
芜菁BrrLOX7基因克隆、表达及生物信息学分析
2023-05-04 05:24燕存尧闫会转
浙江农业学报 2023年4期
燕存尧,贾 凯,闫会转,高 杰
(新疆农业大学 园艺学院,新疆 乌鲁木齐 830052)
芜菁(BrassicarapaL. subsp.rapa),俗名蔓菁、盘菜、恰玛古等,为十字花科两年生草本植物,因具有食用、药用和饲用价值而被广泛种植[1]。南疆地区是芜菁的主产地,土壤盐碱度大,而且芜菁的栽培季节主要在秋冬季,其在生长过程中常受到干旱胁迫、盐碱胁迫以及冷害等非生物胁迫,从而使得芜菁产量减少[2-3],因此研究并提高芜菁的抗逆性具有重要的意义。
脂氧合酶(lipoxygenase)是一种多功能酶[4],目前,在水稻[5]、马铃薯[6]、柿[7]、甜瓜[8]、谷子[9]等作物中对LOX基因功能均进行了相关研究,其广泛参与种子萌发、块根发育、果实软化、香气形成、色素积累等生理过程。同时LOX基因在植物响应逆境胁迫中也发挥着重要作用,研究发现,在干旱胁迫和高盐胁迫下,辣椒CaLOX1在拟南芥中过表达,增强了植株对干旱胁迫和高盐胁迫的耐受性[10]。干旱胁迫下,甜瓜CmLOX10基因在拟南芥中过表达,增强了植株耐旱性[11]。在高温胁迫和叶霉病侵染下,抑制番茄TomloxD基因的表达,降低了番茄耐热性并增加其对叶霉病的易感性[12]。然而关于芜菁BrrLOX基因在芜菁抗性相关内容尚未见报道。
本实验以芜菁为研究对象,从芜菁中克隆BrrLOX7基因,通过生物信息学方法分析BrrLOX7基因的理化性质、序列特征和进化关系,利用荧光定量PCR技术分析芜菁BrrLOX7基因在不同非生物胁迫下的表达模式,为探究该基因在非生物胁迫中的作用和功能奠定基础。
1 材料与方法
1.1 植物材料
以新疆天地禾种业有限公司生产的“卡玛古”种子为材料,将种子播种在湿滤纸上,在25 ℃黑暗条件下催芽,2 d后,将萌动的种子移栽到基质配比草炭∶蛭石∶珍珠岩体积比为2 ∶ 1 ∶ 1的塑料盆中后,放入培养箱中培养(光照周期为16 h光照/8 h黑暗,温度25 ℃白天/20 ℃夜晚,相对湿度为60%)。在幼苗生长到4片真叶时进行后续试验。
1.2 方法
1.2.1 总RNA的提取和cDNA第一链合成
采用Trizol法(总RNA抽提试剂,上海碧云天生物技术有限公司),按照说明书操作,提取不同处理的总RNA,利用1%的琼脂糖凝胶电泳检测RNA的完整性,使用反转录试剂盒(Quantscript RT Kit,北京天根生化科技有限公司)合成cDNA第一链,反应体系和程序参照说明书。
