
牡蛎鳃部腐败菌的分离鉴定及产胺特性分析
2023-05-17 02:58:36何诗盈谢飞鸿苏世京黄鹭强陈慧斌
福建师范大学学报(自然科学版) 2023年3期
何诗盈,孟 鹏,谢飞鸿,苏世京,黄鹭强,陈慧斌
(1.福建师范大学生命科学学院,福建 福州 350117;2.福建省产品质量检验研究院,福建 福州 350002)
牡蛎肉质鲜美,富含大量的蛋白质、糖原、牛磺酸、多糖等营养物质[1],具有促进机体血液循环、保护肝脏等一系列药用价值[2],是重要的海洋生物资源之一.牡蛎属于药食同源的食物,牡蛎的市场需求庞大[3].然而牡蛎的含水量、蛋白质含量高,其中蛋白质含量超过其软体部分的50%[4],腐败菌代谢旺盛,且常发生自溶现象,导致牡蛎极易腐败.此外,开壳后的牡蛎组织易受多种微生物的侵害而发生一系列生化反应,例如牡蛎体内的ω-3不饱和脂肪酸被迅速氧化成有害物质,造成严重的经济损失[5],并且过量的生物胺对人体产生危害[6].
生物胺是一类具有生物活性的低分子质量碱性含氮化合物,主要由氨基酸脱羧酶脱羧形成,根据结构的不同分为脂肪族(腐胺、尸胺、精胺和亚精胺等)、芳香族(酪胺、β-苯乙胺等)、杂环族(组胺、色胺等)[7-8].蛋白质降解后所产生的前体氨基酸在氨基酸脱羧酶作用下生成相应的生物胺[7],如图1.适量的生物胺有助于调节身体,但若摄入过量的生物胺则会对身体产生一定的危害,如组胺对人体影响最大,过量摄入会引发中毒现象[9],酪胺可引起中毒性肿胀,其他生物胺不仅会产生头痛、恶心、高血压等生理性症状[10],还会加重组胺和酪胺的不良作用,因而需从源头控制生物胺的产生[11].
据报道,Pseudomonas、Vibrio、Shewanella等菌是主要的腐败产胺微生物[12],能够产生精胺、腐胺、尸胺、酪胺等8种主要生物胺[13],降低牡蛎的新鲜度.Cao等[14]使用高通量测序技术证实300 MPa的高压处理和pH处理能够显著改善其微生物组成,将太平洋牡蛎的贮藏期延长至12 d.Chen等[15]通过对比不同条件的气体包装,证实CO2与O2体积比为7∶3最有利于牡蛎贮藏,抑制牡蛎的腐败.
据陈慧斌[16]研究,牡蛎依靠鳃部获取食物,鳃部含有种类数量丰富的微生物,并且微生物对牡蛎的新鲜程度影响较大,由此对牡蛎鳃部微生物分离有一定的必要性.牡蛎所产生物胺对食用安全造成威胁,但关于腐败菌群与产胺的关系却鲜有报道,本实验通过分离纯化牡蛎中的主要腐败产胺菌,探究控制牡蛎鳃部生物胺的质量浓度以延长牡蛎贮藏期.

图1 氨基酸与生物胺的关系Fig.1 Relationship between amino acids and biogenic amines
1 材料与方法
1.1 牡蛎
产自连江海域,从市场购得长度为10~15 cm的带壳牡蛎,无菌环境下挑取组织完整的牡蛎,于(4±3)℃冷藏4 d后,取鳃部组织进行腐败菌分离鉴定.
1.2 试剂与仪器
1.2.1 试剂
无水乙醇,国药集团化学试剂有限公司;四硼酸钠,购自上海成海化学工业有限公司;氯化钠、乙腈均购自西陇科学股份有限公司;硼酸,购自广东光华化学厂有限公司;0.9%氯化钠注射液,购自福州海王福药制药有限公司;乙醚,购自衡阳市凯信化工试剂有限公司;L-赖氨酸、L-组氨酸、甘油均购自Solarbio公司;L-色氨酸、L-酪氨酸、L-苯丙氨酸、色谱级甲醇,均购自Mackin公司;L-鸟氨酸购自Acmec公司;L-精氨酸购自Sigma-Aldrich公司;四盐酸精胺、色胺、酪胺盐酸,均购自上海源叶生物科技有限公司;尸胺、三盐酸亚精胺、β-苯乙胺、二盐酸组胺,均购自Dr.Ehenstorfer公司;平板计数琼脂(PCA)培养基、营养琼脂(NA)培养基、2216E液体培养基、LB肉汤培养基,均购自青岛海博生物;细菌基因组DNA提取试剂盒,购自北京全式金生物技术有限公司.
