花瓣的数量对于观赏植物的花型和观赏价值有着重大意义,重瓣也是花期延长的重要基础。月季的重瓣性状为单个基因控制的显性性状;花瓣的数量受多个数量性状基因位点(quantitative trait loci, QTLs)控制,温度、外源激素等外界信号也会对重瓣性有一定的影响。目前普遍认为月季的重瓣性状以雌雄蕊瓣化为主,是外部环境与内源调控共同作用的结果[41-42]。在月季中,通过定位确定了与重瓣性状有关的遗传区间,该区间存在A 类功能基因RcAP2L(APETALA2-like gene)的突变等位基因,该等位基因对miR172 不敏感,存在TE 的插入[36]。通过比较单瓣花与重瓣花中是否存在该插入,确定了该等位基因的存在与重瓣性状有关。研究表明,月季的重瓣性主要受AG、AP2等基因的控制,在重瓣花中,RhAG的表达量较单瓣花低,原位杂交发现,在重瓣花中RhAG的表达域内缩,从而造成花瓣数量增多,雄蕊数量减少[43]。同时,低温也会通过提高RhAG启动子特定区域的甲基化水平,降低RhAG的表达量,从而增加花瓣数量[44]。
1.3.3 ‘绿萼’的形成与A、C和E类基因的表达变化有关 ‘绿萼'是中国古老月季的代表品种,其花型奇特,花瓣、雌雄蕊均为绿色萼片状。‘绿萼'(R. chinensis‘Viridiflora')是花器官的同源异型突变体,花瓣、雄蕊、雌蕊均突变为萼片(萼片化)。1981 年,Krussman[45]报道‘绿萼'是单瓣月季花(R.chinensis‘Jaquin')的自然突变体。对于‘绿萼'形成的原因有2种推测:(1)C类AG基因在雌雄蕊的表达受到抑制[46],A、C类基因的异常表达导致花器官的萼片化。试验证实‘绿萼'中A 类基因RcAP2在雌雄蕊中表达量显著升高,C 类基因RcAG的表达量显著下降,但RcAG如何被抑制、RcAP2的表达量如何上升的分子机制还不明确[47]。(2)E 类MASAKO S3基因在4轮花器官的超表达,使得花器官均发育成萼片(图2)。无独有偶,拟南芥的E类基因sep1、sep2和sep3三重突变体的雄蕊和心皮均转化为萼片[48],与月季花‘绿萼'表型很相似。
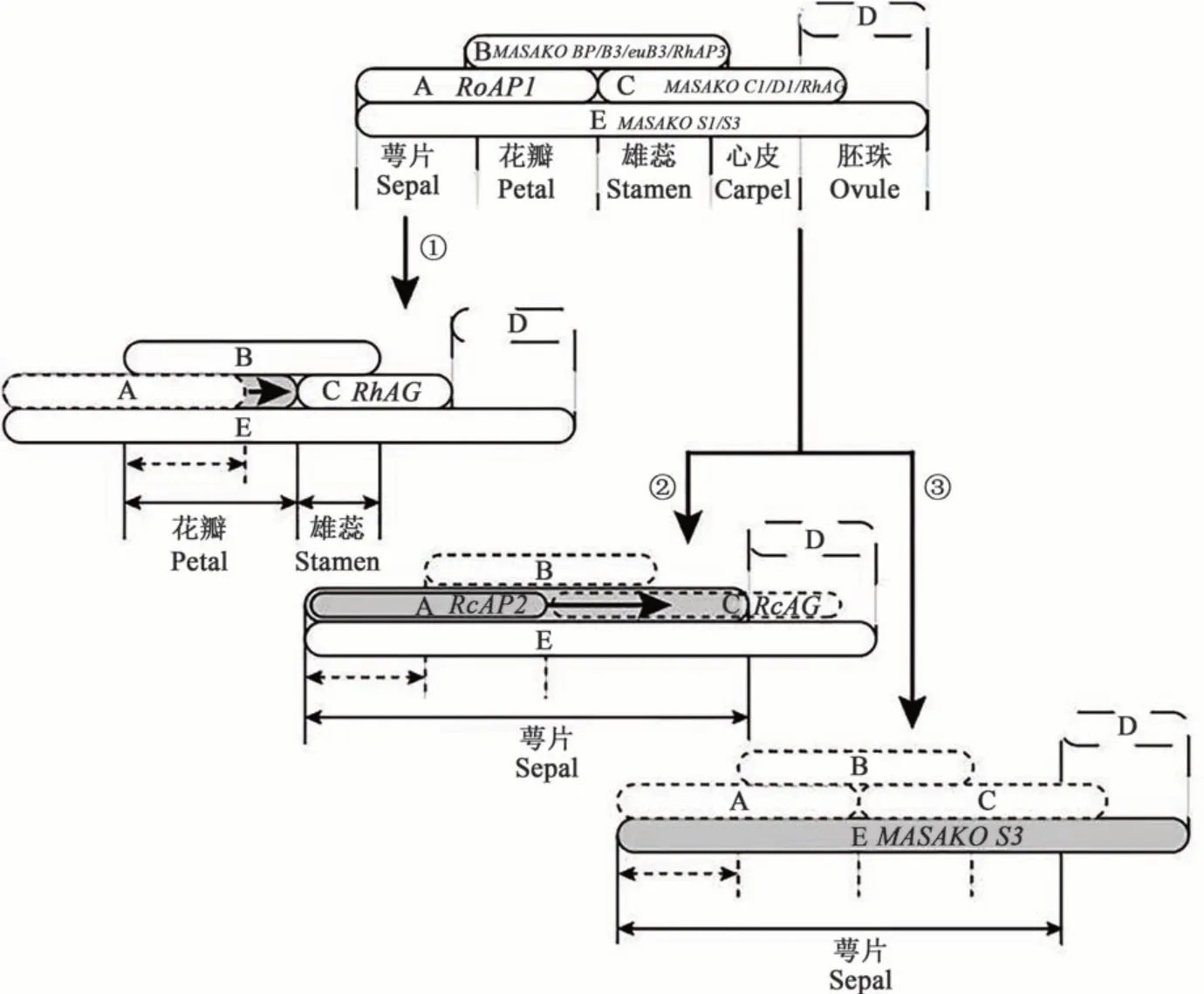
图2 月季花器官发育的ABCDE模型Fig. 2 ABCDE model of organ development in rose
2 花期调控途径
2.1 温度
气温是调控月季开花的重要因子,月季属于常绿或半常绿植物,高温干旱会促使月季进入休眠或莲座化,低温有利于恢复月季植株的生长活性[49]。需要对月季花期进行调控时,可根据预计花期,依据当地情况,对温度进行调节,使夜温保持在8~10 ℃,并根据月季植株和花蕾生长情况,进行昼夜温度调节。当花的采收高峰提前时,白天开窗降温,使温室内温度维持在20~22 ℃,夜间温度维持在8~9 ℃,可延缓花的采收时间;当花的采收高峰推后时,白天温室中进行温度调节,使温室内温度维持在24~26 ℃,夜间温度维持在12~13 ℃。进入采收期后注意夜间保温和加温,并调节温度减小昼夜温差[19]。
2.2 生长调节剂