1.2.2 基因克隆
从BRAD数据库(http://brassicadb.cn/#/)下载芜菁基因组数据,从TAIR(https://www.arabidopsis.org/)下载拟南芥LOX基因序列,利用TBtools的BLAST工具筛选出位于7号染色体上的BrrLOX7基因序列。利用Primer Premier5.0软件参照同源基因序列设计特异性引物:
BrrLOX7-F:5′-ATGTTTTGTAAAGAGTCGTCG-3′;
BrrLOX7-R: 5′-TTAGATAGAGACACTGTAGGG-3′。
以芜菁cDNA为模板进行RT-PCR,利用TIANSeq高保真PCR反应预混液(TIANSeq HiFi Amplification Mix,北京天根生化科技有限公司)进行芜菁LOX基因全长编码区的扩增,PCR体系为50 μL的标准体系。PCR反应条件为:94 ℃ 2 min;98 ℃ 10 s,60 ℃ 30 s,68 ℃ 45 s,25个循环;68 ℃ 5 min。使用1%琼脂糖凝胶电泳检测PCR产物,利用DNA胶回收试剂盒对PCR产物进行回收,并将其连接至pMD19-T载体[pMD19-T Vector Cloning Kit,宝生物工程(大连)有限公司]上,转化大肠埃希菌DH5α后37 ℃培养12 h。挑取单菌落进行PCR验证,阳性克隆送至生工生物工程(上海)股份有限公司测序。
1.2.3 生物信息学分析
将芜菁BrrLOX7蛋白序列提交至NCBI(https://www.ncbi.nlm.nih.gov/),利用Blast搜索功能,获取不同物种的LOX蛋白序列,使用MUSCLE软件进行多序列比对,并使用MEGA-X通过邻接法构建系统发育树(重复1 000次),并使用iTOL(https://itol.embl.de/)进行美化;芜菁LOX蛋白序列使用Expassy ProtParam(https://web.expasy.org/protparam/)进行理化性质分析;使用SignalP 4.1(https://services.healthtech.dtu.dk/service.php?SignalP-4.1)进行信号肽分析;使用TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)进行跨膜结构分析;使用Expasy