1.2.2 仪器
HC-2518R高速冷冻离心机(安徽中科中佳科学仪器有限公司);MJ-78A高压灭菌锅(施都凯设备(上海)有限公司);LRH-150生化培养箱(福州科远贸易有限公司);IS-RDD3恒温振荡器(CRYSTAL公司);KQ-500DE数控超声波清洗器(昆山市超声仪器有限公司);HH-3A数显恒温水浴锅(金坛区易晨仪器制造有限公司);e2695高效液相色谱仪(Waters公司);MiniAmp PCR仪(Applied biosystems公司);DS-11超微量分光光度计(DeNovix公司);荧光多模式酶标仪(Tecan公司)等.
1.3 实验方法
1.3.1 鳃部细菌的分离纯化
无菌条件下称取11.0 g样品,加入89 mL生理盐水进行稀释,取1 mL稀释液梯度稀释至质量浓度为10-2~10-7g·mL-1,选取3个合适的质量浓度梯度菌液,分别吸取100 μL涂布于PCA培养基、2216E培养基、NA培养基,每个培养基做3组平行对照组,分别放入28 ℃和4 ℃培养箱中培养.观察菌落形态,挑取单菌落划线于LB平板,每个筛选的菌株纯化3次,以获取单一菌落[16].将纯化后的单菌落接种于5 mL LB液体培养基中,30 ℃、180 r·min-1条件下摇床培养24 h后,取500 μL菌液与50%的甘油以体积1∶1的比例混合于冻存管,于-80 ℃保存.
1.3.2 菌株的鉴定及进化树的构建
全基因组提取步骤参照北京全式金生物技术有限公司的细菌基因组DNA提取试剂盒进行,并扩增细菌16S rDNA.通用引物为27F /1492R,27F:5′-AGAGTTTGATCCTGGCTCAG-3′,1492R:5′-TACGACTTAACCCCAATCGC-3′.PCR反应条件:94 ℃预变性5 min;94 ℃持续变性30 s,55 ℃退火30 s,72 ℃延伸45 s进行35个循环,最后72 ℃延伸10 min.琼脂糖凝胶电泳和超微量分光光度计检测DNA的质量浓度与纯度,纯化后样品送至生工生物工程(上海)有限公司测序.NCBI-BLAST比对测序菌株,选取相似性最高的菌株,利用MEGA-X软件通过NJ法构建成系统发育进化树,其他参数默认.
1.3.3 生物胺的衍生及高效液相色谱分析
添加鸟氨酸、精氨酸、赖氨酸、组氨酸、赖氨酸、色氨酸、苯丙氨酸于LB液体培养基,由于酪氨酸难溶于水,所以酪氨酸质量浓度为300 mg·L-1,其余氨基酸质量浓度均为500 mg·L-1.将活化2次的菌液取100 μL转移到添加含氨基酸的LB培养基中,于30 ℃条件下摇床(180 r·min-1)培养18 h.将5 mL菌液在5 000 r·min-1、室温的条件下离心5 min,固液分离,提取上清液3 mL用于衍生.
标准品液相图如图2,生物胺衍生的步骤参照Liu等[17]方法.高效液相色谱采用中谱科技RD-C18柱(4.6 mm×250 mm,5 μm),柱温35 ℃,紫外检测波长为254 nm,流量为1.0 mL·min-1,进样体积为10 μL,流动相A为超纯水,流动相B为甲醇,梯度洗脱程序如表1.
