
新疆野核桃自然保护区不同坡向丛生野核桃年龄结构及数量动态
2023-04-11 03:34:44魏海英黎一阳赖晓辉张维
热带亚热带植物学报 2023年2期
魏海英, 黎一阳, 赖晓辉, 张维*
新疆野核桃自然保护区不同坡向丛生野核桃年龄结构及数量动态
魏海英1, 黎一阳2, 赖晓辉1, 张维1*
(1. 伊犁师范大学生物与地理科学学院,资源与生态研究所,新疆 伊宁 835000; 2. 第四师伊犁职业技术学校,新疆 伊宁 835000)
通过对新疆野核桃自然保护区丛生野核桃()进行普查,以径级结构代替年龄结构,采用匀滑技术编制种群静态生命表,并运用时间序列模型预测种群数量动态,从而获得丛生野核桃树的龄级结构与动态。结果表明,不同坡向丛生野核桃以中龄级为主,年龄结构呈两头小中间大的纺锤型;4个坡向种群年龄结构的动态指数V虽大于0,但最大仅为19.10%,且Vʹ极低,最大为1.12%,阳坡种群增长趋势最高,抗干扰能力最强,种群稳定性最好,半阳坡种群增长趋势最弱,抗干扰能力最弱,种群稳定性最差;各坡向种群均存在多个死亡高峰,阳坡最高峰在15龄级,半阳坡在15和18龄级,阴坡和半阴坡均在9龄级,存活曲线更接近Deevey-Ⅱ型;在未来2、5、10龄级后,中龄级个体数量减少,老龄个体呈增加趋势,丛生野核桃种群稳定性难以长期维持。因此对丛生野核桃幼苗的保护非常关键。
野核桃; 静态生命表; 龄级结构; 种群动态
种群数量动态是某一种群个体生存能力和生存环境相互作用的结果,是用来研究种群密度或规模在空间、时间上的变化规律,对阐述濒危植物的保护和利用具有重要意义[1–3]。年龄结构、种群动态指数、种群生命表、存活曲线、时间序列分析模型是研究种群数量动态常用的方法。其中年龄结构决定了种群的繁殖能力和发展趋势[4];种群动态指数既可弥补在种群结构比较评价中等级归类划分的不足,又可解决对群落结构动态进行定量比较评价的问题[5];种群生命表及其所构建的存活曲线既可以反映生存现状,又可以反映植物与外界环境的适合度[6–7];时间序列分析模型能预测同一年龄级种群未来波动变化趋势[8]。因此通过这些方法可以比较科学的评估种群数量动态, 了解种群生存现状,揭示种群生存潜力及致危原因。
野核桃()为胡桃科(Juglandaceae)胡桃属的高大落叶乔木,国家Ⅱ级渐危物种,是珍贵的第三纪温带落叶阔叶林的残遗成分,是新疆野核桃自然保护区的建群物种之一。在亚洲,野核桃仅在巩留县新疆野核桃自然保护区成片分布,霍城县的大、小西沟内零星分布[9]。它对研究栽培核桃的起源、演化,古植物区系的变迁,第三、第四纪气候变化具有特殊重要的科学和生态价值,对保护伊犁野果林生物多样性、维护和改善区域生态平衡起至关重要的作用[10]。前人对野核桃的研究主要集中在种质资源分布[9–11]、生物及生态学特征[9–10]、繁殖生物学[11–13]、种群生态学[14–15]、保护遗传学[16–17]等方面。目前关于丛生野核桃种群结构及数量动态尚未见报道。本研究以野核桃自然保护区4个坡向丛生野核桃为研究对象,对其年龄结构、种群数量动态特征进行定量统计分析,并预测种群未来发展动态趋势,以期对保护区的科学管理和该物种的保护提供科学依据。