protscale(https://web.expasy.org/protscale/)进行亲疏水性进行预测;使用WoLF PSORT(https://wolfpsort.hgc.jp/)进行亚细胞定位预测;使用NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)查看结构域;使用SOPMA(https://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)进行二级结构分析;使用SWISS-MODEL(https://swissmodel.expasy.org/interactive)进行三维结构分析;使用Plant care(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对LOX启动子上游2 000 bp序列进行顺式作用元件的分析并利用TBtools进行可视化。
1.2.4 实时荧光定量PCR(qRT-PCR)分析
进行非生物胁迫处理时,将芜菁幼苗放置在4 ℃和40 ℃培养箱中模拟低温和高温,进行干旱胁迫和盐分胁迫处理时,幼苗叶片背腹面分别喷施20%的PEG6000和200 mmol·L-1的NaCl,以25 ℃常温栽培并只喷施纯水的芜菁幼苗为对照组,在处理后24 h采集芜菁幼苗叶片,迅速置于液氮中保存,每个处理3个生物重复。
根据已获得的目的基因序列,设计实时荧光定量PCR引物,BrrLOX7-qRTF:5′-GGCATCACAGTCAAATAGTGAACC-3′,BrrLOX7-qRTR:5′-CGAGACCTCTCGACCAACG-3′;以β-actin为内参基因,β-actinF:5′-CATGTTCGAGACGTTCAATG-3′,β-actinR:5′-GAACATGTAACCTCTCTCGG-3′。将芜菁不同处理的cDNA用ddH2O稀释20倍后作为模板,采用7500Fast实时荧光定量PCR仪检测BrrLOX7基因在不同处理中的表达情况,用内参基因检测扩增效率一致性,实时荧光定量PCR扩增体系参照说明书。
2 结果与分析
2.1 BrrLOX7基因的克隆
利用特异性引物扩增BrrLOX7基因的CDS区域,获得一条清晰明亮的条带(图1),与已知BrrLOX7基因CDS的大小相近,回收目的片段后连入克隆载体送生工生物工程(上海)股份有限公司进行测序。
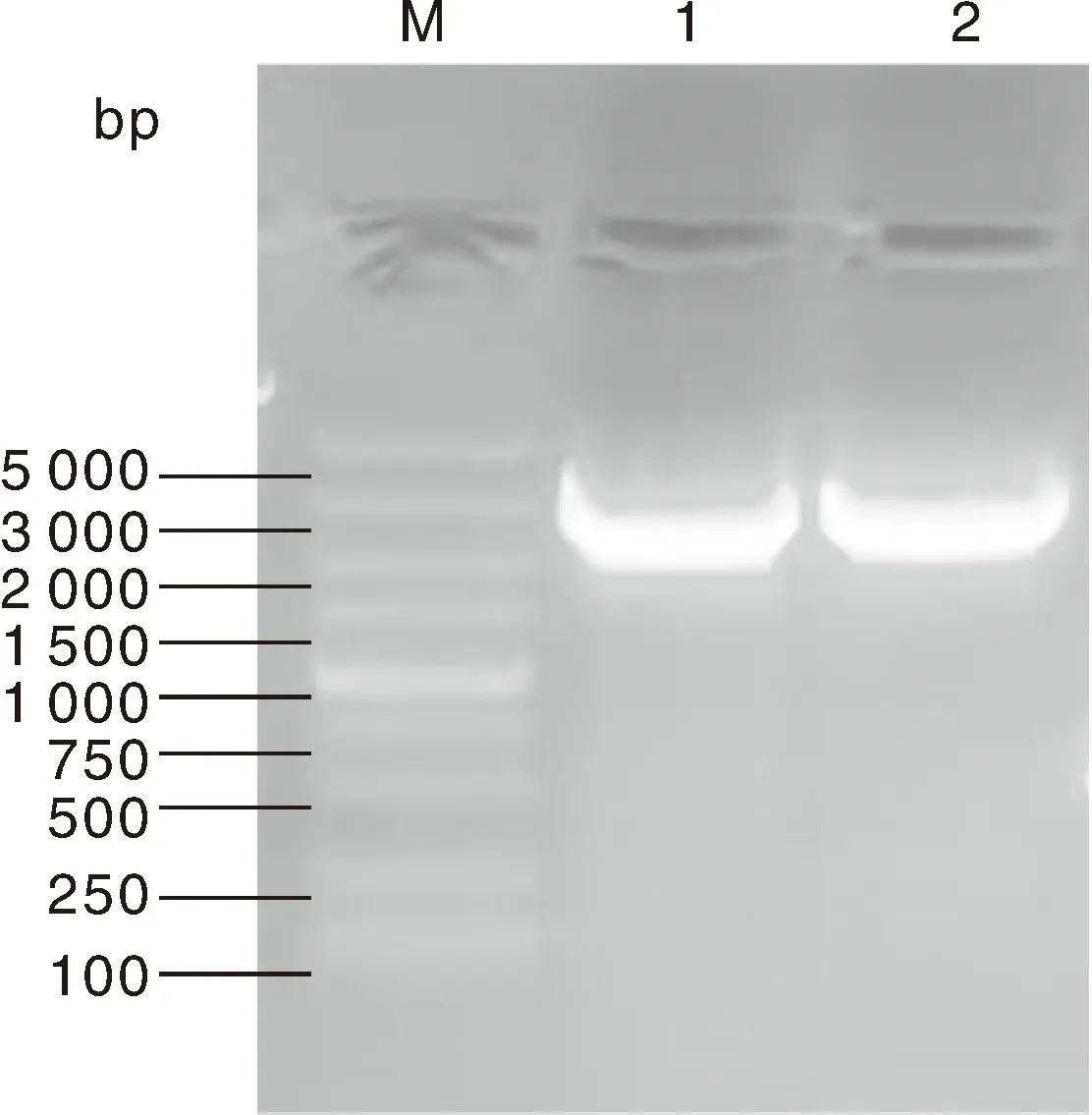