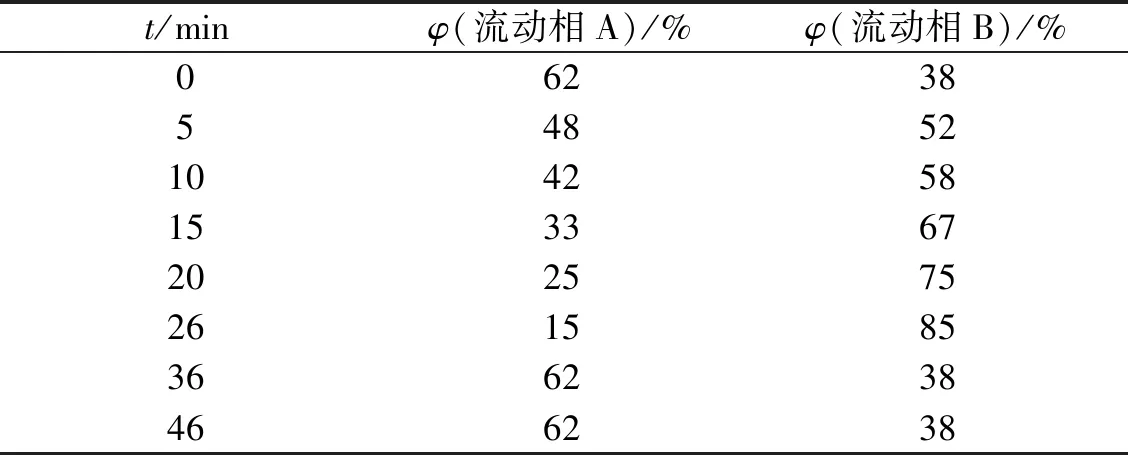
表1 梯度洗脱程序Tab.1 Gradient elution procedure
以时间为横坐标,生物胺与内标的峰面积之比为纵坐标,建立标准曲线,拟合直线,固定截距为0.生物胺的回归方程和相关系数如表2.根据回归方程,计算样品生物胺质量浓度和产胺能力,产胺能力计算公式如下.利用Origin 2019处理样品数据并绘制热图.


表2 8种生物胺的回归方程和相关系数Tab.2 Regression equations and correlation coefficients of 8 kinds of biogenic amines
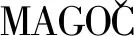
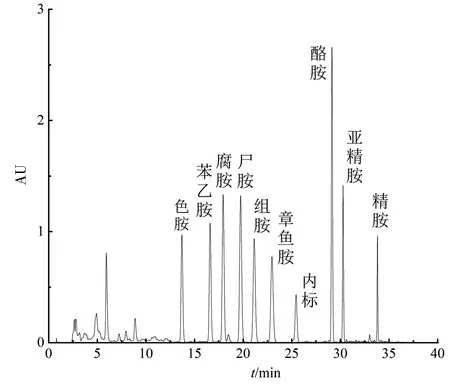
图2 标准品液相图Fig.2 Chromatogramof standard solutions
2 结果与讨论
2.1 腐败菌的分离鉴定
2.1.1 16S rDNA序列的分析
对16株菌的16S rDNA进行测序,它们属于6种菌属,分别为1株Kocuria、1株Aeromonas、1株Oceanisphaera、2株Priestia、2株Marinomonas、9株Shewanella,结果如表3.
2.1.2 进化树的构建及分析
如图3所示,进化树表明A5和A10、A7和A9为同一支,相似性均不低于95%;Shewanella与其他菌属之间的相似性较低,表明它们之间的亲缘关系较远;在不同菌属之间,Kocuria、Oceanospirillum和Priestia之间的相似性均在90%以上,说明亲缘关系较近.
据系统发育树显示A8属于Kocuria菌属,相似性为100%.A2、A4分别属于Oceanisphaera和Aeromonas菌属,与其对应的标准菌株的相似性为100%,结果表明测序菌株基本属于对应的标准菌株;A6与其对应的标准菌株的相似性为88%,说明A6大概率属于Shewanella菌属;其余测序菌株与对应的标准菌株的距离较远,说明测序序列与标准菌株序列有一定的差异.