1 研究区概况
新疆野核桃自然保护区位于伊犁哈萨克自治州巩留县城南伊什格力克山的峡谷中(43°22′56″~ 43°25′40″ N,82°15′28″~82°17′23″ E),保护区东、南、北由大山环抱,向西敞开,能有效减弱冷空气和寒流的侵袭,而有利于黑海湿气和巴尔喀什暖流进入[9,15]。保护区总面积1 180 hm2,其中野核桃分布约44 hm2;野核桃最低分布海拔1 241 m,最高分布海拔1 670 m[9–10]。年均温7.6 ℃,7月均温19.7 ℃,1月均温–3.3 ℃,无霜期150 d,年均降水量580 mm,年均空气相对湿度70%~80%[12–14]。保护区内覆盖深厚的第四纪黄土层,按照海拔高度依次发育了高山草地土、亚高山草地土、山地灰褐色森林土、山地黑钙土、山地栗钙土[10,18–19]。保护区植物资源丰富,维管束植物有56科182属329种, 植被种类以温带植物为主, 热带植物为辅;被子植物有47科171属303种[10]。野核桃种群伴生的主要乔木树种有野杏()、新疆野苹果()、天山桦()等;灌木主要有阿尔泰山楂()、金丝桃叶绣线菊()、异果小檗()等;草本植物主要有短柄草()、竹节菜()、大羊茅()等[15]。
2 研究方法
2.1 野外调查
2019年6月,在野核桃自然保护区的阳坡、半阳坡、阴坡、半阴坡(坡向划分参考陈晨[20]的方法)采用普查法对丛生野核桃(同一树桩可萌生幼苗并发育2~6株丛生)进行调查,记录其丛生野核桃的株树(同一树桩仅形成2个主干即为2株丛生,同一树桩形成3个主干即为3株丛生,以此类推)、组数(同一根系为1组)、胸径(幼苗测量基径)、树高(幼苗测量株高)、坡向等指标。
2.2 种群结构划分
参考张金屯等[21]的方法将丛生野核桃种群胸径划分为19个径级,第I径级为0~2.5 cm,之后每隔5 cm设1径级,即第II径级为2.5~7.5 cm,第III径级为7.5~12.5 cm,以此类推。同种生存环境下同一个物种的生长规律具有一致性[6,15],因此将径级按从小到大的顺序认定为时间顺序,即第I径级对应1龄级,第II径级对应2龄级,以此类推。根据野核桃生长特点,将1龄个体(DBH<2.5 cm)划分为幼苗,2龄个体(2.5 cm≤DBH<7.5)为小树,3~7龄个体(7.5 cm≤DBH<32.5 cm)为中龄树; 8~19龄个体(DBH≥32.5 cm)为老龄树[22]。
2.3 种群年龄结构数量变化动态指标分析

2.4 静态生命表编制
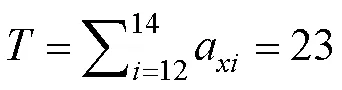
为了选择最符合种群存活状况的存活曲线,本文采用数学统计模型的显著性检验,即比较指数方程y=ae和幂函数y=ax,若指数方程拟合效果好,存活曲线趋于Deevey-Ⅱ型;若幂函数拟合效果好,存活曲线趋于Deevey-Ⅲ型[25]。
2.5 种群数量动态的时间序列预测模型
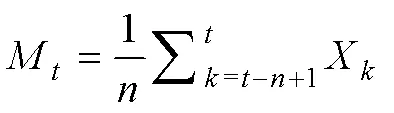