M,DL5000 marker;1和2表示BrrLOX7基因的PCR扩增产物。M, DL5000 marker; 1 and 2 indicate PCR amplification products of the BrrLOX7 gene.图1 BrrLOX7反转录PCRFig.1 RT-PCR of BrrLOX7
2.2 芜菁BrrLOX7基因的生物信息学分析
2.2.1 芜菁BrrLOX7氨基酸组成及理化性质
测序结果显示,芜菁BrrLOX7基因CDS序列长度为2 715 bp,共编码904个氨基酸,亮氨酸、谷氨酸、赖氨酸、脯氨酸的使用频率较高,分别占氨基酸总数的9.85%、8.30%、6.97%、6.86%(表1)。
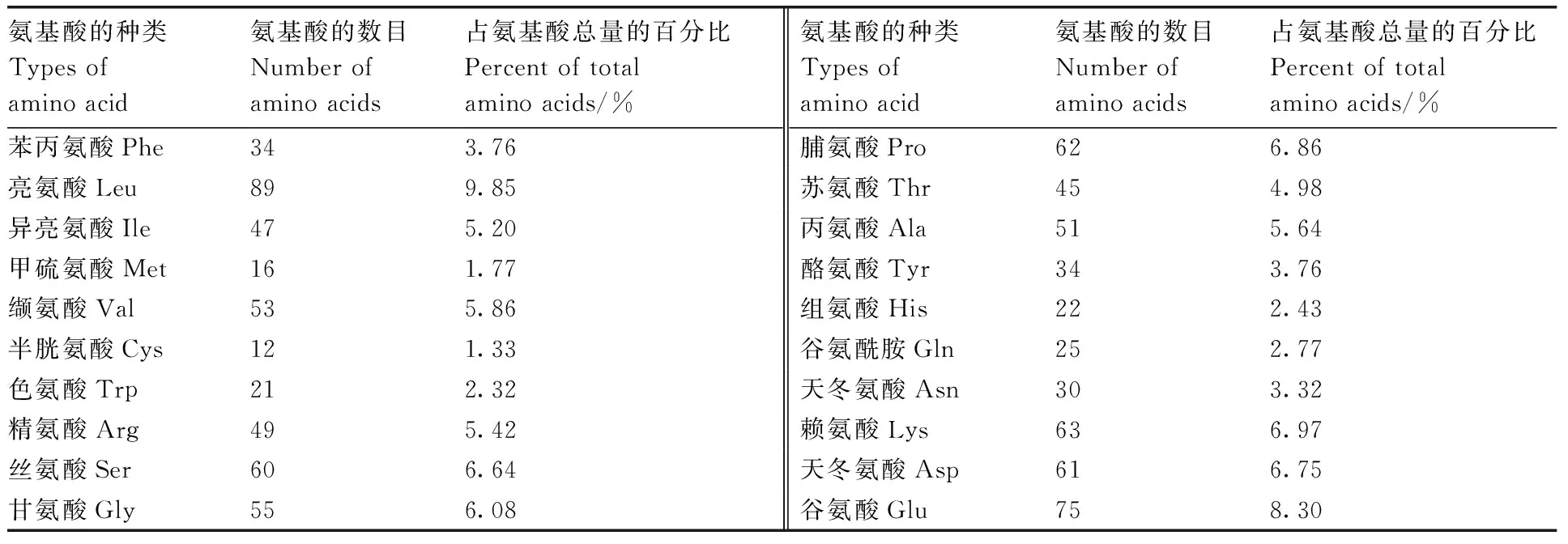
表1 BrrLOX7基因编码蛋白的氨基酸组成Table 1 Amino acid composition of the protein encoded by BrrLOX7 gene
芜菁BrrLOX7蛋白分子式为C4641H7178N1234O1371S28,分子量为103.10 ku,等电点为5.53,脂肪族指数为81.32,不稳定指数为45.92,属于不稳定蛋白,带负电荷的残基(Asp+Glu)为136,带正电荷的残基(Arg+Lys)为112,无信号肽,无跨膜结构。利用WoLF PSORT预测BrrLOX7蛋白的亚细胞定位,结果显示,BrrLOX7蛋白定位于叶绿体中。
2.2.2 BrrLOX7蛋白磷酸化位点和蛋白亲水性分析
蛋白磷酸化位点分析表明,BrrLOX7蛋白中含有41个丝氨酸磷酸化位点、24个苏氨酸磷酸化位点和16个酪氨酸磷酸化位点,第837位氨基酸的Score值最高为0.995(图2)。
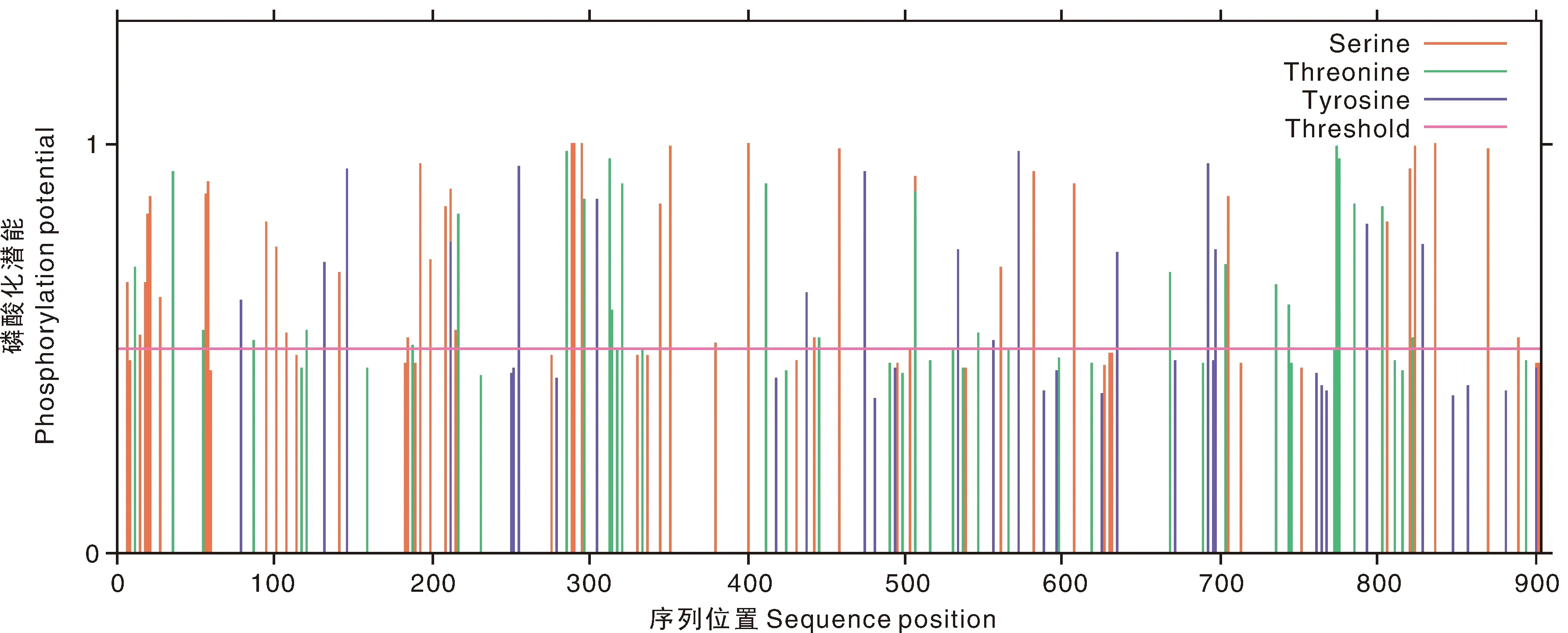
图2 BrrLOX7蛋白编码磷酸化位点预测Fig.2 Phosphorylation site prediction of BrrLOX7 protein
利用Expasy protscale软件分析BrrLOX7蛋白质的亲水性和疏水性,第328位的氨基酸残基的疏水性最强为2.256,第295位的氨基酸残基的亲水性最强为-2.922;另外,在该序列中,亲水性氨基酸累计有614个,疏水性氨基酸累计有280个,亲水性氨基酸的数目大于疏水性氨基酸的数目,因此,推测BrrLOX7蛋白属于亲水性蛋白(图3)。