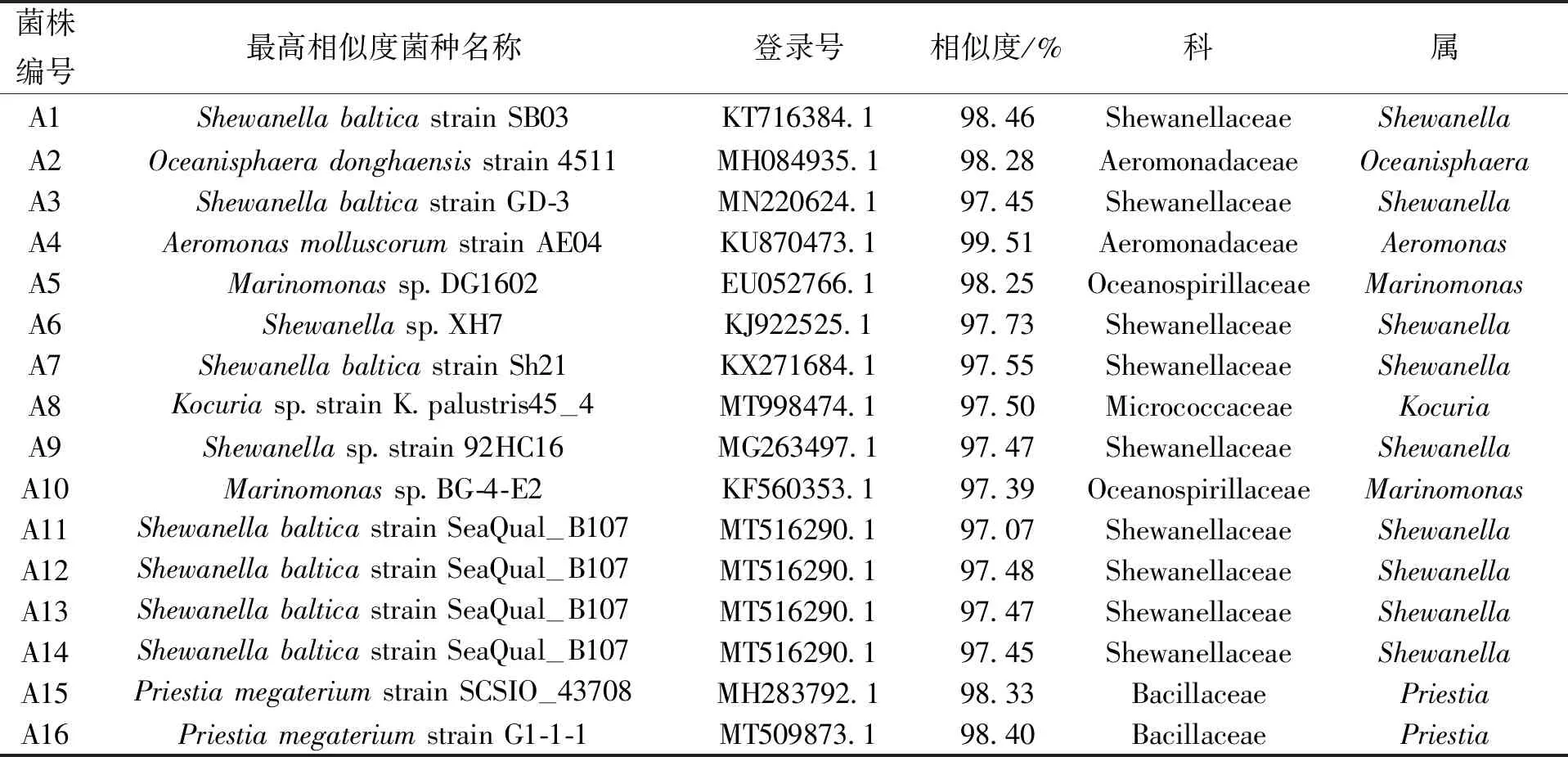
表3 16株分离菌测序结果Tab.3 Sequencing results of 16 isolates
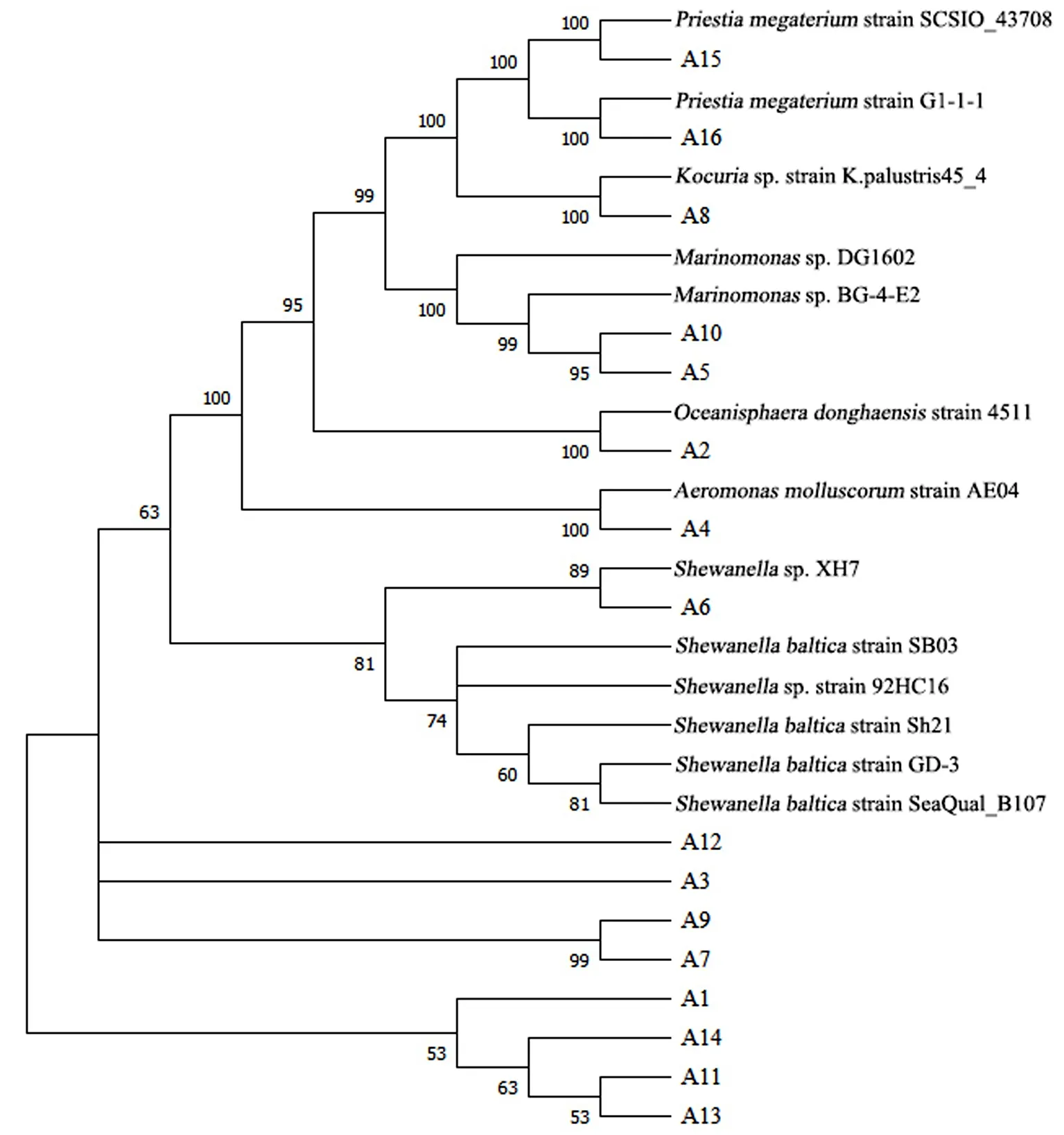
图3 16S rDNA系统发育进化树Fig.3 16S rDNA phylogenetic tree
2.1.3 属水平下的细菌群落组成分析
将牡蛎鳃部样品(0,4 d)分为2组样品(G0,G4),细菌菌群组间排名前10的细菌群落结构分布情况如图4所示,2组样品的菌属相对丰度有一定差异,说明4 d的贮藏期内牡蛎鳃部的优势菌不断变化.从4个样品中共得到18个优势菌属,在2组样品组内间也呈现出个体物种丰度的差异,其中G0的2个样品的优势菌种为Shewanella、Colwellia、Mitochondria、Vibrio、Pseudoalteromonas.G4的2个样品优势菌种为Pseudoalteromonas、Shewanella、Colwellia.在4个样品中共有的优势菌属为Shewanella、Pseudoalteromonas、Colwellia.