3 结果和分析
3.1 种群的分布及年龄结构
同一野核桃树桩可发育2~6株丛生,不同坡向丛生野核桃均以2株丛生最多,其中阴坡占比重最大,为65.31%,半阴坡次之,阳坡最少;其次为3株丛生,阳坡>半阳坡>半阴坡>阴坡;6株丛生最少,半阳坡、阴坡、半阴坡分别为3.17%、2.04%、3.18%,阳坡既无5株丛生,也无6株丛生(图1)。保护区丛生野核桃共348组921株,其中阳坡16组43株,半阳坡126组339株,阴坡49组128株,半阴坡157组411株,即阳坡和阴坡丛生野核桃的丛数、植株数均小于半阳坡、半阴坡,表明丛生野核桃更喜半阳坡、半阴坡的微生境。
根据实地调查不同坡向丛生野核桃径级数据,得到丛生野核桃种群的年龄结构(图2)。不同坡向丛生野核桃年龄结构均不完整,具体表现为:阳坡缺1龄级、12~16龄级个体,半阳坡缺1龄级个体,阴坡缺1、12、13、15龄级个体,半阴坡缺17龄级个体,表明丛生野核桃在其生长发育过程中是不连续的。4个坡向的龄级以半阳坡最大,为19龄级, 其次为半阴坡18龄级,再次为阳坡17龄级,阴坡最小,为16龄级。不同坡向上幼龄树(1~2龄级)个体数量极少,其中阴坡总占比最大,为7.03%,其次为半阳坡,阳坡最少,为2.32%,这可能与丛生野核桃萌枝能力弱和林下郁闭度强有关。笔者调查时发现,保护区内极少见到野核桃在其根茎部位萌生幼苗(DBH<2.5 cm),大部分都是由野核桃实生苗在幼龄时期受到低温或倒春寒的影响形成了分杈的多主干树形。不同坡向中龄级(3~7龄级)个体数量最多,且均在54.00%以上,其中阴坡最多为68.75%,其次为阳坡,半阳坡最少为54.57%。老龄级(8~19龄级)个体数量急剧减少,总占比介于24.22%~39.23%,表明不同坡向中老龄级丛生野核桃种群受自然干扰和人为干扰较大,导致丛生野核桃大量死亡。由此可知4个坡向上各龄级的数量呈现为两头小,中间大的纺锤形。
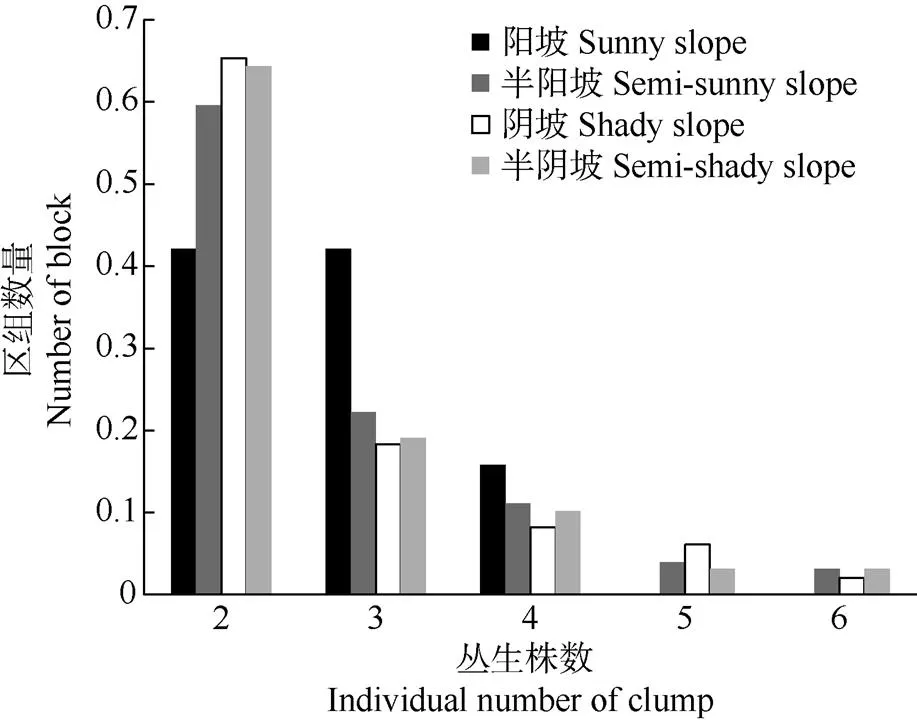
图1 不同坡向丛生野核桃的分布及株数
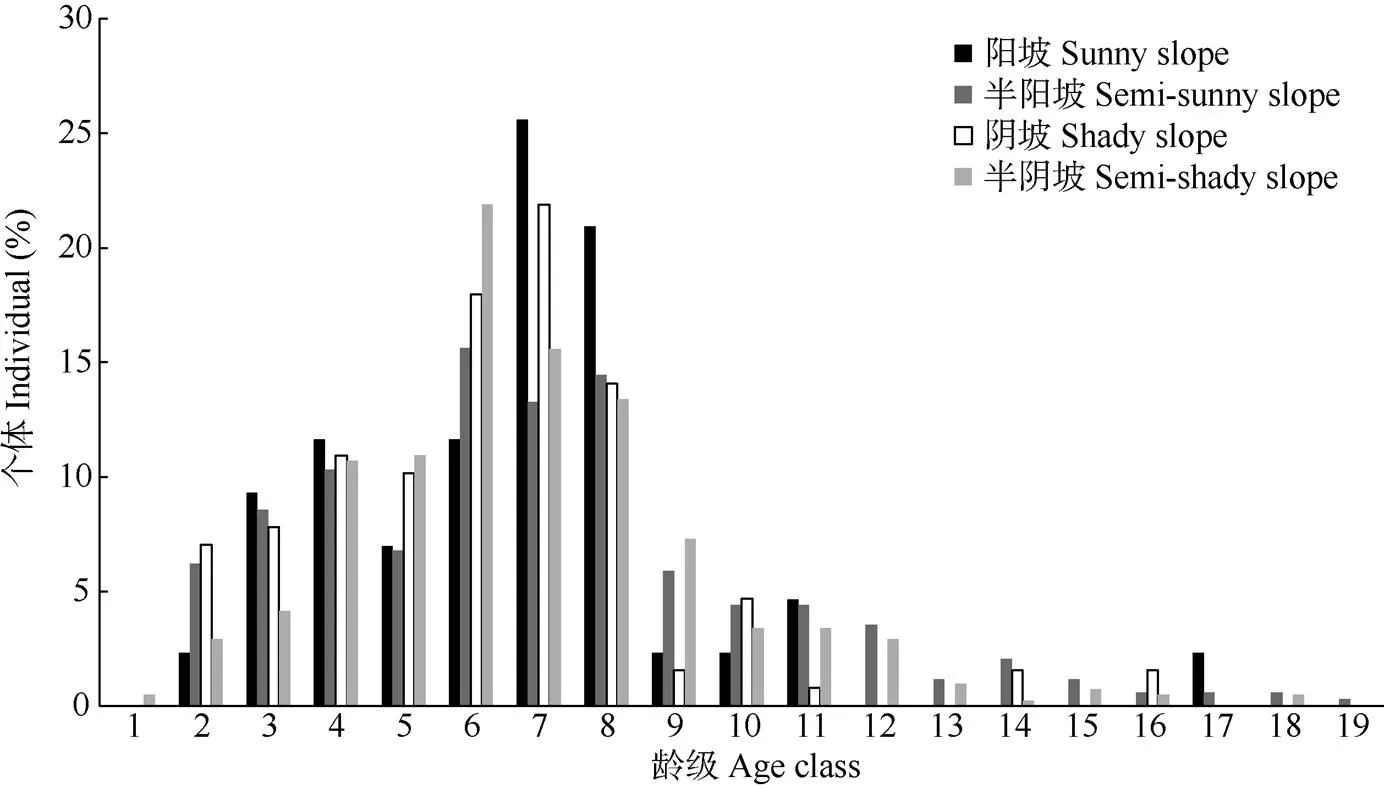
图2 不同坡向丛生野核桃种群年龄结构
3.2 种群的动态
丛生野核桃种群相邻年龄级间个体数量整体上呈“衰退-增长-衰退”波动的结构动态关系(表1)。4个坡向丛生野核桃种群1~3均为负值,说明丛生野核桃种群在1~3龄级经历强烈的环境筛,呈衰退型结构,但只要通过此阶段环境筛的筛选,大多数植株可生长发育为成龄树,直至完成整个生活史。4个坡向,阳坡种群不考虑外部干扰,种群年龄结构动态指数V值最大,为19.10%,增长趋势最高,考虑外部干扰时种群年龄结构动态指数Vʹ值也最大,为1.12%,显示种群最强的抗干扰能力,种群稳定性最好,与阳坡相反,半阳坡种群表现出增长势最低,种群抗干扰能力最弱,种群稳定性最差,阴坡、半阴坡种群抗干扰能力和增长趋势介于阳坡和半阳坡之间。不同坡向丛生野核桃种群数量动态指数虽然均大于0,4个种群均呈增长型结构,但Vʹ值极低,显示4个坡向均有衰退的趋势,与野外调查结果一致。统计表明阳坡、半阳坡、阴坡、半阴坡种群最大随机干扰风险值max最大,仅相差0.010,差异不显著(>0.05)。