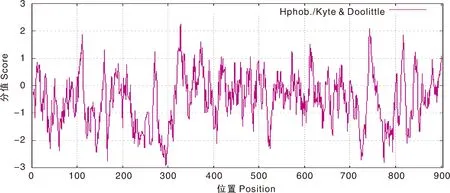
图3 BrrLOX7基因编码蛋白亲水性Fig.3 BrrLOX7 gene encodes a protein with hydrophilicity
2.2.3 蛋白的二级结构和三级结构预测
芜菁BrrLOX7蛋白二级结构主要由无规则卷曲、α-螺旋、延伸链和β-折叠组成,其中无规则卷曲(41.70%)和α-螺旋(40.27%)的占比最高,β-折叠(5.75%)占比最低(图4)。使用4wfo.1.A蛋白模型进行建模,芜菁BrrLOX7蛋白三级结构与模型的相似性为42.34%,模型覆盖率为91%(图5)。
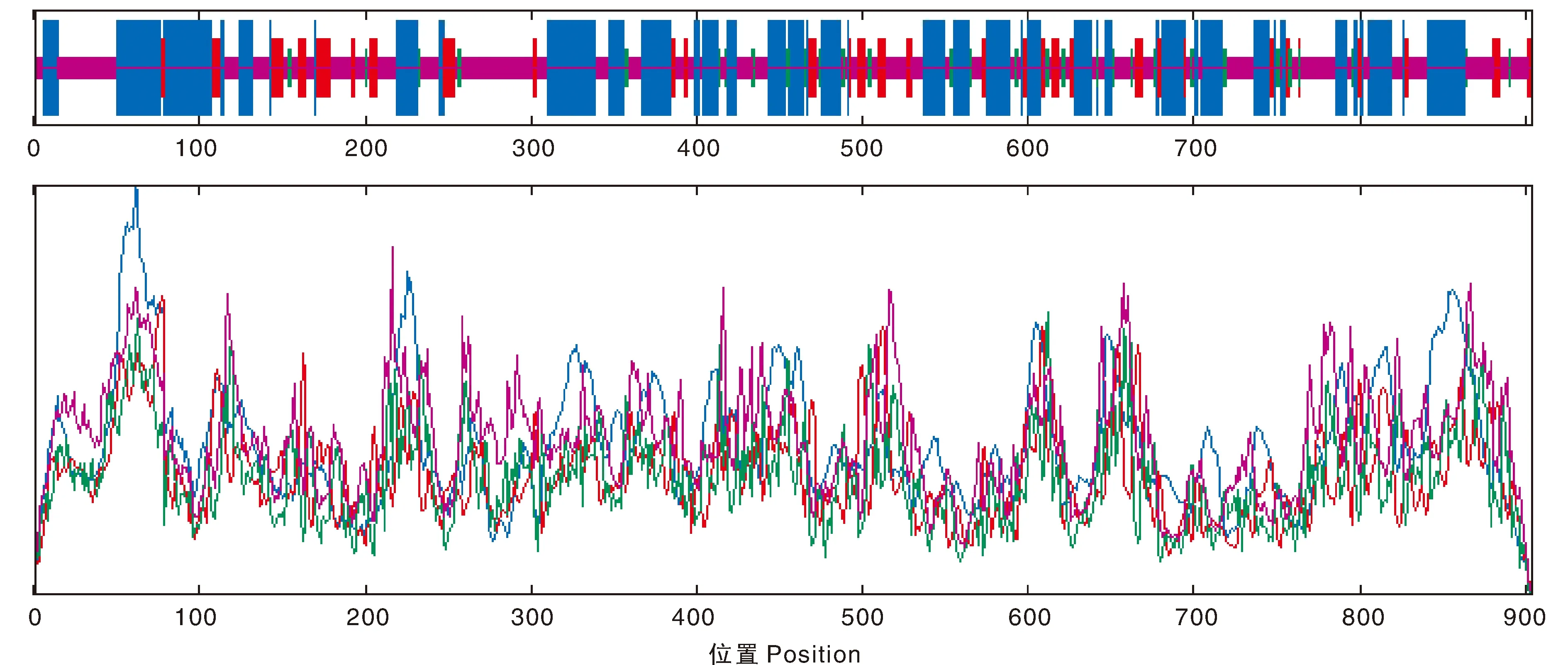
蓝色代表 α-螺旋; 红色代表延伸链; 绿色代表 β-折叠; 紫色代表无规则卷曲。Blue for α-helix; red for extended chain; green for β-turn; purple for irregularly curled.图4 BrrLOX7蛋白二级结构预测Fig.4 Secondary structure of BrrLOX7 protein
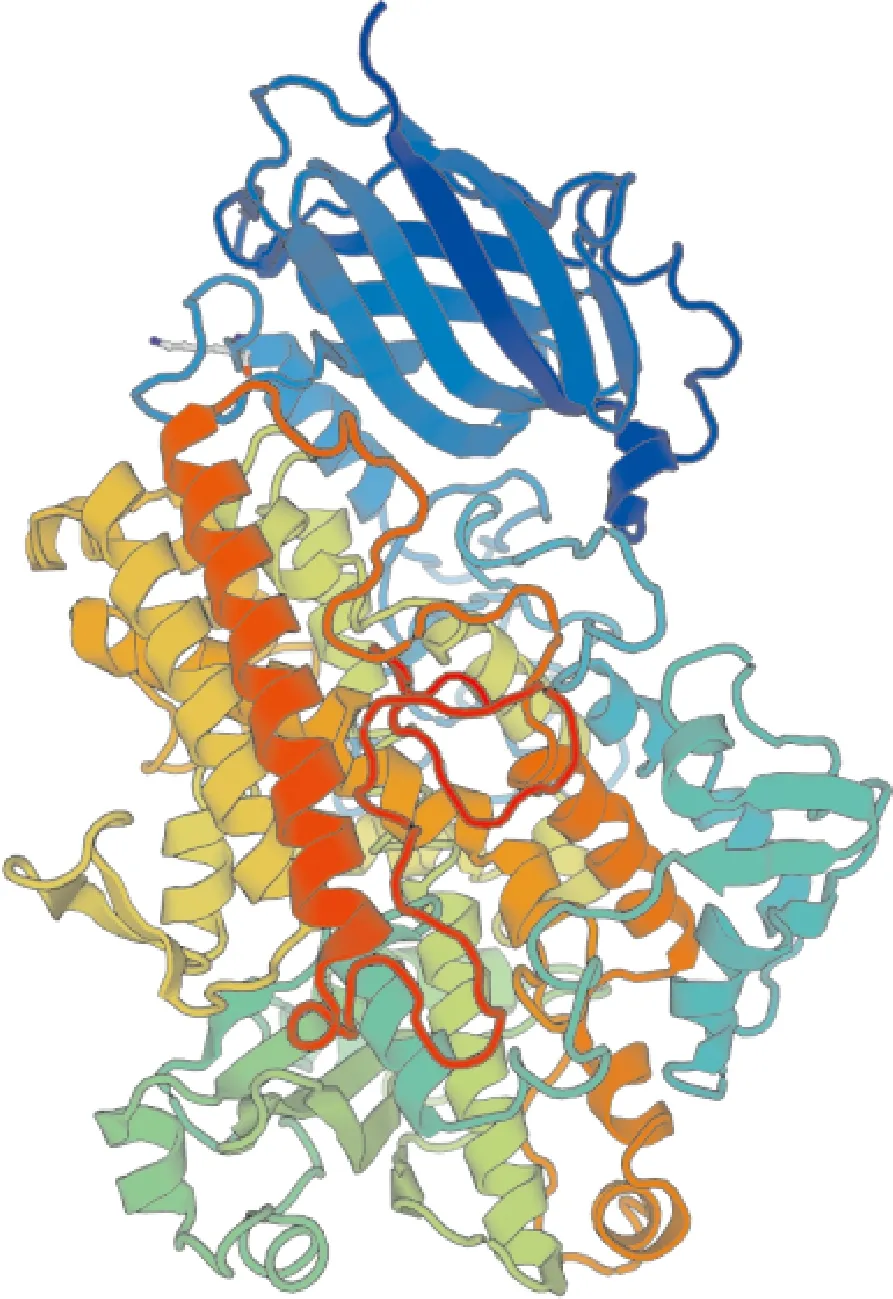
图5 BrrLOX7蛋白三级结构预测Fig.5 Tertiary structure of BrrLOX7 protein
2.2.4 BrrLOX7蛋白功能结构域预测
NCBI CDD预测显示,BrrLOX7蛋白序列包含2个保守结构域,PLAT_LH2结构域(位于第86~209位氨基酸)和Lipoxygenase结构域(位于第221~887位氨基酸)。PLAT_LH2结构域和Lipoxygenas结构域是LOX家族基因所特有的,进一步说明本试验克隆得到的基因为LOX基因(图6)。

图6 BrrLOX7蛋白序列结构域预测Fig.6 Structural domain prediction of the BrrLOX7 protein sequence
2.2.5 芜菁BrrLOX7蛋白的同源性比对