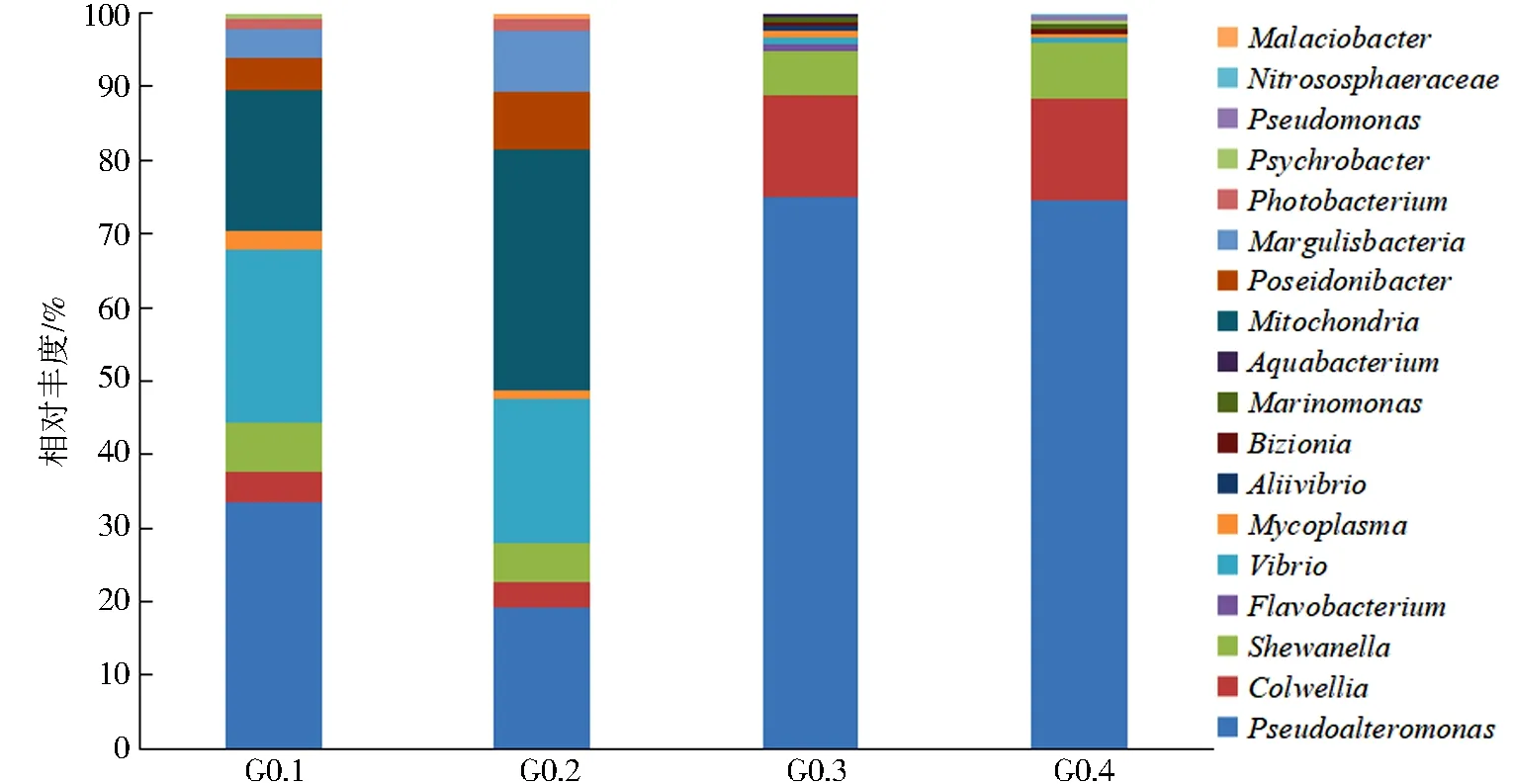
图4 牡蛎鳃部样品(0,4 d)在属水平下的细菌群落结构Fig.4 Bacterial community structure at the genus level for oysters gill samples (0,4 d)
2.2 腐败菌产胺能力分析
牡蛎含有丰富的蛋白质,在自身氨基酸脱羧菌作用下会产生大量的生物胺,加速牡蛎的腐败程度.为分析从牡蛎鳃部筛选得到的16株腐败菌,将其进行摇床培养,并加入适量的氨基酸促进其合成生物胺,通过高效液相色谱分析腐败菌所产的8种胺质量浓度如表4,总胺质量浓度及产胺能力如图5.