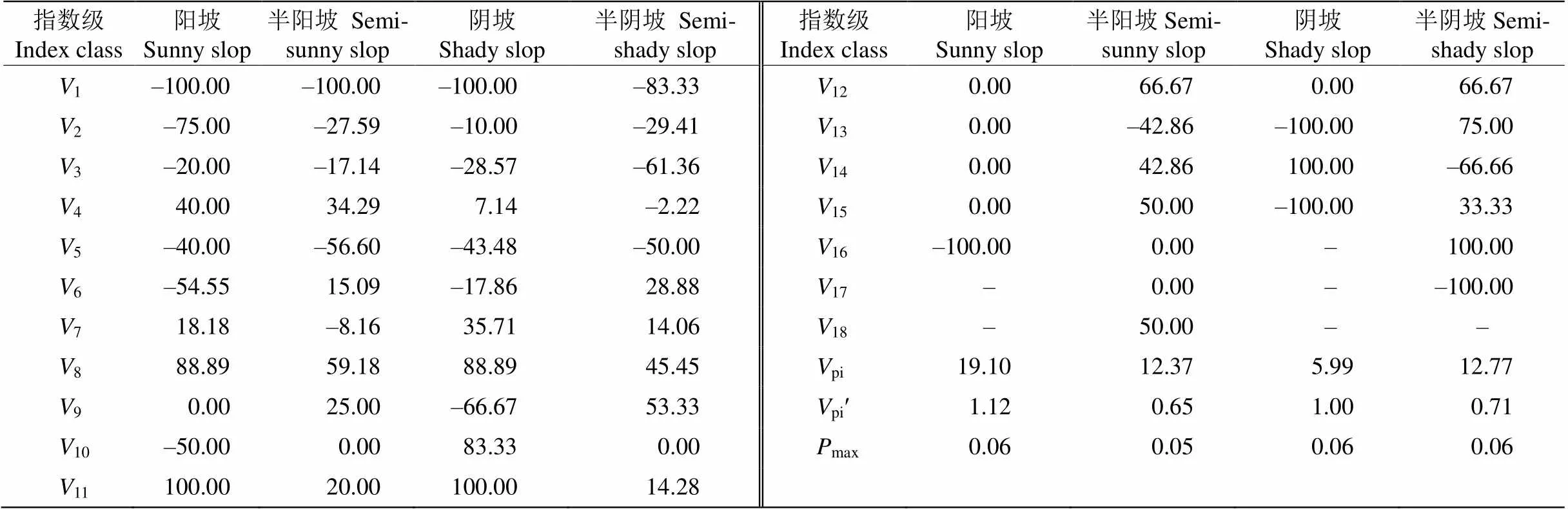
表1 不同坡向丛生野核桃种群年龄结构动态变化指数(%)
3.3种群的静态生命表、存活曲线及死亡曲线
静态生命表显示,不同坡向上丛生野核桃种群随龄级增加,存活个体数a先增加后减少,但个别龄级存在波动(表2)。4个坡向上丛生野核桃期望寿命随龄级的增加而呈现降低趋势,但同一坡向不同龄级(尤其老龄树)e存在较大波动,其中1龄级的期望寿命最大,究其原因主要与丛生野核桃幼苗可利用空间大,个体间竞争较小、生命力较强等因素有关。进入中龄级后,个体生长迅速,生存空间变大,从而引起种间、种内竞争导致期望寿命持续性下降。
以各龄级的死亡率q为纵坐标,以龄级为横坐标绘制丛生野核桃种群的死亡曲线(图3)。4个坡向种群死亡率均呈现“增大-减少-增大”的波动趋势。阳坡种群存在7和15龄级2个死亡高峰,死亡最高峰在15龄级;半阳坡存在1、5、9、11、15、18龄级6个死亡高峰,死亡最高峰在15和18龄级; 阴坡种群存在2、5、9、11、13、15龄级6个死亡高峰,死亡最高峰在9龄级;半阴坡种群存在6、9、11、14龄级4个死亡高峰,死亡最高峰在9龄级。各坡向丛生野核桃种群均存在多个死亡高峰,且死亡高峰的龄级不同,揭示不同坡向丛生野核桃种群在不同龄级经历环境筛的强弱不同,且微生境对丛生野核桃种群的维持和发展有重要的影响。
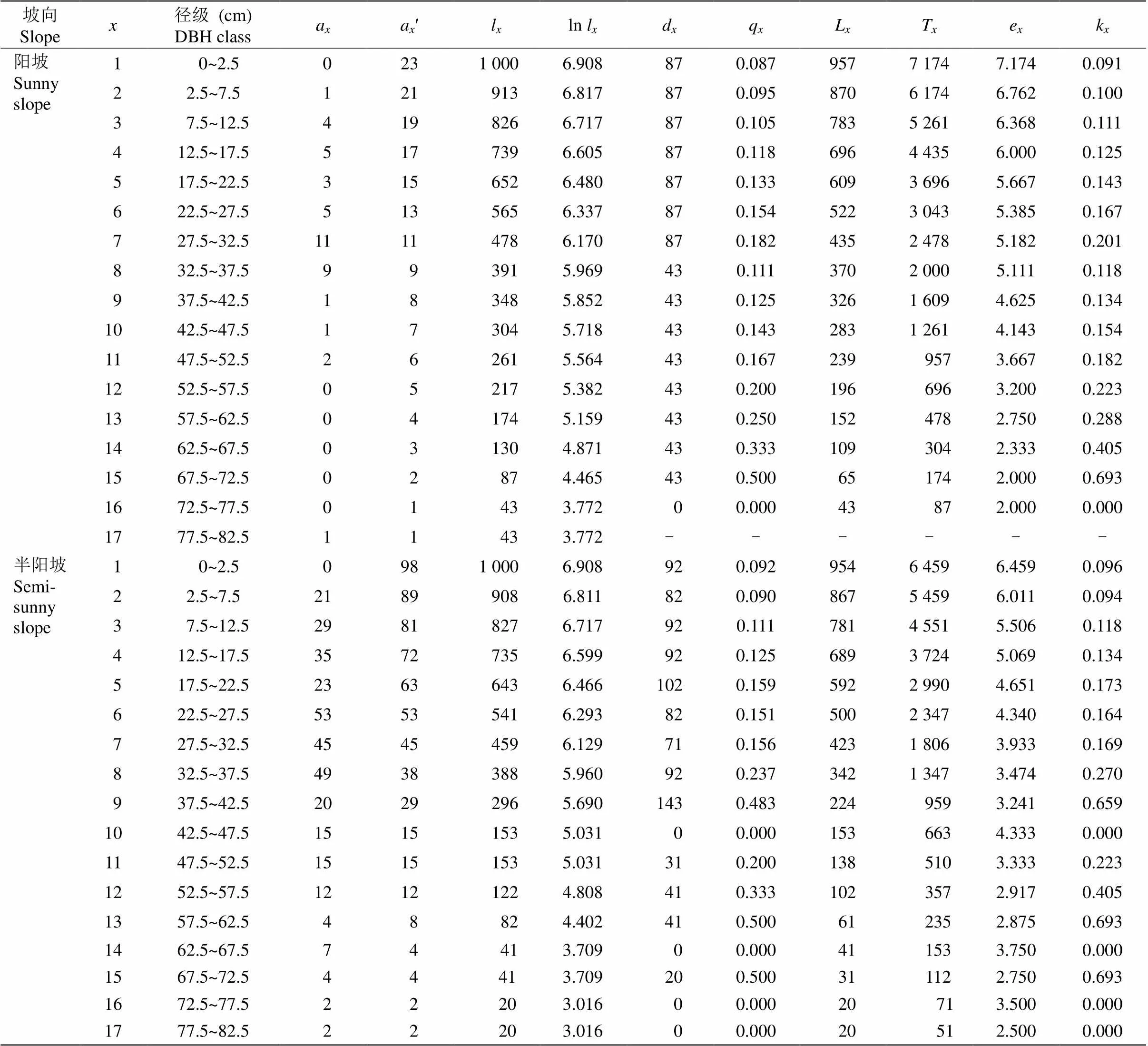
表2 不同坡向丛生野核桃种群的静态生命表
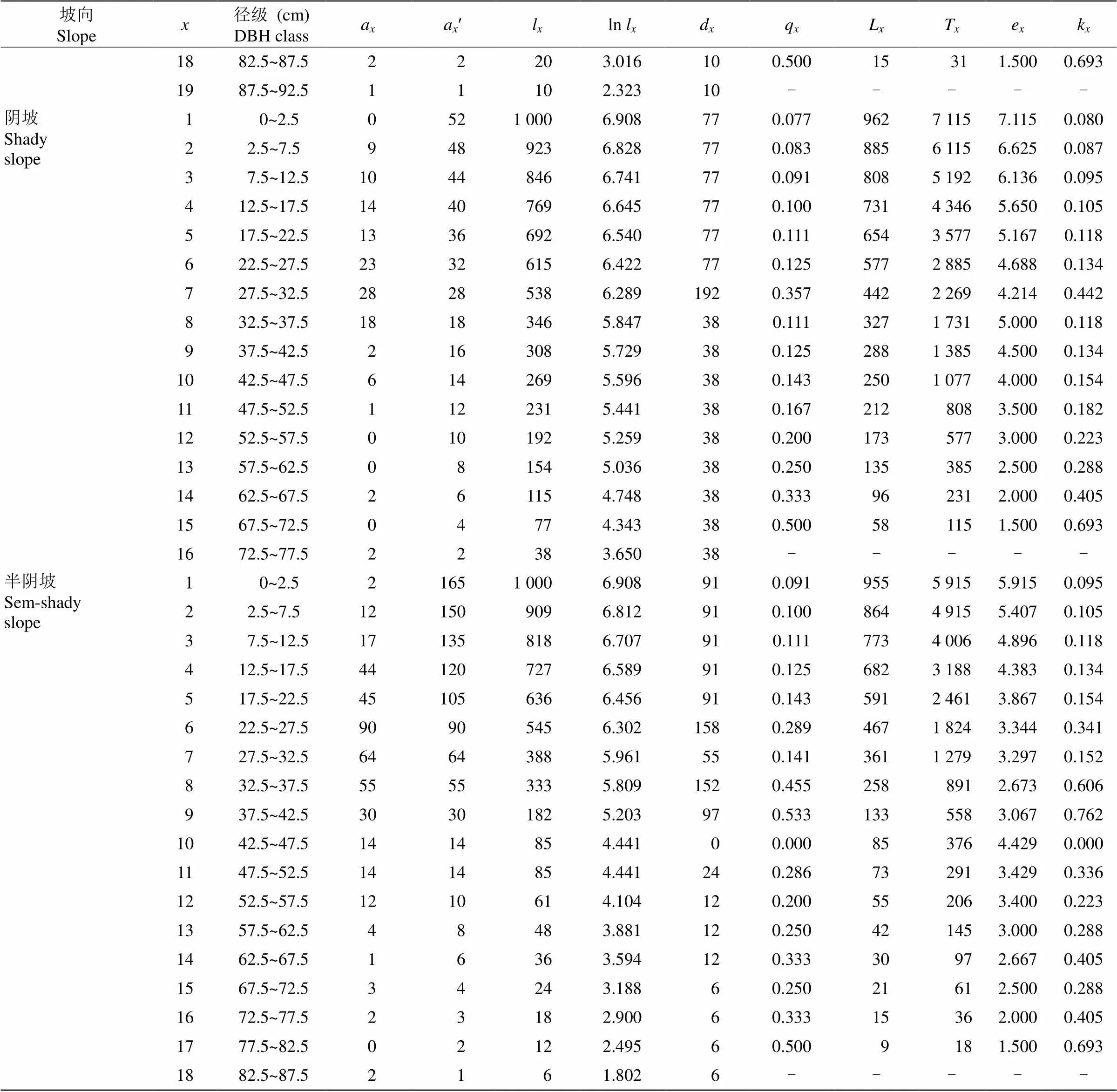
续表(Continued)
: 龄级;a:龄级个体数;aʹ: 匀滑修正后龄级个体数;l: 在龄级开始时标准化存活个体数;d:到+1龄级标准化死亡个体数;q: 从到+1龄级的死亡率;L: 从到+1龄级还存活的个体数;T: 从龄级到超过龄级的总个体数;e: 进入龄级个体的期望寿命;k: 消失率。
: Age class;a: Number of alive organisms at age;aʹ: Revised date ofa;l: Standard survival individuals number;d: Standardized number of death individuals from age classto+1;q: Mortality from age classto+1;L: Number of surviving individuals from age classto+1;T: Total individual number of age classand age classes older than;e: Life expectancy of individuals in the age class;k: Disappearance rate.