将芜菁BrrLOX7蛋白质序列提交至NCBI,利用BLASTP搜索功能,获取了11个物种的LOX蛋白序列,蛋白多序列比对结果表明,BrrLOX7与白菜(XP_009 104 416.3)、萝卜(XP_018 446 361.1)、盐芥(XP_024 016 495.1)、拟南芥(CAA0384539.1)4种同科物种的序列同源性分别为100%、91.59%、86.92%、81.01%,与桃(XP_007 220 253.2)、甜瓜(NP_001 315 400.1)、苹果(RXH94122.1)、黄瓜(XP_004 142 135.2)、辣椒(PHT59204.1)、马铃薯(NP_001 274 843.1)、葡萄(XP_034 705 038.1)7种非同科物种的序列同源性分别为63.02%、62.07%、62.07%、61.82%、61.14%、61.09%、56.84%(图7)。进一步利用MEGA-X构建BrrLOX7蛋白的系统发育进化树,其结果表明,BrrLOX7与芥菜亲缘关系最近,与桃和苹果亲缘关系最远(图8)。

图7 芜菁与其他植物BrrLOX7蛋白的氨基酸序列比对Fig.7 Comparison of amino acid sequence of BrrLOX7 protein in turnip and other plants
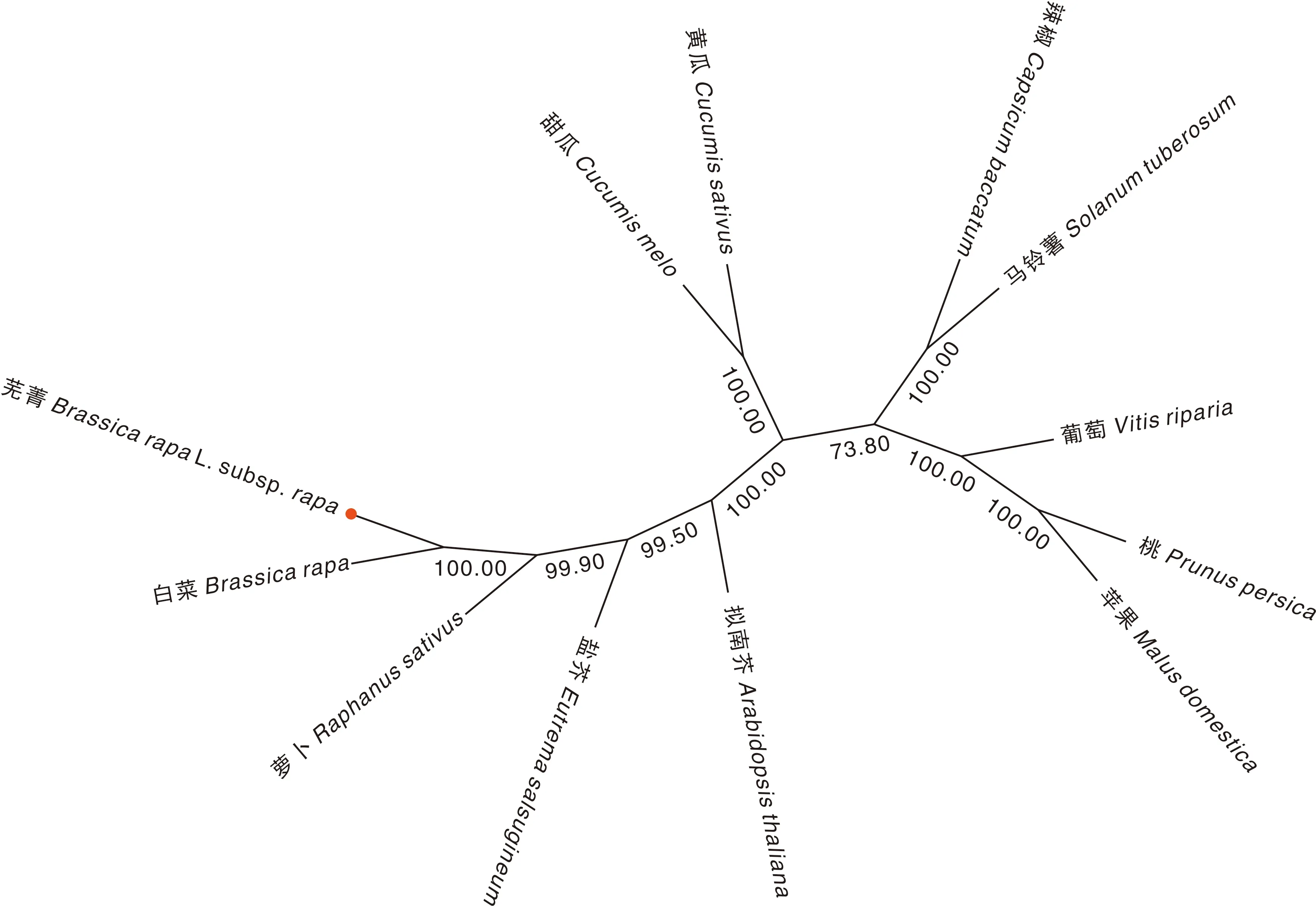
图8 芜菁BrrLOX7蛋白与其他植物相关蛋白序列的系统进化树Fig.8 Phylogenetic tree of turnip BrrLOX7 protein and other plant-related protein sequences
2.2.6 芜菁BrrLOX7基因启动子顺式作用元件预测
使用TBtools软件提取芜菁基因组中BrrLOX7上游2 000 bp启动子序列并提交至Plantcare在线网站预测可能的顺式作用元件(图9)。结果显示,在启动子中发现了13种(108个)顺式作用元件,除核心启动子元件TATA-box和CAAT-box外,还存在脱落酸诱导顺式作用元件ABRE,昼夜节律顺式作用元件Circadian,参与玉米醇溶蛋白代谢调节元件02-site,光响应元件TCT-motif、G-box、GATA-motif、Box 4,参与防御和应激反应的顺式作用元件TC-rich repeats,无氧诱导元件ARE和顺式调节元件A-box。
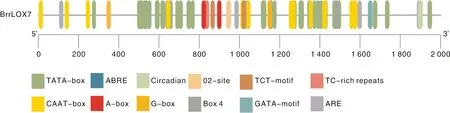
图9 芜菁BrrLOX7基因启动子区顺式作用元件预测Fig.9 Prediction of cis-acting elements of the BrrLOX7 gene promoter in turnip