表4 各菌株生物胺质量浓度Tab.4 Biological amine mass concentration of each bacterial strain
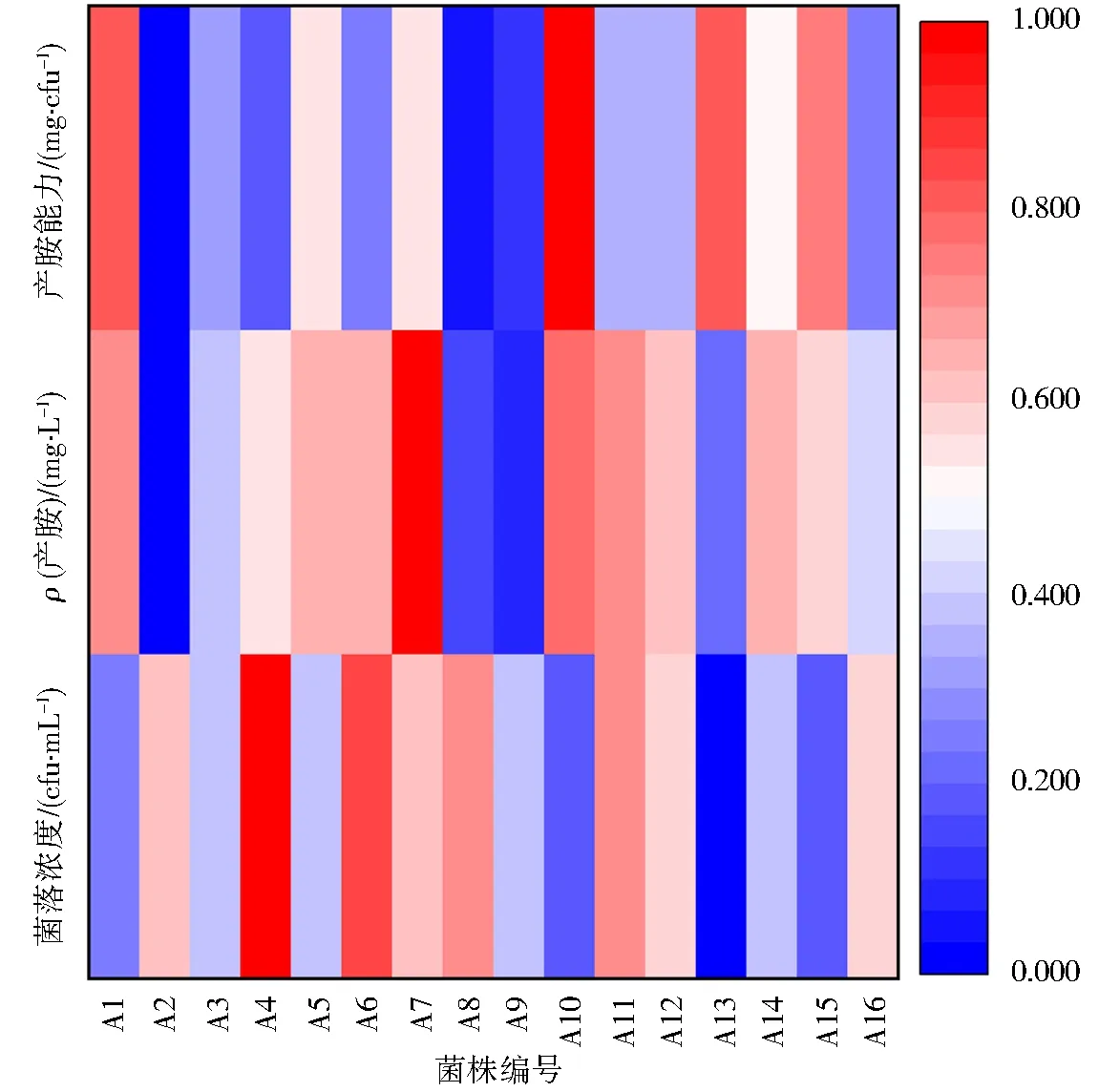
图5 产胺能力的热图分析Fig.5 Heatmap of amine producing ability
从培养基中均能检测出多种生物胺,但所产生物胺的种类和数量差异较大.其中除A2不产腐胺、尸胺、色胺,A13不产腐胺、尸胺、色胺和亚精胺,其余腐败菌均能产生8种生物胺.值得注意的是,A6和A7所产腐胺质量浓度分别高达(82.21±9.65)和(111.19±8.78) mg·L-1,均属于Shewanella;A1所产β-苯乙胺质量浓度高达(43.89±21.74)mg·L-1,属于Shewanella;A10所产酪胺质量浓度高达(68.99±9.99) mg·L-1,属于Marinomonas;产亚精胺、精胺和组胺质量浓度高于10 mg·L-1的菌株不超3株,表明牡蛎的腐败过程中这3种生物胺的积累不多;各菌株所产腐胺、色胺、β-苯乙胺和酪胺的质量浓度较大,可能在牡蛎腐败过程中过多积累.