以各龄级存活量的对数lnl为纵坐标,以龄级为横坐标绘制丛生野核桃种群存活曲线(图4)。4个坡向均显示随龄级的增加存活率逐渐下降。分别用幂函数、指数函数2种数学模型对4个坡向的丛生野核桃种群存活量进行检验(表3)。两种模型的拟合在各坡向均达到极显著水平(<0.01),指数模型2、值均大于幂函数模型,说明4个坡向丛生野核桃种群存活曲线均趋向Deevey-Ⅱ型。
3.4 种群数量动态的时间序列预测模型
以丛生野核桃种群各龄级的植株数为基准,按照一次平均推移法预测各年龄级在未来2、5、10龄级时间后的植株数(图5)。4个坡向丛生野核桃种群在未来2、5、10龄级后的个体数量不同,其中阳坡丛生野核桃种群个体数量的峰值随时间序列依次向后推移;半阳坡与半阴坡种群动态相似,2~8龄级个体数量减少,9龄级及其以后个体数量明显增加,老龄树数量持续增加,而幼龄级、中龄级个体数量逐渐减少;阴坡丛生野核桃种群在未来2、5、10龄级老龄级个体数量比原始种群数量分别增加了41.94%、135.48%、535.45%。因此,4个坡向丛生野核桃种群由于幼龄个体严重匮乏,不能及时补充中龄级个体,如若长期保持这种趋势会增加丛生野核桃种群出现龄级断层,自然更新受阻的可能性。
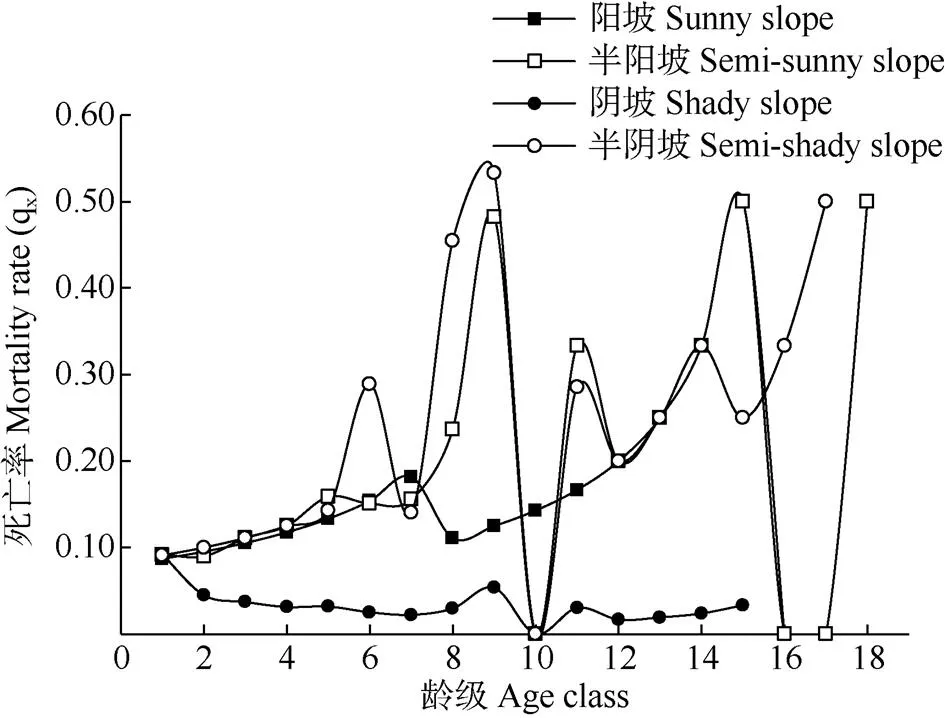
图3 不同坡向丛生野核桃种群的死亡率曲线
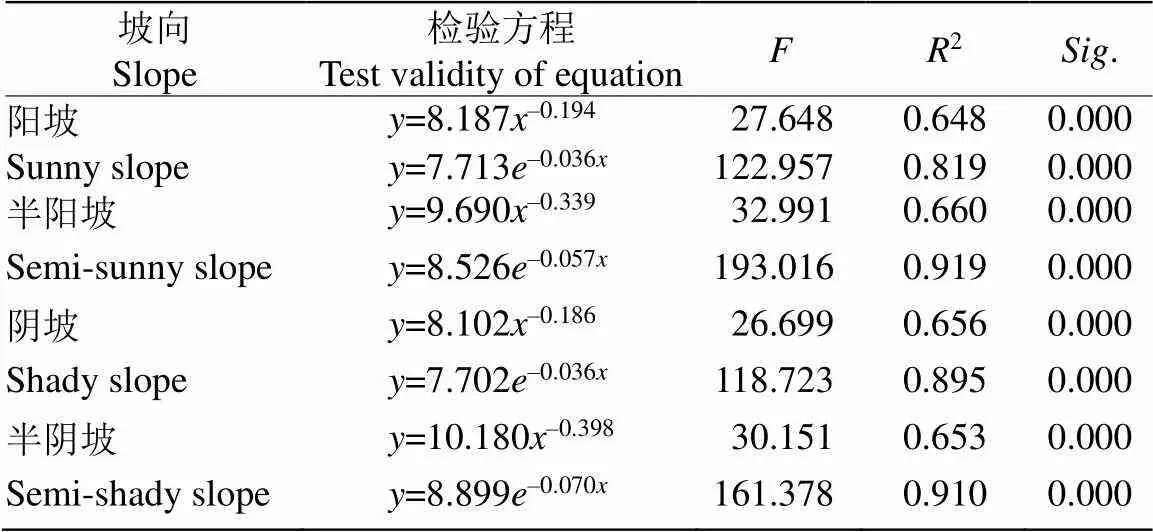
表3 不同坡向丛生野核桃种群存活曲线的检验模型
4 结论和讨论
4.1 野核桃种群的维持策略
在森林生态系统中,部分乔木树种的更新必须依靠种子发育的实生苗,另有一些乔木树种的更新虽然主要是种子萌发的实生苗,但其树桩上的萌生苗对种群的维持和发展也有不小的贡献[27]。张钊等[28]报道野核桃无根蘖繁殖,依靠自然落种发育为成龄树,曾斌[9]报道野核桃在幼龄级时易受低温和春寒的影响形成分杈的多主干树形。根据作者野外实地调查,丛生野核桃一方面是野核桃一个主干在树桩基部分杈形成多主干,另一方面是成龄野核桃树树桩基部萌生幼苗并生长发育成小树甚至成龄树,确实非实生苗。野核桃出现丛生化现象,可能与土壤缺乏B元素、生物病原侵害(如褐斑病、叶螨病等)、土壤砂粘比等多种因素有关。保护区土壤剖面以细沙、粉砂为主[18],较高的温度不仅促进了野核桃蒸腾作用加强,还加快了土壤水分蒸发,这使得野核桃根系从原本持水能力弱且养分含量少的土壤摄取的水分、溶解性养分更少,当根系吸收的水分和养分满足不了野核桃幼苗生长,野核桃新梢便会枯死,来年休眠芽萌发。由此可知野核桃新梢枯死是野核桃丛生化的直接原因之一。张维等[14]2015年调查统计野核桃为5 500余株,丛生野核桃约占野核桃种群总株数的16.75%,其对野核桃种群的延续有重要贡献,需持续关注丛生野核桃种群的发展动态。
4.2 丛生野核桃种群的分布差异
4个坡向丛生野核桃株数分布差异较大,半阳坡和半阴坡分别占36.81%和44.63%,阳坡和阴坡分别仅占4.66%和13.90%,株数占比差异可能受新疆野核桃的生物学特性控制,新疆野核桃喜温暖、湿润的环境[9,15],阳坡光照太强,水分蒸发量大, 光照和土壤水分条件不适合幼龄野核桃生长发育,导致丛生野核桃株数少;阴坡光照又相对较弱,也不适合幼龄野核桃生长发育,致使丛生野核桃株数占比小;半阳坡和半阴坡的光照条件、土壤水分条件可能正好符合丛生野核桃生长发育需要,则丛生野核桃株数占优势;阳坡和阴坡丛生野核桃株数远少于半阳坡和半阴坡,推测同样是光照强度和土壤水分条件(阳坡光照过强,土壤水分不足;阴坡光照不足,土壤含水量太大[15])造成的。因此,推测丛生野核桃生长发育的限制因子主要是光照、土壤水分,这需要通过生理控制实验来验证。丛生野核桃这种分布差异可能蕴含野核桃种群在不同坡向上的生长与物质分配策略,因为植物在生长发育过程中,总是不断调整其生长和分配策略,但也要遵循生长与物质分配相协调的原则[29–30]。有研究表明丛生乔木的分布与土壤剖面砂粘比呈正相关关系,即砂粘比越高,丛生株越多;反之,丛生株越少[31], 由此推断保护区半阳坡、半阴坡比阳坡、阴坡土壤剖面砂粘比更高,使得丛生野核桃在半阳坡、半阴坡分布株数明显多于阳坡和阴坡,这还需深入研究。