2.2.7BrrLOX7基因在不同非生物胁迫下的差异表达
为检测BrrLOX7基因在4种逆境胁迫下的表达特性,以4叶期芜菁为材料,提取不同胁迫处理下的芜菁叶片总RNA,以反转录稀释后的cDNA为模板,进行qRT-PCR检测。结果如图10所示,BrrLOX7基因在4种胁迫处理下均有响应,但表达量存在一定差异,在高温处理下BrrLOX7基因表达量较高,盐胁迫和低温居中,在低温处理下表达量增加的最少。
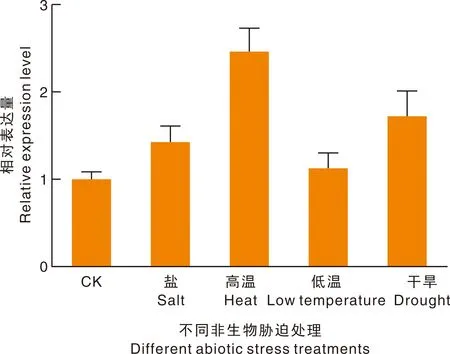
图10 芜菁BrrLOX7基因在非生物胁迫下的表达Fig.10 Expression of the BrrLOX7 gene in turnip under abiotic stress
3 讨论
本试验从芜菁中克隆得到BrrLOX7基因,该基因开放阅读框为2 715 bp,预测其分子量为103.10 ku,等电点为5.53,高于苹果MdLOX1a基因的分子量97.69 ku和等电点5.14[13],同时小于薄皮甜瓜CmLOX08基因的分子量104.93 ku和等电点8.44[14]。在BrrLOX7蛋白磷酸化位点的分析中,预测到41个丝氨酸磷酸化位点,24个苏氨酸磷酸化位点和16个酪氨酸磷酸化位点,这些磷酸化位点可能在调节蛋白质的结构和功能上发挥着重要作用[15]。预测的BrrLOX7蛋白无信号肽,不具有跨膜结构,说明该基因编码的蛋白不属于分泌蛋白,这一结果在小白杏PaLOX中也有发现[16],将BrrLOX7与11个物种的同源序列进行多序列比较和进化树分析发现,芜菁的BrrLOX7基因与芥菜和萝卜同科作物的遗传距离较近,说明该基因在种属进化中具有高度保守性。NCBI CDD的结果显示,BrrLOX7蛋白含有脂氧合酶家族特有的PLAT_LH2结构域和Lipoxygenas结构域,前人研究发现,PLAT_LH2结构域在酶的催化和膜脂结合能力中发挥作用[17],Lipoxygenas结构域中,由组氨酸(His)组成的高度保守的38氨基酸残基区域[His-(X)4-His-(X)4-His-(X)17-His-(X)8-His]在LOX与铁离子结合发挥酶稳定性和活性中发挥作用[18]。我们对BrrLOX7基因进行了亚细胞定位预测,结果显示,BrrLOX7基因主要定位于叶绿体。这一结果与拟南芥的AtLOX2[19],马铃薯LOXH1和LOXH3[20],甜瓜的CmLOX13蛋白的亚细胞定位结果一致[21],因此,BrrLOX7基因可能在叶绿体中发挥着重要作用。