水产品的腐败程度通常以生物胺含量来衡量,生物胺的含量越高腐败程度就越高,摄入过量生物胺会导致中毒,组胺毒性最大,酪胺会引起毒性肿胀,其余生物胺会加重组胺和酪胺的毒性.在16株菌株中,A7、A10所产生物胺总量较高,质量浓度分别高达169.25、136.70 mg·L-1;A4的菌落浓度最高,为5.70×108cfu·mL-1.生物胺含量、菌落浓度与腐败菌的产胺能力紧密相关,通过比较16株菌的菌落浓度及总生物胺含量以分析腐败菌的产胺能力.不同腐败菌所产生物胺种类和含量不同,结合产胺质量浓度和菌落浓度可计算出腐败菌的产胺能力.据热图分析结果显示,16株腐败菌的产胺能力差距较大,其中A10、A13、A1、A15均属于强产胺菌,且致腐能力由强到弱;A2的产胺质量浓度和产胺能力最低,属弱产胺菌;其余菌株属于次产胺菌.结合热图综合比较得出Shewanella、Marinomonas菌属的致腐能力较强,明显高于Oceanisphaera、Aeromonas、Priestia、Kocuria,由此推测Shewanella和Marinomonas两个菌属积累过多的生物胺,也是导致牡蛎发生变质的重要原因.对牡蛎鳃部样品(0,4 d)进行高通量测序,筛选分离得到的16株腐败菌中Marinomonas、Oceanisphaera、Aeromonas、Priestia、Kocuria菌属在高通量测序中相对丰度低,Shewanella、Colwellia和Pseudoalteromonas是牡蛎鳃部的优势菌种,然而在分离纯化牡蛎鳃部微生物过程中并未筛选到Pseudoalteromonas和Colwellia菌属,这可能是由于牡蛎中Pseudoalteromonas菌属无法在特定培养基生长[20],Colwellia菌属为兼性厌氧,在大气压下难以培养[21].此外,Shewanella的不同菌株腐败能力不同,其中A11、A12、A13、A14经NCBI-BLAST比对得到的最高相似度菌种相同,然而菌株序列与对应的标准菌株序列有一定的差异,并不能确定这4株菌在种水平上为同一种,且产胺特性结果也不相同.
3 结论
牡蛎鳃部的微生物群落复杂,高通量测序结果显示牡蛎鳃部的优势菌种为Shewanella、Pseudoalteromonas、Colwellia,从牡蛎鳃部分离得到的16株菌经16S rDNA鉴定后结果显示其分别属于Shewanella、Oceanisphaera、Aeromonas、Marinomonas、Kocuria和Priestia菌属,这可能是由于Pseudoalteromonas和Colwellia不易培养所致.腐败菌所产主要生物胺为腐胺、色胺、β-苯乙胺和酪胺,因而推测腐胺、色胺、β-苯乙胺和酪胺是造成牡蛎腐败的主要生物胺,亚精胺、精胺和组胺在牡蛎腐败的过程中积累较少.综合分析产胺质量浓度、产胺能力和各生物胺产生情况可得,Shewanella和Marinomonas的致腐能力较强,对牡蛎的影响最为严重,重点研究腐败菌的产胺机制有助于提高牡蛎贮藏期.大部分食品原本含有生物胺,且在微生物的氨基酸脱羧酶作用下不断积累生物胺,如在巧克力[22]、奶酪[23]、葡萄酒[24]中均检测出来生物胺.在贮藏运输的过程中食品可能感染腐败菌而产生过量生物胺,探究出抑制腐败菌的产胺代谢的方法,将有益于延长海鲜、酒类等食品的贮藏期.
猜你喜欢
食品工业科技(2024年6期)2024-03-14 08:03:32
基层中医药(2021年7期)2021-11-02 07:19:54
食品科学(2021年19期)2021-10-31 11:26:18
食品工业科技(2020年24期)2020-12-09 07:36:24
华南农业大学学报(2017年4期)2017-08-07 06:25:16
饮食与健康·下旬刊(2017年4期)2017-05-26 20:00:47
大众考古(2015年6期)2015-06-26 08:27:16
饲料工业(2015年11期)2015-01-19 02:26:36
小说月刊(2014年8期)2014-04-19 02:39:15
饲料工业(2014年2期)2014-01-21 11:31:36