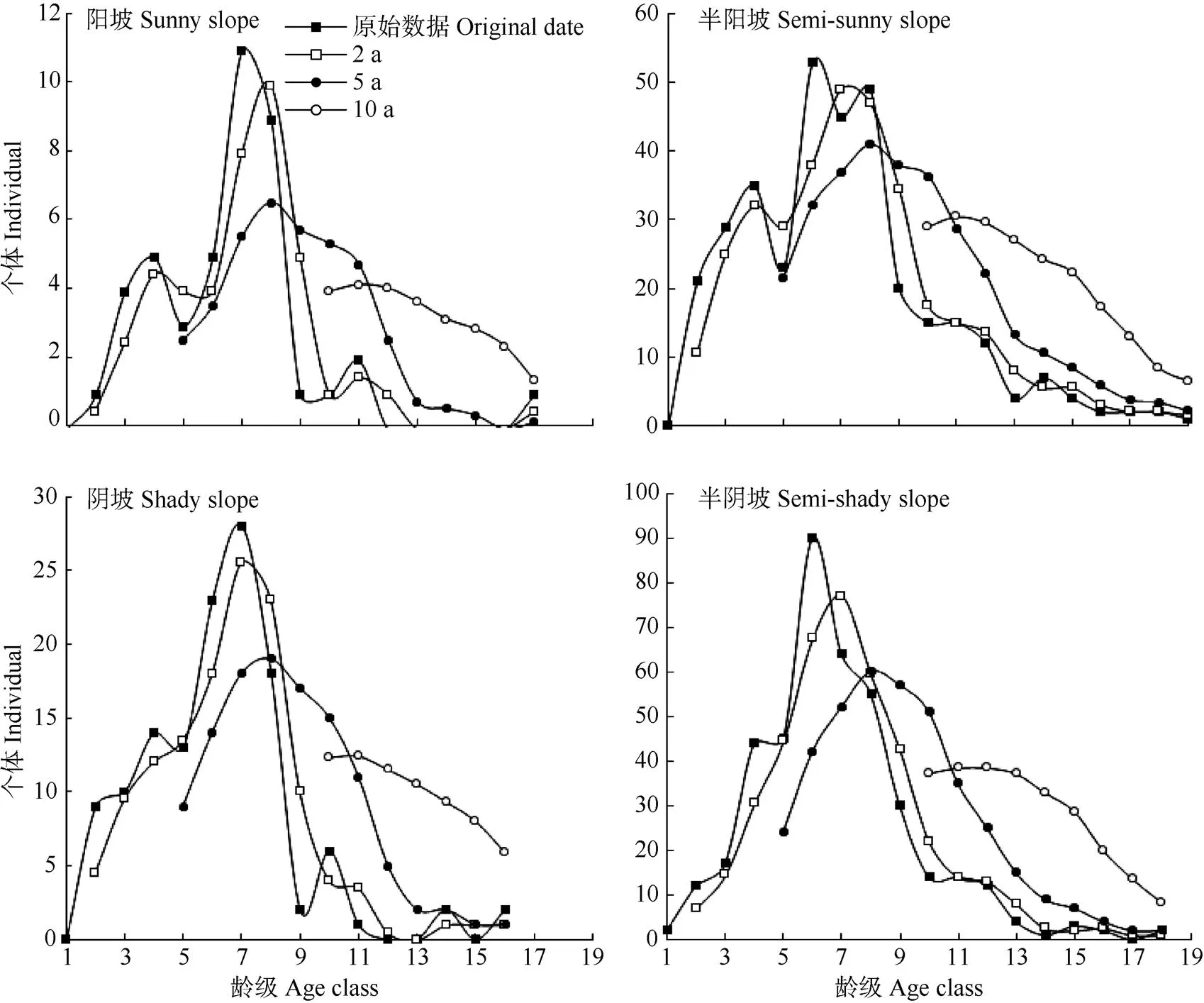
图5 不同坡向丛生野核桃种群数量动态的时间序列模型
4.3 丛生野核桃种群的年龄结构与动态趋势
种群年龄结构是生物学特征与生存环境共同作用的结果,其变化影响生态系统的稳定和维持[28]。按照径级,野核桃种群有19个龄级[15],丛生野核桃种群也有19个龄级,表明野核桃丛生现象是在环境长期干扰下,野核桃种群形成的最有效的种群维持机制之一。丛生野核桃年龄结构呈纺锤型,不同坡向丛生野核桃种群目前处于衰退状态,这与整个野核桃种群的年龄结构研究结果不一致[15],可能是种群定义边界、研究区域生境划分标准不同造成的,也可能是丛生野核桃这一“小”种群的固有特性决定的。因此未来可以对野核桃自然保护区所有野核桃5 500余株[15](不采用样方法)进行全面调查, 统计年龄组成进行深入分析。
种群数量动态可以了解种群的形成机制及其生长机制[1,3]。数量化动态分析表明,不同坡向Vʹ和Vʹ虽为正数,但Vʹ值接近0,丛生野核桃种群稳定性差、种群抗干扰能力弱,且目前种群幼龄级个体严重匮乏,中老龄个体数量丰富,如若不能及时采用有效措施促进丛生幼苗补充,丛生野核桃种群将难以维持而呈老化趋势。4个坡向丛生野核桃种群死亡最高峰均集中在中、老龄级,当野核桃生长发育到中龄级,其树高、冠幅都得到极大扩展,对水分、养分等资源需求量激增,种内竞争加剧,导致较高的死亡率;老龄级野核桃的高死亡率,是生物学特性决定的,野核桃生长发育到老龄级,生理机能衰退、抗逆性差,死亡率自然升高。此外,引起中老龄级丛生野核桃死亡率明显增加也可能是负密度制约造成的[32]。从整个野核桃生活史上来看,中、老龄级丛生野核桃个体死亡后,其树干可发生根系[9],不仅为该野核桃组其他个体生长发育创造了有利的生存条件,还为丛生野核桃种群更新和发展提供了最优的生存策略。
4.4 丛生野核桃种群保护措施
一方面,野核桃自然保护区是一个保护野核桃及其生存环境的自然保护区,开垦、樵采、放牧等活动受到严格的限制,缺乏适度干扰活动改变丛生野核桃年龄结构;另一方面,尽管目前在丛生野核桃生长过程中不断萌生新的个体,但由于负密度效应、空间生态位限制等因素均可导致可持续更新的幼苗数量不足,如若继续保持目前丛生野核桃的自然生长状态,极可能造成林分的衰败。因此需要采用适当的干扰措施来调整丛生野核桃的年龄结构与动态。
1) 阳坡:对丛生野核桃幼苗采用遮光处理,避免光照太强,水分蒸发量太大的问题;阴坡:对林下灌木采取择伐、间伐,减小郁闭度,以解决光照不足的问题。
2) 根据低温、寒潮时野核桃幼苗易形成分叉主干的特性[9],在野核桃坚果成熟季采摘野核桃时有意识预留一些野核桃,使实生苗种源充足,提高野核桃丛生机会。
3) 根据野核桃自然保护区处于“逆温带”的特点,再结合保护措施2),在保护区野核桃分布的海拔高限和低限位置,尝试在保护区人工埋种野核桃坚果,增加实生苗的数量,以此提高丛生株的数量。
[1] KANF D, GUO Y X, REN C J, et al. Population structure and spatial pattern of main tree species in secondaryforest in Ziwuling Mountains, China [J]. Sci Rep, 2014, 4(1): 6873. doi: 10. 1038/srep06873.