植物激素参与调控月季生长发育的各个方面,向月季幼枝喷施GA3可以解除休眠,增加开花枝条的数量,促成长花枝。研究证明喷施300 mg·L-1的GA3,对株高、花枝长、花柄长均有促进作用,增加了叶片长宽比[50]。用1~10 mL·L-1的1-甲基环丙烯(1-methylcyclopropene,1-MCP)熏气处理数小时也能有效延长月季的开花时间[51]。此外,利用乙烯利在月季展叶期进行喷洒或浇灌,可以延长或促成月季开花[52]。
2.3 修剪措施
在实际工作中,露天月季因生长温度无法调节,主要依靠修剪来控制月季花期,提高月季花的数量和质量。修剪要遵循“疏弱留强、去横留直”的原则,有利于维持树势及植株更新,提高花的品质。在调控月季花期时,除了根据月季植株综合长势、修剪季节温度和植株品种特性外,还要根据花芽的发育情况选择适宜的修剪部位,以确保按预期开花和花的质量。在适宜月季生长的季节,轻度修剪(修剪花下2~5 叶处),发出花枝短,现蕾早,修剪后20 d左右开花,花朵较小;中度修剪(修剪花下6~9 叶处),圆芽发出的花枝长,现蕾时间较长,修剪后30 d 左右开花,花朵大;重度修剪(修剪枝条基部),芽活性低,发枝慢,易发徒长枝,花枝现蕾时间通常为40 d,花朵较大[53]。
3 结语
蔷薇属不同种类甚至同一种的不同品种都具有不同的开花习性,这里涉及到成花转变和花芽分化。赤霉素途径通过GA促进RoTFL1c的表达,抑制下游开花整合子的表达而抑制开花,形成一季开花;反之,反转录转座子插入引起RoTFL1c的突变,是连续开花的分子基础。长日照条件下调控的RoCO和短日照条件下调控的RoCOL4是月季感受光周期信号,表现日中性的分子基础。月季的开花习性与低温、适温和童期有关,其成花转变表现出与春化途径、年龄途径和温敏途径的相关性。杂交试验遗传分析的结果也表明,月季的开花习性由多基因控制。可见,寻找RoTFLC1c和赤霉素途径以外的开花途径和开花习性相关的基因,是目前的研究重点。
月季的重瓣性和绿萼这2 种性状与ABCDE模型中相关基因的突变有关,其中重瓣性受AG、AP2等基因的控制,绿萼的出现与A、C 类基因的异常表达有关,还可能与E类基因的过表达有关。重瓣性既是月季观赏价值的重要体现,也是低温伤害的结果和花朵开放的障碍,对于切花月季产业具有重大影响,需要找到有效的调控途径。
月季具有生命周期短、花芽分化时间短、高度可变的倍性水平以及较完善的基因组,可作为木本植物花发育研究的模式植物[5]。目前,由于月季的转化体系不够稳定,转基因植株获得困难,因此大多数月季基因的功能验证在异源系统中进行。同时,月季的品种较多,杂合性较高,已测得的基因组和转录组数据十分有限。今后可从基因组和转录组信息挖掘新基因,对成花关键基因的表达模式、成花转录因子的靶点分析等方面进行研究,不断推进对月季成花机理和花形态建成的全面和深入理解。
猜你喜欢
蔬菜(2022年1期)2022-11-11
麦类作物学报(2022年4期)2022-05-23
金山(2021年10期)2021-11-02
文萃报·周五版(2021年28期)2021-08-19
知识就是力量(2021年6期)2021-07-09
热带生物学报(2019年1期)2019-04-18
江苏农业科学(2018年11期)2018-06-29
新课程·下旬(2018年7期)2018-01-19
现代园艺(2017年23期)2018-01-18
上海农业学报(2017年3期)2017-04-10