Ju等[22]利用Plant CARE软件在甜瓜CmLOX09启动子中预测到激素响应元件、应激响应元件和信号类物质,并以此为突破口,探究了CmLOX09基因对逆境、激素和信号类物质的响应,结果表明,CmLOX09基因通过氢过氧化物裂解酶(HPL)途径产生的绿叶挥发物(GLV)来响MeJA,并通过丙二烯合酶(AOS)途径产生的茉莉酸来响应真菌胁迫。在本研究中,通过对BrrLOX7基因启动子预测发现了脱落酸响应元件和防御胁迫应答元件,推测BrrLOX7基因可能通过调控脱落酸等植物激素参与了植物逆境胁迫响应。这为今后继续深入研究芜菁BrrLOX7基因的表达调控及防御胁迫响应机制奠定了基础。
植物在生长过程中,时常会受到干旱和盐害等非生物胁迫的影响,导致作物的生长发育受阻,因此植物需通过响应并适应胁迫而正常生长,当植物受到非生物胁迫时,通过诱导脂氧合酶基因的表达提高植物抗性已有许多文献报道。研究发现,在高温处理下,番茄TomLOXC基因超表达植株通过JA积累来调节植株的抗性[23]。在干旱胁迫和盐胁迫处理下,甜瓜CmLOX08基因通过提高植株的LOX活性、胁迫基因表达水平和清除活性氧能力,增强了植物的抗旱和抗盐性[14]。在干旱胁迫下,甜瓜CmLOX13在拟南芥中过表达,通过降低分解代谢相关基因的表达和ABA诱导的气孔关闭,提高了抗旱能力[21]。在本研究中,BrrLOX7基因在NaCl、高温、低温、干旱中均有应答,暗示该基因可能在芜菁抗逆生长中扮演重要的角色。
本研究从芜菁中克隆得到了BrrLOX7基因序列,对其进行了生物信息学分析和表达特性分析,初步确定BrrLOX7基因在芜菁响应不同非生物胁迫中发挥功能,为后续深入探究BrrLOX7基因在芜菁中的功能奠定了基础。
猜你喜欢
蔬菜(2022年2期)2022-02-17
电脑迷(2020年7期)2020-08-07
天津医科大学学报(2019年6期)2019-08-13
广州大学学报(自然科学版)(2019年1期)2019-05-07
新疆农业科学(2018年8期)2018-10-23
分析化学(2017年12期)2017-12-25
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
安徽医科大学学报(2015年9期)2015-12-16
现代检验医学杂志(2015年2期)2015-02-06