[2] YAO L, AI X R, YI Y M, et al. Structure and dynamics of dominant populations in the mixed forest of subtropical evergreen and deciduous broad-leaved tree species in the Southwest of Hubei Province [J]. Sci Silv Sin, 2017, 53(2): 10–18. [姚兰, 艾训儒, 易咏梅, 等. 鄂西南亚热带常绿落叶阔叶混交林优势种群的结构与动态特征 [J]. 林业科学, 2017, 53(2): 10–18. doi: 10.11707/j.1001-7488.20170202.]
[3] ZHAO Y, CAO X W, LI B, et al. Structural characteristics of 4 natural populations in the southern forest region of Gansu Province [J]. Sci Silvae Sin, 2020, 56(9): 21–29. [赵阳, 曹秀文, 李波, 等. 甘肃南部林区4种天然林种群结构特征 [J]. 林业科学, 2020, 56(9): 21–29. doi: 10.11707/j.1001-7488.20200903.]
[4] CHU C J, ADLER P B. When should plant population models include age structure? [J]. J Ecol, 2014, 102(2): 531–543. doi: 10.1111/1365- 2745.12212.
[5] CHEN X D. A study on the method of quantitative analysis for plant population and community structural dynamics [J]. Acta Ecol Sin, 1998, 18(2): 214–217. [陈晓德. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, 1998, 18(2): 214–217. doi: 10.3321/j.issn:1000- 0933.1998.02.017.]
[6] HAN L, WANG J Q, WANG H Z, et al. The population structure and dynamics ofat the upper reaches of the Tarim River [J]. Acta Ecol Sin, 2014, 34(16): 4640–4651. [韩路, 王家强, 王海珍, 等. 塔里木河上游胡杨种群结构与动态[J]. 生态学报, 2014, 34(16): 4640–4651. doi: 10.5846/stxb201212271887.]
[7] HARCOMBE P A. Tree life tables [J]. Bioscience, 1987, 37(8): 557– 568. doi: 10.2307/1310666.
[8] HE P. Conservation Biology of the Rare & Endangered Plants [M]. Chongqing: Southwest China Normal University Press, 2005: 205–206.[何平. 珍稀濒危植物保护生物学[M]. 重庆: 西南师范大学出版社, 2005: 205–206.]
[9] ZENG B. Present status, problems, and further development strategies on natureDode resource and their exploitation in Xinjiang [J]. N Fruits, 2005(4): 1–3. [曾斌. 新疆野生核桃资源的现状与发展[J]. 北方果树, 2005(4): 1–3. doi: 10.3969/j.issn.1001-5698. 2005.04.001.]
[10] ZHANG W. Basic researches on conservation biology of endangered speciesin Tianshan Valley in Xinjiang, China [D]. Changchun: Northeast Normal University, 2016. [张维. 新疆天山峡谷渐危植物野核桃保护生物学基础研究 [D]. 长春: 东北师范大学, 2016.]
[11] ZHANG W, YANG Y F, LI J D. Dynamic of the population quantity ofin different habitats in Xinjiang, China [J]. Pak J Bot, 2015, 47(3): 911–918.
[12] ZENG B, HE T M, WU Y X, et al. Tissue culture and plantlet regeneration ofDode in Xinjiang [J]. Xinjiang Agric Sci, 2011, 48(7): 1227–1230. [曾斌, 何天明, 吴玉霞, 等. 新疆野生核桃的组织培养和植株再生[J]. 新疆农业科学, 2011, 48(7): 1227–1230.]
[13] ZHANG W, YANG Y F, LI J D. Static life table and growth analysis of seedlings ofin Xinjiang, China [J]. Aust J For Sci, 2015, 132(3): 131–144.
[14] ZHANG W, JIA F Q, NA S B T, et al. Population dynamics and growth ofseedlings of different slope aspects in the west Tianshan Valley in Xinjiang, China [J]. Chin J Ecol, 2014, 33(10): 2596–2602. [张维, 贾风勤, 纳森巴特, 等. 新疆西天山峡谷不同坡向野核桃幼苗种群动态及生长分析[J]. 生态学杂志, 2014, 33(10): 2596–2602. doi: 10.13292/j.1000-4890.2014.0219.]
[15] ZHANG W, JIAO Z W, REN Y L, et al. Life tables and fluctuation cycle ofpopulations on different slope aspects in wild walnut natural conservation area of Xinjiang, China [J]. Acta Bot Boreali-Occid Sin, 2015, 35(6): 1229–1237. [张维, 焦子伟, 任艳利, 等. 新疆野核桃自然保护区不同坡向野核桃种群生命表与波动周期[J]. 西北植物学报, 2015, 35(6): 1229–1237. doi: 10.7606/j.issn. 1000-4025.2015.06.1229.]
[16] LIU X L, CHEN X S, ZHANG M Y, et al. Population genetic structure analysis ofusing SSR markers [J]. J Fruit Sci, 2008, 25(4): 526–530. [刘晓丽, 陈学森, 张美勇, 等. 普通核桃() 3个群体遗传结构的SSR分析 [J]. 果树学报, 2008, 25(4): 526– 530. doi: 10.3969/j.issn.1009-9980.2008.04.016.]
[17] WANG Z Y. Analysis of resource and genetic diversity of wild walnut in Xinjiang [D]. Urumqi: Xinjiang Agricultural University, 2011. [王肇延. 新疆野核桃资源及遗传多样性的分析 [D]. 乌鲁木齐: 新疆农业大学, 2011.]
[18] SHE D Y. Formation and characteristics of wild walnut forest soils in Gongliu County [J]. Arid Zone Res, 1994, 11(2): 11–15. [佘定域. 巩留野核桃林土壤的形成及其特征特性[J]. 干旱区研究, 1994, 11(2): 11–15.]
[19] LIU L C, PAI Z L, XU H J. The formation characteristics and taxonomic classification of the soil under the wild-fruit forests in Yili Valley, Xinjiang [J]. Arid Land Geo, 1997, 20(2): 34–40. [刘立诚, 排祖拉, 徐华君. 伊犁谷地野果林下的土壤形成特点及其系统分类 [J]. 干旱区地理, 1997, 20(2): 34–40. doi: 10.3321/j.issn:1000-6060. 1997.02.005.]
[20] CHEN C. Study on slope, aspect and slope position extraction algorithm for forest subcompartments [D]. Hangzhou: Zhejiang Agricultural & Forestry University, 2019. [陈晨. 面向林业小班的坡度、坡向及坡位提取算法研究[D]. 杭州: 浙江农林大学, 2019.]
[21] ZHANG J T, RU W M. Population characteristics of endangered speciesvar.and its conservation strategy in Shanxi, China [J]. Popul Ecol, 2010, 52(3): 407–416. doi: 10.1007/s 10144-009-0192-y.
[22] ZHANG W, LI H Y, LAI X H, et al. Distribution patterns ofpopulations at different slope aspects in Tianshan Valley in Xinjiang, China [J]. Chin J Appl Ecol, 2016, 27(10): 3105–3113. [张维,李海燕, 赖晓辉, 等. 新疆天山峡谷不同坡向野核桃种群分布格局 [J]. 应用生态学报, 2016, 27(10): 3105–3113. doi: 10.13287/j.1001- 9332.201610.004.]
[23] WRATTEN S D, FRY G I A. Field and Laboratory Exercises in Ecology [M]. London: Edward Arnold Publishers Limited, 1980: 27–35.
[24] JIANG H. Ecological Study ofPopulations [M]. Beijing: China Forestry Publishing House, 1992: 11–12. [江洪. 云杉种群生态学 [M]. 北京: 中国林业出版社, 1992: 11–12.]
[25] HETT J M, LOUCKS O L. Age structure models of balsam fir and eastern hemlock[J]. J Ecol, 1976, 64(3): 1029–1044. doi: 10.2307/ 2258822.
[26] ZU Y G. Conservation Biology of the Endangered Plants:[M]. Beijing: Science Press, 1999: 215–223. [祖元刚. 濒危植物裂叶沙参保护生物学 [M]. 北京: 科学出版社, 1999: 215–223.]
[27] BARNES M E. Seed predation, germination and seedling establish- ment ofin northern Botswana [J]. J Arid Environ, 2001, 49(3): 541–554. doi: 10.1006/jare.2001.0805
[28] ZHANG Z, YAN Z F. The investigation ofin Xinjiang [J]. Xinjiang Agric Sci, 1962(10): 404–407. [张钊, 严兆福. 新疆野生核桃的调查研究 [J]. 新疆农业科学, 1962(10): 404–407.]
[29] WHITE J. The allometric interpretation of the self-thinning rule [J]. J Theor Biol, 1981, 89(3): 475–500. doi: 10.1016/0022-5193(81)90363-5.
[30] ZHANG W, LUO X Z, ZHANG N, et al. Phenotypic variability and growth characteristics of wild walnut seeds in Xinjiang, Northwest China [J]. Chin J Appl Ecol, 2013, 32(9): 2281–2288. [张维, 罗新泽, 张娜, 等. 新疆野核桃种子的表型变异及生长特征[J]. 生态学杂志, 2013, 32(9): 2281–2288. doi: 10.13292/j.1000-4890.2013.0351.]
[31] TAN Y P, LIU C, DING Y Y, et al. Investigation and evaluation of the growth of caespitose[J]. J Chin Urban For, 2015, 13(3): 60–62. [谭艳萍, 刘畅, 丁蕴一, 等. 美国丹佛生态廊道丛生美洲黑杨生长量调查与评价 [J]. 中国城市林业, 2015, 13(3): 60–62. doi: 10.3969/j. issn.1672-4925.2015.03.016.]
[32] CHEN G P, ZHAO W Z. Age structure and dynamics ofbranches in southern edge of the Mu US sandy land [J]. J Desert Res, 2015, 35(6): 1520–1526. [陈国鹏, 赵文智. 毛乌素沙地南缘沙柳()丛生枝年龄结构与动态特征 [J]. 中国沙漠, 2015, 35(6): 1520–1526. doi: 10.7522/j.issn.1000-694X.2015.00001.]
Age Structure and Quantitative Dynamics of Cespitosein Different Slope Directions in Xinjiang Wild Walnut Nature Reserve
WEI Haiying1, LI Yiyang2, LAI Xiaohui1, ZHANG Wei1*
(1. School of Biology and Geography, Yili Normal University; Institute of Resources and Ecology, Yili Normal University,Yining 835000, Xinjiang, China; 2. Yili Vocational and Technical Collage of the 4th Division,Yining 835000, Xinjiang, China)
Based on the general survey ofin Xinjiang Wild Walnut Nature Reserve, the diameter level structure was used instead of age structure, the static life table of population was compiled by smoothing technique, and the population dynamics was predicted by time series model, so as to obtain the age level structure and dynamics of. The results showed that the middle age class was dominant in different slope directions, and the age structure was spindle-shaped with two small ends and big middle. Although the dynamic indexVof age structure of the four slope populations was greater than 0, the maximum was only 19.10%, andVʹ was extremely low, the maximum is 1.12%, the growth trend ofpopulation on sunny slope was the highest and the anti-interference ability and the population stability was the strongest, whereas the semi-sunny slope showed the lowest growth trend, the worst anti-interference ability and stability. There were multiple death peaks in all slope populations. The highest peak was at 15 age class on sunny slope, 15 and 18 age class on semi-sunny slope, and 9 age class on shady slope and semi-shady slope. The survival curves were closer to Deevey-II type. The number of individuals at middle age would decline after 2, 5 and 10 age classes in the future, while the number of old individuals would increase. Therefore, it was very critical for the protection of cespitoseof young individuals.
; Static life table; Age structure; Population dynamic
10.11926/jtsb.4573
2021-11-21
2022-02-20
国家自然科学基金项目(31560095, 31160072)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31560095; 31160072).
魏海英(1995年生),女,硕士研究生,主要从事植物生态学研究。E-mail: 1354447420@qq.com
通讯作者Corresponding author.E-mail: 343956580@qq.com
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
南方农业(2022年23期)2022-02-14 09:45:26
思维与智慧·下半月(2021年7期)2021-08-13 16:55:24
河南农业大学学报(2020年2期)2020-05-22 02:56:30
江河文学(2020年6期)2020-01-04 07:06:37
防护林科技(2019年7期)2019-08-19 00:50:58
山西林业科技(2019年1期)2019-05-27 12:24:52
现代农村科技(2018年11期)2018-11-15 11:27:58
浙江农业科学(2016年11期)2016-05-04 04:16:48
华声(2015年11期)2015-06-10 11:35:58
